
Abstract
Background. Impaired voluntary neuromuscular activation of agonist muscles is a primary determinant of weakness and motor dysfunction following stroke. Objective. To determine whether eccentric resistance training (ECC) resistance training is superior to concentric resistance training (CON) resistance training to enhance neuromuscular activation, strength, and walking speed after stroke. Methods. A total of 34 adults poststroke participated in a staged intervention comprising (1) either CON-only or ECC-only resistance training of the paretic leg followed by (2) gait training. Changes in voluntary neuromuscular activation and power were assessed for both the trained paretic and untrained nonparetic legs. Self-selected and fast walking speeds were also assessed. Results. In response to resistance training, the ECC group experienced larger improvements in neuromuscular activation of paretic leg muscles, rectus femoris and vastus medialis (P < .005), and the largest gains in paretic leg power (+74% for ECC contractions, P < .0001). ECC also had greater cross-education of increased power to the untrained nonparetic leg (12%-14%, P = .006). Over the course of gait training, much of the gain in paretic leg activation in the ECC group was lost, such that the net change in agonist activation was comparable between the CON and ECC groups when the full intervention was completed. Nevertheless, improvement in walking speed postintervention was more prevalent in the ECC than CON group. Conclusion. ECC resistance training was more effective for improving bilateral neuromuscular activation, strength, and walking speed following stroke. Future research should assess whether a longer duration ECC training program can provide further benefit.
Introduction
Impaired ability to voluntarily activate the agonist musculature is a primary determinant of weakness and coincident motor dysfunction following stroke.1-3 Accordingly, persons poststroke are likely to benefit from rehabilitation interventions that maximize the capacity for voluntary neuromuscular activation. In adults poststroke and in nondisabled young and older adults, resistance training is effective for increasing strength and neuromuscular activation.4-10 Resistance training has also been shown to produce substantial improvements in functional performance after stroke, including walking, stair climbing, chair-stand, balance, and upper-extremity function, including reaching.8,9,11-16,27 With accumulating evidence of the beneficial effects of resistance training as a component of motor rehabilitation, it is now critical to identify the training parameters that provide the most potent stimulus for driving motor recovery.
The use of eccentric contractions in resistance training may provide unique benefits for increasing neuromuscular activation poststroke. Eccentric strength is more preserved than concentric strength following stroke,1,16 suggesting that training with eccentric contractions may provide a more intense training stimulus. Furthermore, evidence from healthy adults suggests that supraspinal motor systems are engaged differently by eccentric contractions when compared with concentric contractions. 17 For instance, cortical activation measured with EEG during eccentric contractions shows earlier onset, higher magnitude, and larger area of activation.18,19 Training with eccentric contractions also induces a larger cross-transfer effect to the untrained limb,20,21 indicating greater involvement of bilateral motor control mechanisms. In adults poststroke, Engardt and colleagues 7 noted that the strength of the trained paretic leg, relative to the untrained nonparetic leg, improved in an eccentrically trained but not a concentrically trained group. Also, the symmetry of body-weight loading during sit to stand was improved in the eccentric group but not the concentric group. 7 In support of these findings, a review by Patten et al 3 notes that interventions that have focused on the use of eccentric contractions seem to indicate larger strength gains that generalize to enhanced function in persons poststroke.
Here, we conducted a randomized controlled trial* of dynamic resistance training followed by clinic-based gait training in adults in the intermediate phase of recovery from stroke. The overall goal of the intervention was to enhance walking function by increasing the capability for lower-extremity neuromuscular activation and power production. The specific objective of the study was to determine whether resistance training with eccentric contractions is superior to training with concentric contractions for accomplishing this goal. We hypothesized that eccentric training would be more effective for increasing quadriceps neuromuscular activation and dynamic strength (eg, power) in both the trained paretic legs and untrained nonparetic legs. We further hypothesized that eccentric training would be more effective in improving walking speed.
Methods
Participants
In all, 34 adults meeting the following inclusion criteria participated: clinical presentation of a single, unilateral stroke (confirmed by CT or MRI) 6 to 18 months prior to enrollment, ability to ambulate independently a minimum of 25 feet on a level surface using a walking aid and/or ankle-foot orthosis at a minimum of 0.30 m/s, 22 absence of lower-extremity joint pain or passive range of motion limitations, and no major deficits of sensation or proprioception.
Study Design
The study involved a staged intervention of 5 weeks of dynamic high-intensity resistance training followed by 3 weeks of clinic-based gait training. During each phase of the intervention, treatment was delivered 3 times weekly. In the first phase, participants were randomized to either a concentric resistance training group (CON) or an eccentric resistance training group (ECC; Figure 1). To ensure comparable baseline motor function between groups, the synergy subscore of the Fugl-Meyer Lower Extremity Motor Assessment 24 was used to stratify participants as higher (15-22 points) or lower functioning (0-14 points). A blinded assessor performed assessments at baseline, following resistance training, and following gait training, including overground walking speed, dynamometric assessment of isokinetic knee extension strength, and neuromuscular activation.
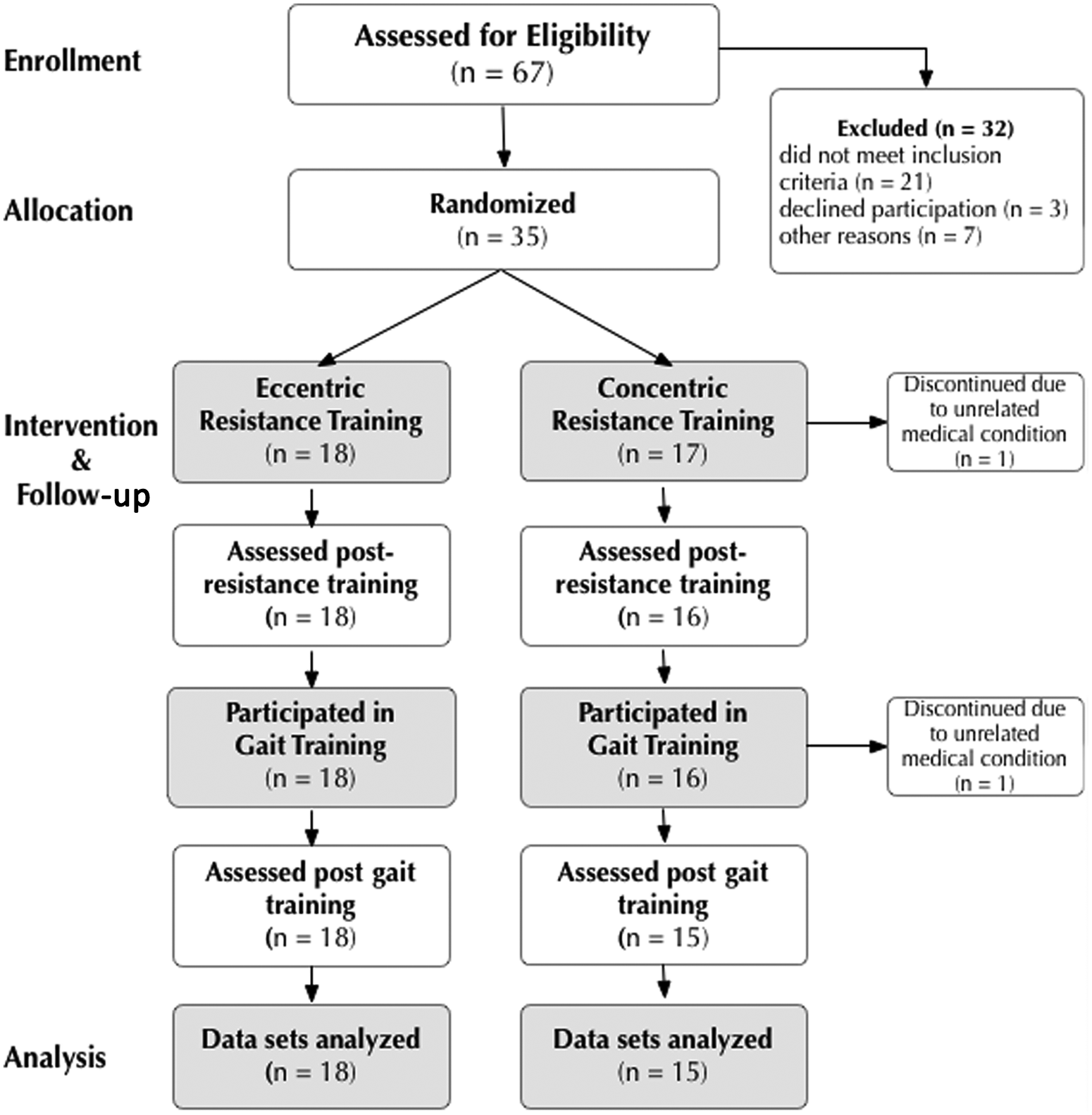
Flow diagram of participant progression through the trial.
Dynamic Resistance Training
Paretic limb resistance training was performed 3 days per week using a Biodex System 3 Pro isokinetic dynamometer (Biodex Medical Systems Inc, Shirley, New York). The nonparetic leg was not trained. Each 90-minute session involved one-on-one training with a licensed physical therapist. Training was progressed according to the schedule presented in Table 1. We used a triangle pyramid training paradigm, which repeated lower criterion speeds in weeks 4 and 5. Our rationale was to first train to increase the speed at which force could be produced and then increase the load produced during dynamic contractions. Joint actions trained included the following: ankle dorsiflexion and plantarflexion, knee extension and flexion, hip abduction and a multisegmental task involving hip flexion/extension, knee extension/flexion, and ankle plantarflexion/dorsiflexion. Attachments were custom designed to enable optimal biomechanical alignment for proper performance of each movement. Verbal encouragement was provided throughout the training sessions to motivate maximal effort. This form of training is conceptually similar to power training, which involves rapid muscle contractions. 24 Power training is considered a safe but intense form of training that is more effective than traditional resistance training that uses slower contractions.7,9,15,25-27
Prescription for Dynamic Resistance Training a
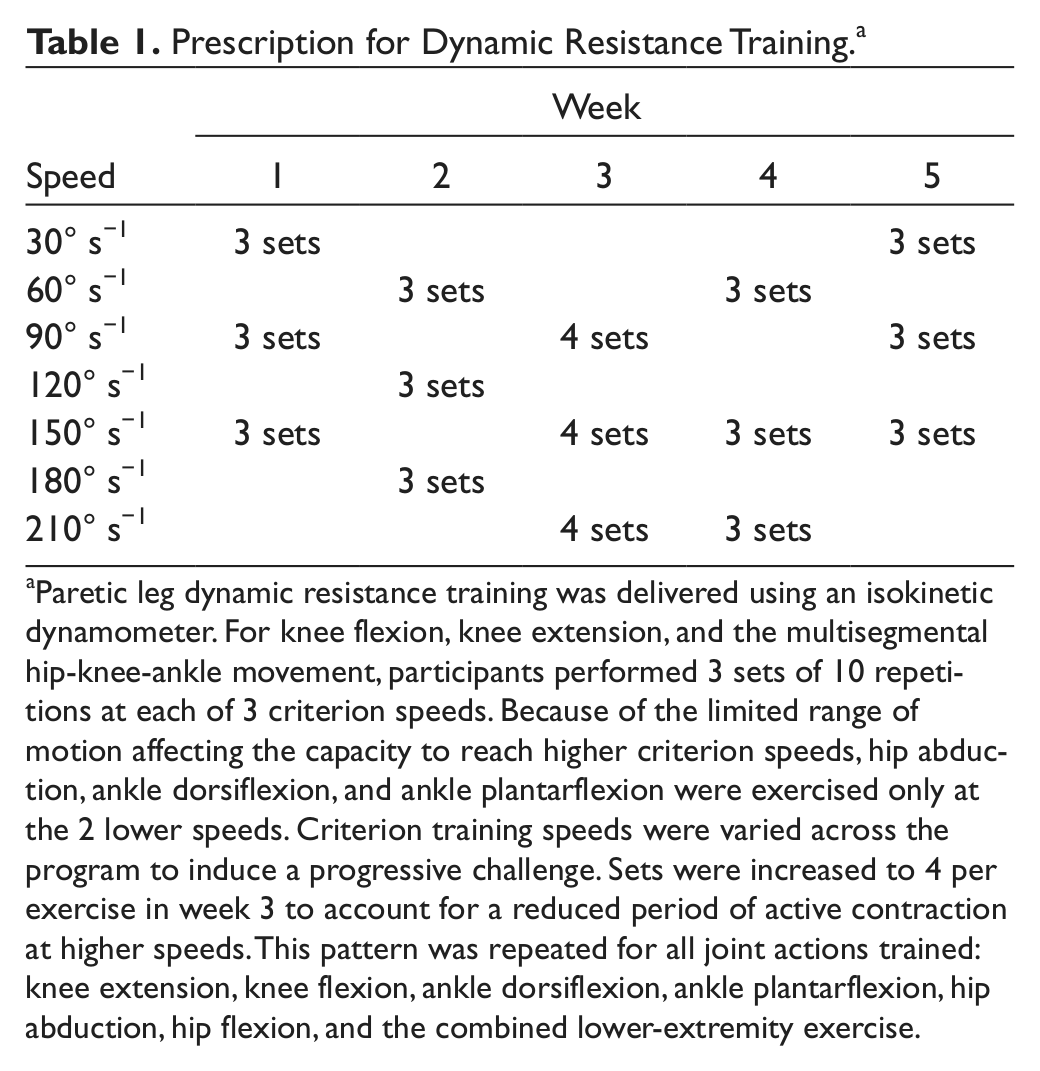
Paretic leg dynamic resistance training was delivered using an isokinetic dynamometer. For knee flexion, knee extension, and the multisegmental hip-knee-ankle movement, participants performed 3 sets of 10 repetitions at each of 3 criterion speeds. Because of the limited range of motion affecting the capacity to reach higher criterion speeds, hip abduction, ankle dorsiflexion, and ankle plantarflexion were exercised only at the 2 lower speeds. Criterion training speeds were varied across the program to induce a progressive challenge. Sets were increased to 4 per exercise in week 3 to account for a reduced period of active contraction at higher speeds. This pattern was repeated for all joint actions trained: knee extension, knee flexion, ankle dorsiflexion, ankle plantarflexion, hip abduction, hip flexion, and the combined lower-extremity exercise.
Gait Training
Gait training sessions were held 3 days per week. Each 90-minute session involved one-on-one training with a licensed physical therapist and included: stretching (15 minutes), activities to target specific components of gait (30 minutes), balance and/or obstacle course (15 minutes), and treadmill walking (30 minutes). Treatment goals for components of gait were based on principles of motor relearning 28 and advanced from weight acceptance (week 1) to single-limb support (week 2) and limb advancement (week 3). The treadmill walking program was motivated by the approach of Pohl and coworkers 29 and involved interval training with short bouts (75-150 s) of walking and striding (ie, taking longer than normal steps) at the participant’s maximal speed, which was determined during the first session of each week. Partial body-weight support was reduced progressively from 30% (week 1) to 10% (week 3). No therapist assistance was provided for either limb advancement or kinematics.
Assessment
We evaluated quadriceps neuromuscular performance during leg extension. This action produces reliable and stable assessment of lower-extremity performance in individuals poststroke.30-32 Assessments were conducted at baseline, following the resistance training intervention, and following the gait training intervention. Neuromuscular activation and strength of the knee extensors were assessed concurrently using an isokinetic dynamometer and surface electromyography (Motion Lab Systems MA300, Baton Rouge, Louisiana). Torque, angular position, and angular velocity were acquired directly from the dynamometer. EMGs were recorded from the rectus femoris (RF), vastus medialis (VM), semitendinosus (ST), and biceps femoris (BF) using double differential surface electrodes with 17-mm interelectrode distance (Motion Lab Systems MA411, Baton Rouge, Louisiana). All signals were acquired at a sampling rate of 1 kHz using a Powerlab/16SP A/D system (ADInstruments, Colorado Springs, Colorado) and LabChart software and saved directly to disk for offline analysis. A detailed description of our assessment paradigm has been described previously. 1 Briefly, participants were secured to the chair of the dynamometer with the knee aligned to the dynamometer axis of rotation. Participants were instructed to keep their arms folded across the chest or on the lap. Both the paretic and nonparetic legs were tested over a range of velocities, including concentric (30° to 180° s−1) and eccentric (−30° to −180° s−1). Participants were instructed to “push as hard and as fast as you can” or “resist and try to stop the dynamometer” for concentric and eccentric trials, respectively. The range of motion for dynamic knee extension was set to 0° to 90° of knee flexion. Data from passive limb movement trials were used to correct for the effects of limb weight and passive resistance to movement.
Walking speed was measured during self-selected (SS) and fastest comfortable (FC) overground walking using a motion analysis system (Qualysis, Gothenburg, Sweden). Speed was calculated by tracking the anterior displacement of the calcaneus marker over at least 6 complete gait cycles and dividing by the concurrent change in time.
Data Analysis
Analysis of torque and muscle activation data were performed using Matlab 7.0 (The Mathworks, Natick, Massachusetts) and JMP statistical software (v 7 SAS Institute Inc, Cary, North Carolina). Torque recorded during maximal voluntary efforts was corrected for the effects of limb weight and resistance to passive movement. Torque was calculated as the mean value between 50° and 60° of knee flexion. Raw EMG signals were corrected for baseline offset and band-pass filtered using a bidirectional second-order Butterworth filter (10-500 Hz), and the root-mean-square amplitude was calculated over the same window just described for torque. These criteria capture torque and neuromuscular activation at biomechanically equivalent conditions across contraction modes and participants. Torque was converted to power by multiplying the torque value by the concurrent angular velocity in radians/s. For each participant, power and activation were averaged across trials for each criterion velocity. To assess potential changes in excessive antagonist activity caused by resistance training, a coactivation ratio was calculated for each CON criterion velocity for each participant by expressing antagonist muscle activation magnitude relative to agonist muscle activation magnitude: (ST + BF)/(RF + VM).
Statistics
Power and neuromuscular activation were analyzed using mixed-model repeated-measures analysis of variance (ANOVA). Separate tests were used for each variable (power, RF activation, and VM activation). Of primary interest for within-groups analysis was the effect of time (ie, pretraining, postresistance training, and postgait training) and the effect of Time × Contraction Mode. Of primary interest for between-groups analysis was the Group × Time × Contraction Mode interaction effect or, in the absence of a difference, Group × Time interaction. The main effect of group was not interpreted because it is not meaningful without the added context provided by Time and Contraction Mode.
Self-selected and fast walking speeds were analyzed using paired t tests for within-groups analysis and mixed-model repeated-measures ANOVAs for between-groups analysis. Of primary interest for between-groups analysis was the Group × Time interaction effect. For all variables, post hoc analysis of significant ANOVA effects was conducted with paired t tests. In addition, walking speed was evaluated according to the proportion of responders and nonresponders in each group using Fisher exact test. Responders were defined as participants whose change in walking speed exceeded a minimal important difference, defined as one half of the standard deviation of baseline walking speed for all study participants. 33 This criterion was applied separately to self-selected and fast walking speeds. We considered a participant to be a responder if either self-selected or fast speed (not necessarily both) met the criterion.
Results
Participants
The number of participants who were randomized to and completed CON and ECC resistance training were 15 and 18, respectively. Baseline demographics are shown in Table 2. The groups did not differ for any baseline variable measured (P > .05).
Participant Demographics at Baseline
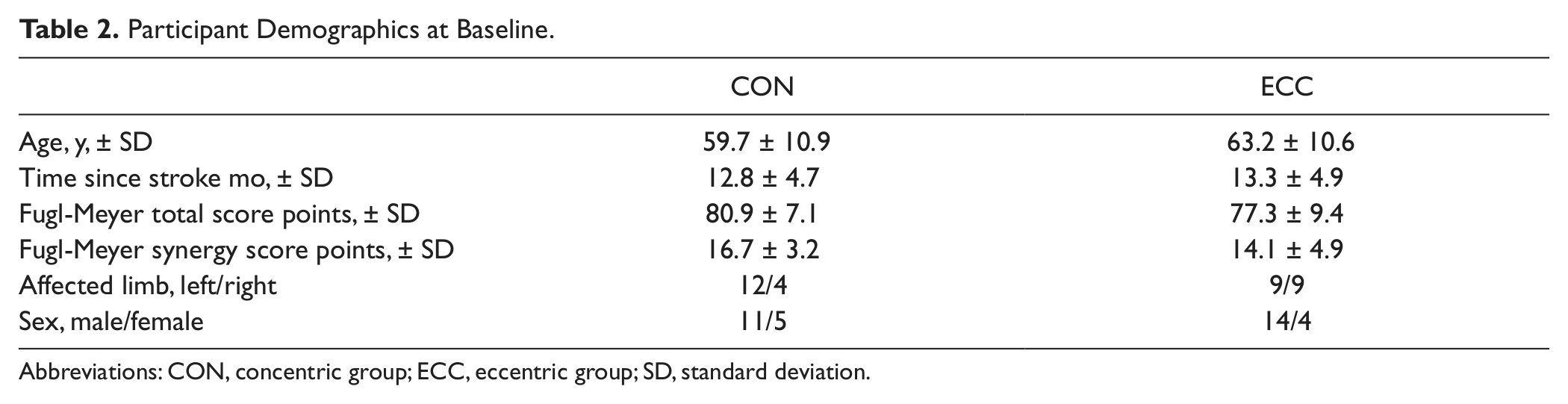
Abbreviations: CON, concentric group; ECC, eccentric group; SD, standard deviation.
Paretic (trained) leg power
Figure 2A summarizes the changes in paretic leg power as percentage change following each training phase. Within both CON and ECC groups, ANOVA for power revealed a significant effect of Time (P < .0001) and Time × Contraction Mode (P < .0001 for ECC, P = .03 for CON). Post hoc analysis revealed specificity of training during the resistance training phase. Power gains for ECC were greater for eccentric (+74%) than concentric (+25%) contractions (P < .0001). Power gains for CON were greater for concentric (+51%) than eccentric (+29%) contractions (P = .02). Post hoc analysis also revealed that power production was not significantly altered during the gait training phase in either group. Accordingly, at the end of the intervention, both concentric and eccentric groups had retained a significant power gain over baseline levels for both concentric and eccentric contraction modes (all P < .005).
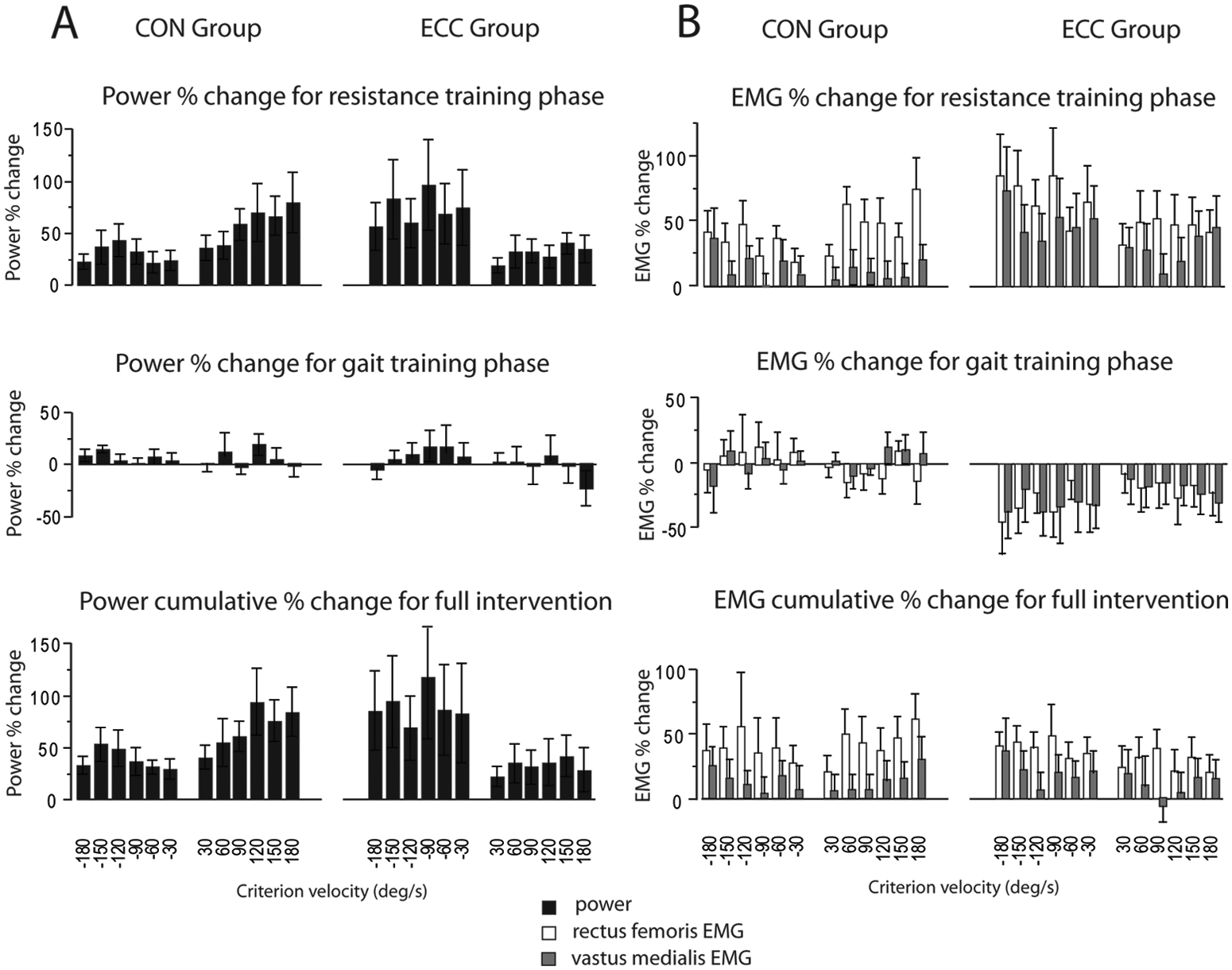
Power and neuromuscular activation of the paretic leg: The percentage change for paretic knee extensor power (A) and neuromuscular activation (B) are illustrated for the concentric group (CON) and eccentric group (ECC) at each testing velocity. Percentage change is shown in response to the resistance training phase, gait training phase, and cumulatively over the full intervention.
Between-groups ANOVA for power revealed a significant Group × Time × Contraction Mode interaction (P = .02). Post hoc analysis revealed that this result was primarily a result of the task specificity of resistance training, with ECC and CON demonstrating the largest gains in eccentric and concentric power, respectively, compared with other Group/Time/Contraction Mode combinations (P < .05).
Paretic (trained) leg activation
Figure 2B summarizes the changes in paretic leg activation as percentage change following each training phase. Within both CON and ECC groups, ANOVA for RF activation revealed a significant effect of Time for CON and ECC groups (P < .0001) and a significant effect of Time × Contraction Mode for the ECC group only (P = .009). Post hoc analysis revealed that resistance training increased RF activation in both groups (P < .0001). Specifically, RF activation increased for both concentric and eccentric contractions in the CON group (+41% and +28%, respectively) and ECC group (+44% and +69%, respectively). Post hoc analysis revealed that the ECC group exhibited task specificity of resistance training, with larger gains in RF activation during eccentric versus concentric contractions. The ECC group also experienced somewhat reduced RF activation following gait training (−17% and −30% for concentric and eccentric contractions, respectively; P = .003).
Between groups, the Time × Group × Contraction Mode interaction for RF activation was not significant (P = .11). However, there was a significant Time × Group interaction (P = .0012). Post hoc analysis revealed that this was because relative to the CON group, the ECC group revealed larger increases in RF activation following resistance training (P = .002) and reduced RF activation after gait training (P = .002). By the end of the intervention, the net change in RF activation (averaged across all speeds) relative to baseline did not differ between groups (+40% in ECC and +33% in CON; P = .18).
Within groups, VM activation showed a significant effect of Time for the ECC group (P = .002) but not for the CON (P = .71) group. Time × Contraction Mode was not significant for either group. Post hoc analysis revealed that the ECC group had increased VM activation (averaged across all speeds) following resistance training (+32%, P < .001) and, subsequently, a reduction with gait training (−24%, P = .05). Similar to RF, the cumulative change postintervention (resistance + gait training) showed that neither group experienced a significant net change in VM activation relative to baseline (P > .46).
Between groups, the Time × Group × Contraction Mode Interaction for VM activation was not significant (P = .70). However, there was a significant Time × Group interaction (P = .005). Post hoc analysis revealed that this was because, relative to the CON group, the ECC group had both a larger increase in VM activation following resistance training (P = .003) and a greater reduction in VM activation post–gait training (P = .006). Antagonist coactivation was not increased in the CON or ECC groups following resistance training (P = .37 and .94, respectively).
Nonparetic (untrained) leg power
Figure 3A summarizes the changes in nonparetic leg power as percentage change following each training phase. Within groups, the ECC group but not the CON group had a significant effect of Time for power in the untrained nonparetic leg (P < .0001 and P = .47, respectively). There was also a marginally significant Time × Contraction Mode interaction effect in the ECC group (P = .065). Post hoc analysis revealed increased power in the ECC group (P < .0001) following resistance training, with the eccentric increase (+14%) being marginally larger than the concentric increase (+12%, P = .05). Post hoc analysis also revealed increased power (averaged across all velocities) in the ECC group following the gait training phase (+6%, P = .0003).
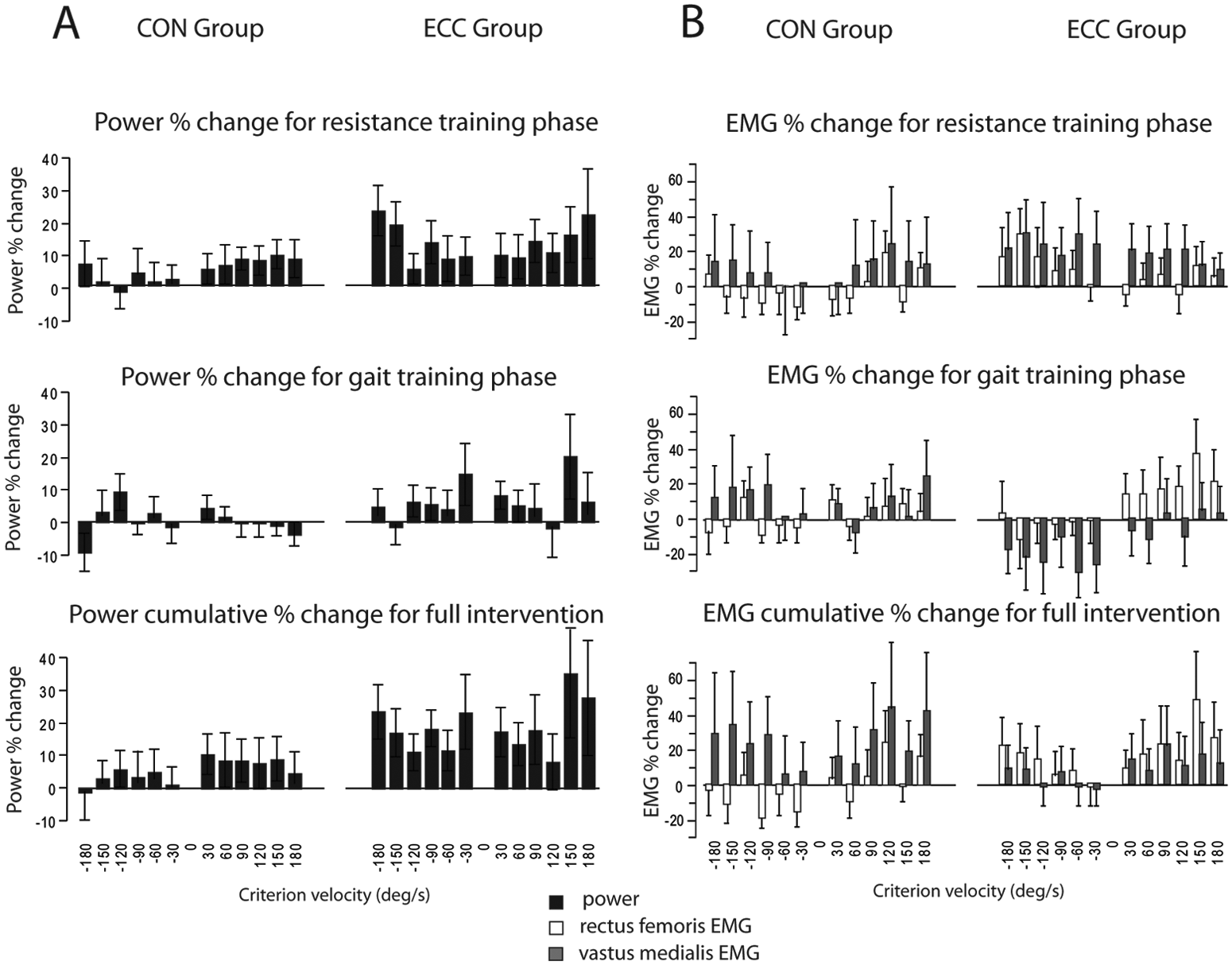
Power and neuromuscular activation of the untrained nonparetic leg: The percentage change for nonparetic knee extensor power (A) and neuromuscular activation (B) are illustrated for the concentric group (CON) and eccentric group (ECC) at each testing velocity. The percentage change is shown in response to the resistance training phase, gait training phase, and cumulatively over the full intervention.
Between groups, the Group × Time × Contraction Mode interaction was not significant (P = .14), but there was a strong Time × Group interaction (P < .0001). Post hoc analysis revealed that this was because ECC group power increased more than CON group power following both the resistance training (P = .006) and the gait training phases (P = .01).
Nonparetic (untrained) leg activation
Figure 3B summarizes the changes in nonparetic leg activation as percentage change following each training phase. The effect of Time for the CON group was significant for RF activation (P < .001), but post hoc analysis revealed that this effect was a result of reduced RF activation following resistance training (−4%). The effect of Time was not significant for ECC in either RF or VM (P > .11), although both showed a tendency for increased activation with resistance training (+11% and +24%, respectively, averaged across all speeds). The Time × Contraction Mode interaction effect was not significant for either group or muscle (all P > .11).
Between groups, the Group × Time × Contraction Mode interaction was not significant for either RF or VM (P = .54 and .79, respectively). However the Group × Time interaction was significant for RF (P = .0006) because of the previously mentioned reduction in activation in the CON group and a concomitant tendency toward increased activation in the ECC group following resistance training.
Walking Speed
Within groups, over the course of the full intervention (resistance training phase + gait training phase), the self-selected walking speed increased 0.12 m/s in both CON (P = .002) and ECC (P < .0001) groups. Fast walking speed increased 0.15 m/s in the CON group (P = .0006) and 0.18 m/s in the ECC group (P < .0001). Complete walking speed results by group and training phase are shown in Table 3. Between-groups analysis revealed that both groups improved similarly for both self-selected speed (P = .86) and fast walking speed (P = .73). However, the proportion of “responders,” that is individuals who increased either self-selected or fast walking speed by at least 1 minimal important difference (0.11 m/s for SS, 0.18 m/s for FC), was significantly greater in the ECC than in the CON group (81% vs 42%, P = .04). The median change in self-selected walking speed for the ECC group (0.112 m/s) was almost 2-fold that of the CON group (0.067 m/s). Even the lower quartile of the ECC group (0.074 m/s) was higher than the median of the CON group. Box plots illustrating the distribution of changes in walking speed are shown in Figure 4.
Walking Speeds
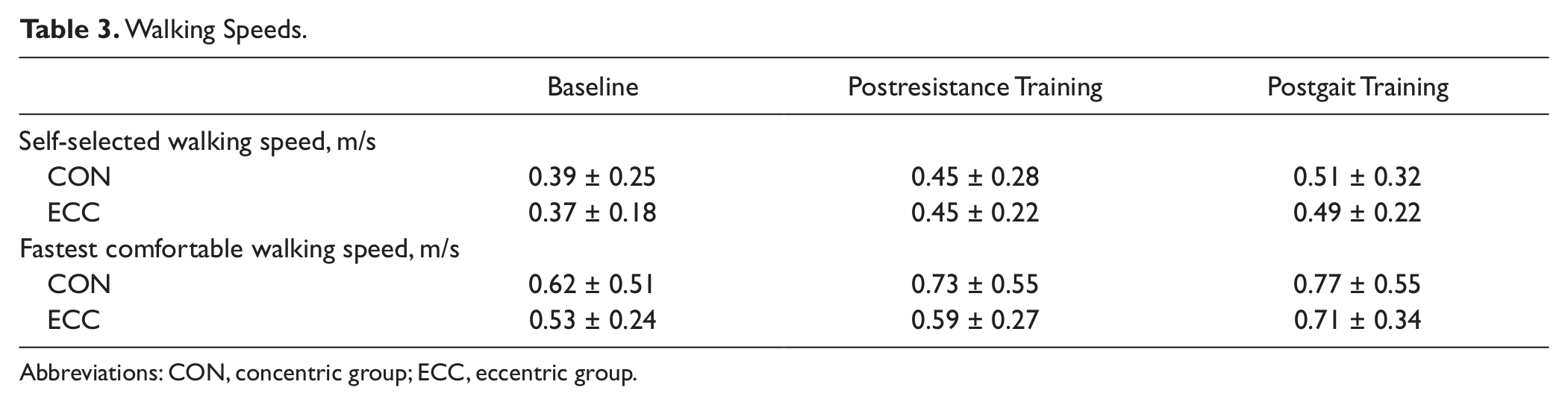
Abbreviations: CON, concentric group; ECC, eccentric group.
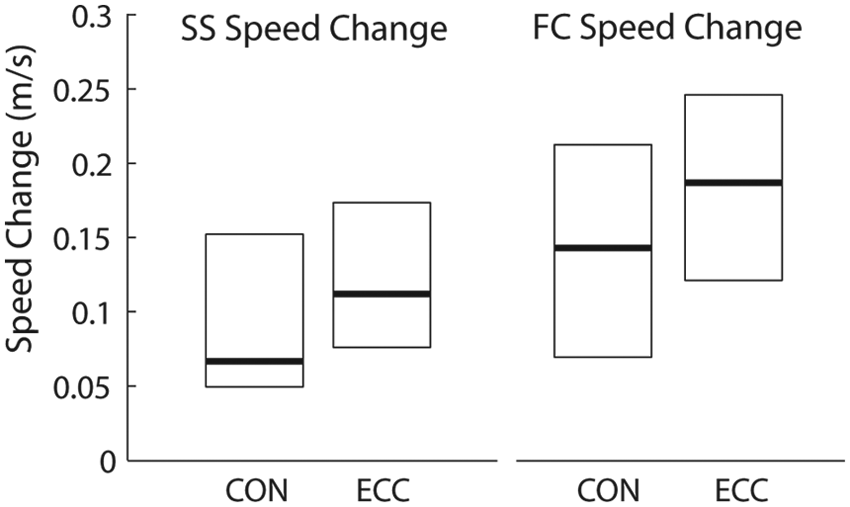
Distribution of walking speed improvements: box plots showing the distribution of walking speed improvements for the concentric (CON) and eccentric (ECC) groups. Each plot displays the median (bold line), upper quartile, and lower quartile of change in walking speed (m/s) calculated between baseline and the end of the training intervention.
Discussion
The findings of this study support the hypothesis that ECC resistance training is more effective than CON training for inducing central neural adaptation that enhances bilateral power production and walking speed after stroke. The evidence of a central neural adaptation in the ECC group includes cross-education of power, strength, and improved neuromuscular activation to the untrained nonparetic leg. Cross-education is a well-known phenomenon and has previously been demonstrated with ECC resistance training in healthy adults.20,21 In contrast, the CON group revealed no significant cross-transfer of resistance training effects, which is consistent with the findings of a recent study in adults poststroke showing that CON resistance training of the paretic leg did not influence nonparetic leg strength. 8 Cross-education is argued to be mediated by increased efficacy of central motor areas or pathways involved in control of force and movement. 34 Consistent with the hallmark characteristics of cross-education, 35 the magnitude of changes in the nonparetic leg of the ECC group were smaller and more variable than in the trained paretic leg. Further evidence of greater neural adaptation in the ECC than in the CON group is the larger improvements observed for paretic leg neuromuscular activation and power following resistance training. Indeed, power and activation demonstrated similar training responses that were specific to the contraction mode trained (ie, specificity of training21,36). Furthermore, the short 5-week duration of the resistance training phase ensures that most of the improvements in power production are mediated by neural, not muscular, factors. 34
As discussed in the Introduction, greater neural adaptation with ECC training may be a result of the unique neural demands of ECC contractions18,19 or a result of heightened contraction intensity.1,16 These explanations and our findings of increased voluntary neuromuscular activation contrast with the explanation put forth by Engardt and coworkers. 7 They proposed that larger strength gains with ECC training after stroke were a result of the effect of heightened quadriceps stretch reflex activity that added to the voluntary neural drive during ECC knee flexion contractions. However, it is not definitively known whether the observed increase in coactivation was the result of a genuine “spastic” response (eg, velocity-dependent stretch-induced activity) or an appropriate coactivation as occurs in response to higher joint torque production posttraining (ie, for joint stabilization). However, we observed that neither CON nor ECC training increased coactivation. This finding is consistent with other recent work10,38 and adds to the cumulative evidence showing that resistance training improves voluntary agonist activation without deleterious effects on antagonist coactivation poststroke.
During gait training, the paretic leg of both the CON and ECC groups maintained, but did not further advance, power production capability. Despite stable power production, paretic limb activation in RF and VM was reduced but remained higher than baseline. This relative reduction in activation may stem from the overall lower intensity of gait training relative to resistance training. High-intensity force production and neural drive39,40 are required to induce strength gains. Gait training does not appear to provide sufficiently high-intensity contractions that require high levels of neural activation. Furthermore, high-intensity resistance training promotes activation of high-threshold motor units and synchronous motor unit firing, both of which contribute increased magnitude to the surface EMG signal.41-43 Reduction in these recruitment and firing parameters may readily explain reduced EMG amplitude. Reflex potentiation of central neural drive with resistance training may also contribute to increased EMG amplitude,42,44-46 which is subsequently lost after resistance training is stopped. The major finding in the nonparetic leg during the gait training phase was that power increased modestly in the ECC group but did not change in the CON group. Although it is unclear why gait training improved power in the ECC group but not the CON group, we can speculate that the central neural adaptations experienced by the ECC group during resistance training contributed favorably to the participants’ ability to engage in and benefit from gait training.
At the conclusion of the full intervention (resistance + gait training), the training mode–specific gains in paretic leg power and activation were comparable between the CON and ECC groups. That is, improvements were similar for concentric contractions in the CON group compared with eccentric contractions in the ECC group and for eccentric contractions in the CON group compared with concentric contractions in the ECC group. However, the cross-education of increased power production in the nonparetic leg remained considerably greater in the ECC than in the CON group. Therefore, the central neural adaptation that is evidenced by cross-education was a robust and persistent benefit of eccentric resistance training.
Lower-extremity muscle strength is associated with walking performance, which indicates a potential benefit for resistance training in rehabilitation of walking performance after stroke. 47 However, previous investigations of the effect of resistance training interventions on walking function have yielded inconclusive results. Some studies have reported significant, albeit modest, improvements in walking speed or function,8,10,48 whereas other have not.49-51 Our clinic-based gait training intervention emphasized motor learning (eg, whole and partial task practice and knowledge of performance 28 ) to promote acquisition of a more appropriate gait pattern and faster gait speed. Over the course of the full intervention, we found that a higher proportion of ECC-trained individuals responded to the intervention. Indeed, both the median improvement in walking speed and the number of clinically defined responders in the ECC group were almost 2-fold greater than in the CON group. Despite the absence of a difference between groups for mean walking speed improvement, this finding of a larger number of responders in the ECC group is an important indicator of a beneficial effect of eccentric resistance training. We propose that the higher proportion of responders in the ECC group was facilitated by the bilateral improvements in leg power production, which generalized to improvement at the behavioral level.
There are some methodological limitations in this study. The use of surface EMG to assess changes in neuromuscular activation with training is a common6,8,52,53 but imperfect approach because the variability of EMG across sessions is affected by a variety of factors. 54 In some cases, this variability may have precluded the ability to detect significant changes. In general, however, we found agreement between the power and EMG measurements, which supports the important role of neural adaptation in contributing to increased strength and walking function poststroke. A second methodological issue is that we did not measure the size or contractile properties of muscle and thus cannot account for any potential muscular responses that may have occurred over the course of the 8-week intervention.
In conclusion, eccentric and concentric resistance training produce modality-specific adaptations in both power production and EMG modulation. Eccentric resistance training was the more effective training mode for inducing central neural adaptation, as indicated by cross-education of resistance training effects. As a result, eccentric training appears to enhance the likelihood that the benefits of resistance training will generalize to improved walking speed.
Footnotes
Acknowledgements
We thank Christine Dairaghi, Theresa McGuirk, and Andrew Bauer for their contributions to data analysis and C. Maria Kim, PT, MSc, Jody Dozono, MPT, Abigail Andrade, and Jeff Jaramillo, PT, MS, for conducting participant interventions and testing.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the US Department of Veterans Affairs Rehabilitation Research and Development Service (B7176W to DJC; B29792R and F7823S to CP).
*
This phase III clinical trial was registered with the Research and Development Committee of the VA Palo Alto Health Care System and the VA Rehabilitation Research and Development Service.