
Abstract
Microvascular complications are now recognized to play a major role in diabetic complications, and understanding the mechanisms is critical. Endothelial dysfunction occurs early in the course of the development of complications; the precise mechanisms remain poorly understood. Mitochondrial dysfunction may occur in a diabetic rat heart and may act as a source of the oxidative stress. However, the role of endothelial cell-specific mitochondrial dysfunction in diabetic vascular complications is poorly studied. Here, we studied the role of diabetes-induced abnormal endothelial mitochondrial function and the resultant endothelial dysfunction. Understanding the role of endothelial mitochondrial dysfunction in diabetic vasculature is critical in order to develop new therapies. We demonstrate that hyperglycaemia leads to mitochondrial dysfunction in microvascular endothelial cells, and that mitochondrial inhibition induces endothelial dysfunction. Additionally, we show that resveratrol acts as a protective agent; resveratrol-mediated mitochondrial protection may be used to prevent long-term diabetic cardiovascular complications.
Introduction
Cardiovascular (CV) complications account for a substantial proportion of the morbidity and mortality that occurs in patients living with diabetes. In particular, the development of endothelial dysfunction is thought to play a major role in the pathogenesis of diabetes-associated cardiovascular disease (CVD) in addition to other pathology including retinopathy and nephropathy.1,2 The prevention and early identification of vascular complications are a central issue in the care of patients with diabetes. Recent data suggest that endothelial dysfunction occurs very early in the course of the development of vascular pathology. For example, these features have been reported in preadolescent children with type I diabetes, 3 and other studies have also reported the increased presence of markers of endothelial activation and perturbation along with carotid intima–media thickening, 4 in early type I diabetes. Thus, understanding the mechanisms of early endothelial dysfunction in diabetes is critical for prevention of long-term complications.
Several crucial factors, such as alterations in oxidative stress, lipid metabolism, insulin resistance and inflammation, have been suggested to play a role in the causation of endothelial dysfunction. The precise source of the oxidative stress remains controversial, and potential sources include nicotinamide adenine dinucleotide phosphate (NADPH) oxidases and mitochondria. Interestingly in the context of inflammation, mitochondrial sources of reactive oxygen species (ROS) have been proposed to be particularly important. 5 Consistent with this concept, mitochondrial function has been shown to be impaired in the rat heart, providing a possible explanation for the pathogenesis of diabetic cardiomyopathy, 6 although the possibility that abnormal endothelial mitochondrial function is a specific contributor has not been examined in detail.
In this study, we hypothesized that diabetes induces abnormal endothelial mitochondrial function, and that this contributes to endothelial dysfunction in diabetes. Given the relatively low abundance of mitochondria in endothelial cells (ECs), it had been assumed that they were unlikely to contribute significantly to cellular function. However, it is now apparent that in addition to adenosine triphosphate (ATP) generation, mitochondria also play a role in EC homeostasis via modulation of nitric oxide (NO), ROS and Ca2+. 7 Stimulation of mitochondrial Ca2+ uptake can activate NO production that in turn can modulate mitochondrial Ca2+ uptake and efflux, demonstrating a negative feedback regulation. 8 Thus, understanding the role of EC-specific mitochondrial dysfunction in diabetic vasculature is critical in order to develop new therapies.
Our data show that coronary perfusion is reduced early in the course of experimental diabetes. To understand the underlying cause of reduced microvascular function in diabetic heart, we examined the role of mitochondrial dysfunction as a cause for endothelial dysfunction. We demonstrate that hyperglycaemia leads to mitochondrial dysfunction in microvascular ECs, and that mitochondrial dysfunction induces endothelial dysfunction that manifests as impaired pharmacologic and regenerative capacity. Finally, we showed that resveratrol exerts protective actions in the setting of diabetes-associated endothelial dysfunction.
Methods
Animal model
All experiments were carried out in accordance with the guidelines set forth by the institutional animal ethics committee. Hyperglycaemia was induced in pathogen-free male Sprague-Dawley rats (200–250 g) with a single dose of streptozotocin [STZ, 65 mg/kg intraperitoneal (i.p.) prepared fresh in citrate buffer pH 4.5]. Control animals received equal volume of the citrate buffer. Blood glucose level was determined at each time point using Optium Omega (Abbott Diabetes Care, Alameda, CA) clinical blood glucose monitor (Figure 8; supplementary data available online). Animals with blood glucose level of <200 mg/dL at day 2 were excluded from the study. Animals were housed in a cage rack system with continuous air circulation (~50 air changes/h) for the duration of the studies with daily cage changes. Animals were studied longitudinally at 0, 7 and 14 days post-STZ (n = 6 per time point). Age-matched rats were treated with vehicle and studied in parallel.
Resveratrol treatment was initiated after confirmed hyperglycaemia 2 days after the STZ injection. The diabetic animals were randomly assigned to receive daily subcutaneous (s.c.) injection of 10 mg/kg/day of resveratrol or equal volume of vehicle [40% dimethyl sulphoxide (DMSO)]. Resveratrol stock was prepared at 10 mg/mL. The desired dose of resveratrol was 10 mg/kg/day. The injection volume was 0.1 mL/100 g. So, for a 250-g rat, the injection volume was 0.25 mL. Vehicle was also injected at 0.1 mL/100 g in rats. Overall, injection volumes varied between 0.2 and 0.25 mL in all animals. In the pilot studies, vehicle injection for 14 days did not result in any cardiac functional abnormalities (Figure 9; supplementary data available online).
Endothelial cell studies
Single-donor cardiac human microvascular endothelial cells (HCMVECs) were obtained from PromoCell GmbH (Germany). Cells were cultured in recommended endothelial growth media (EGM) with 5.5 mM glucose. For hyperglycaemic conditions, EGM was supplemented with glucose to achieve a final concentration of 25 mM.
Measurements of CV performance
Langendorff isolated heart preparation was used for measurement of myocardial function and coronary flow measurements in rats. The perfusion solution (modified Krebs–Hanseleit buffer) will be equilibrated with gas (95% O2 + 5% CO2) and warmed to 38°C. After anaesthetizing with sodium pentobarbital, the animal was fixed on an operating table, and the chest wall was rapidly dissected. After carefully removing the pericardium, the ascending aorta was isolated and cut rapidly just underneath the first branch. The heart was immediately mounted on a primed cannula and perfusion started at 7 mL/min. MikroTip Millar catheter is inserted through a small incision at the base of left atrium and secured in place by a surgical suture. Electrocardiography (ECG) electrodes are inserted in the outer wall of the ventricle. Continuous left ventricular (LV) pressure and ECG are recorded using LabChart v.7 (ADInstruments, New South Wales, Australia).
Measurement of coronary perfusion rate
During the functional analysis, the flow of perfusion was maintained at 7 mL/min, and pressure was maintained at 80 mmHg. After initial 15-min equilibration period, the outgoing perfusate was collected in a graduated beaker over 10 min, and the coronary perfusion rate was calculated in millilitre per minute.
Mitochondrial function in cultured ECs
Mitochondrial function in cultured HCMVECs was measured using Seahorse XF analyser (Seahorse Bioscience, MA, USA). Cells were exposed to normal glucose (NG; 5.5 mM glucose), high glucose (HG; 25 mM glucose), normal glucose + 20 µM resveratrol (NG + RESV) or high glucose + 20 µM resveratrol (HG + RESV) for 24 h prior to mitochondrial functional measurement. Mitochondrial oxygen consumption rate (OCR) within the treatment groups was measured using standard protocols (Figure 1). Briefly, OCR was measured after serial addition of mitochondrial inhibitors and uncouplers [oligomycin, carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) and antimycin-A]. Oligomycin is an inhibitor of ATP synthase by blocking its proton channel, and it is used to prevent state 3 respiration. FCCP is a potent reversible inhibitor of mitochondrial oxidative phosphorylation (OXPHOS). Antimycin-A is also an inhibitor of OXPHOS. It disrupts the proton gradient across the inner membrane and inhibits ATP production.
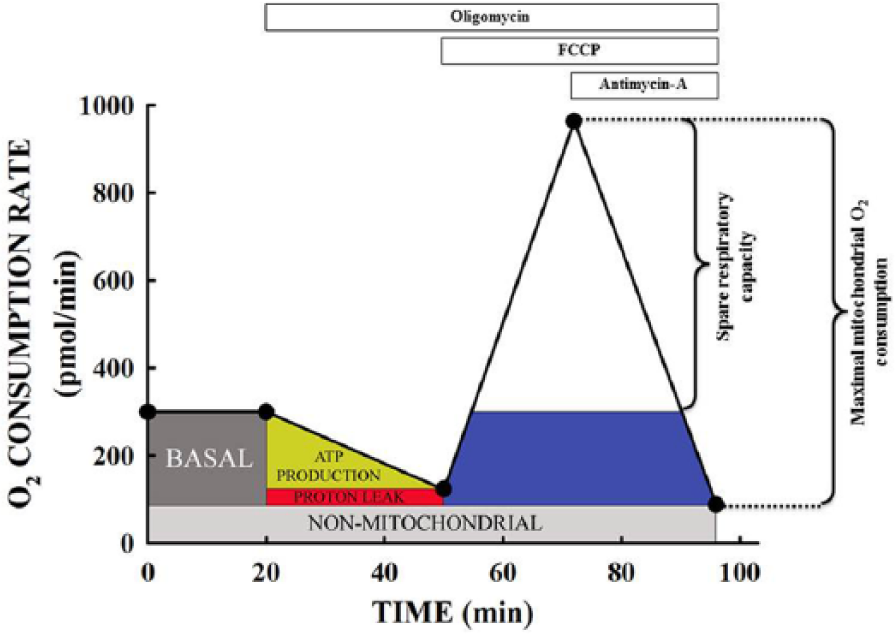
A representative graph showing mitochondrial oxygen consumption rate (OCR) in human coronary microvascular endothelial cells using Seahorse XF24 analyser. OCR was measured after the addition of oligomycin, FCCP and antimycin-A. Mitochondrial oxygen consumption linked to various respiratory processes is calculated as shown in the graph.
Isolated aortic rings
Mice were euthanized, and the thoracic aorta was excised and placed into ice-cold Kreb’s modified solution (composition in mM: NaCl 119, KCl 4.7, MgSO4·7H2O 1.17, NaHCO3 25, KH2PO4 1.18, CaCl2 2.5, glucose 11 and ethylenediaminetetraacetic acid (EDTA) 0.03). The adipose and connective tissues were removed, and 2-mm-length segments were mounted in the myograph. After 1 h of equilibration, vessels were contracted with Potassium physiological salt solution (KPSS) (KPSS in mM; KCl 123, MgSO4·7H2O 1.17, NaHCO3 25, KH2PO4 1.18, CaCl2 2.5, glucose 6.05 and EDTA 0.03), a potassium depolarizing solution to assess vessel viability. Responses to vasodilators were then examined in arteries preconstricted to ~50% KPSS with cirazoline (10–100 nmol/L). Full concentration–response curves to acetylcholine (1 nM–100 µM) and sodium nitroprusside (0.1 nM–100 µM) were constructed. Separately, full concentration–response curves to acetylcholine were also constructed in the absence and presence of rotenone (2 µM) or rotenone + resveratrol (10 µM). For these experiments, vessels were pre-incubated with rotenone or rotenone + resveratrol for 30 min prior to measuring responses to ACh. All vasorelaxation responses were expressed as percent relaxation from the preconstriction response. Variable slope sigmoidal concentration–response curves to each agonist were fitted and graphed, and the −log EC50 M value (i.e. the concentration giving 50% of the maximum response) was calculated for individual curves using GraphPad Prism (v 5.0).
In vitro scratch assay
The in vitro scratch assay is an easy, low-cost and well-developed method to measure EC function.
9
Eight-well chamber slides (Nunc Corporation, Rochester, NY, USA) were coated with 50 µg/mL poly-
Statistical analysis
All data are presented as mean ± standard error of the mean (SEM). All data from cell experiments are shown from 9 to 12 replicate measurements. Differences between treatment groups were assessed using one-way analyses of variance, with post hoc Dunnett’s tests (comparisons against control values) to evaluate significant comparisons. A p value of < 0.05 is described as statistical significance.
Results
Hyperglycaemia induces endothelial mitochondrial dysfunction
Using HCMVECs, we demonstrate that mitochondrial function is significantly impaired in cells exposed to hyperglycaemia (25 mM glucose) for 24 h (Figure 2). ATP-linked mitochondrial respiration, maximal respiration and spare respiratory capacity were significantly lower in cells exposed to hyperglycaemia. By contrast, cells treated with resveratrol demonstrated improved mitochondrial function parameters as measured by Seahorse XF analyser.
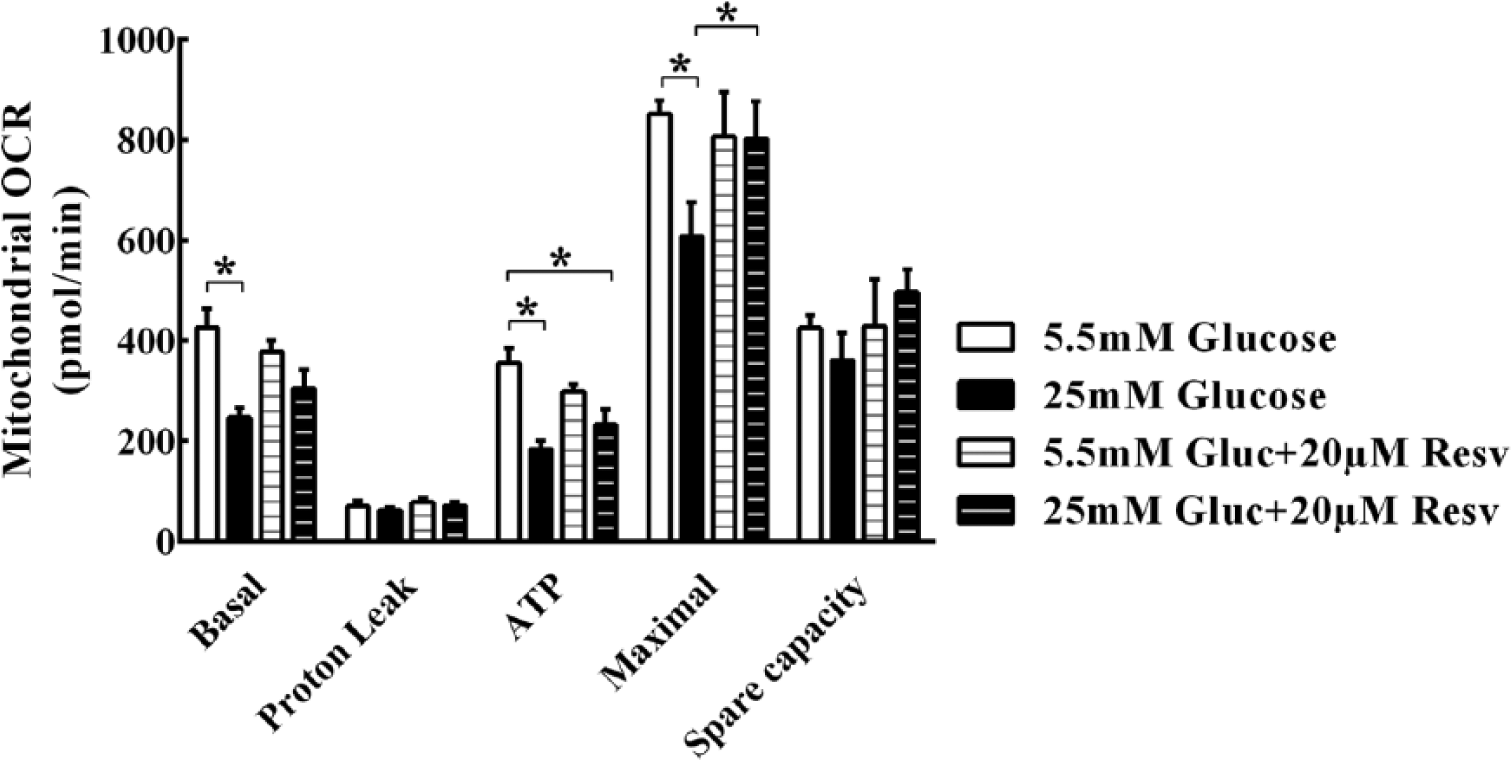
Mitochondrial oxygen consumption rate (OCR) measured in human coronary microvascular endothelial cells using Seahorse XF24 analyser. OCR was measured after the addition of agents to enhance or block mitochondrial respiration. Exposure to hyperglycaemia significantly impairs mitochondrial function. ATP production–linked oxygen consumption, maximal respiration and spare respiratory capacity were significantly lower in cells exposed to hyperglycaemia. Resveratrol treatment in the presence of hyperglycaemia rescued mitochondrial function in these endothelial cells. All data are expressed as mean ± SEM in five replicates from three different experiments for each group.
Mitochondrial inhibition impairs vascular reactivity
Given the impact of hyperglycaemia on endothelial mitochondrial function, we next aimed to determine whether impaired mitochondrial function directly influences vascular tone. As shown in Figure 3, inhibition of mitochondrial function with the complex I inhibitor, rotenone, significantly impaired the endothelium-dependent modulation of vascular tone by acetylcholine. Moreover, consistent with the above observations, resveratrol treatment reverses the impairment caused by rotenone in this setting (EC50 −log10 [ACh] (M): (Vehicle) VEH; ROT; ROT + RESV: −7.40 ± 0.13; −7.01 ± 0.07*‡; −8.18 ± 0.12: *p < 0.05 vs control, ‡p < 0.05 vs ROT + RESV). Isolated vessel responses to sodium nitroprusside were not different among the treatment groups.
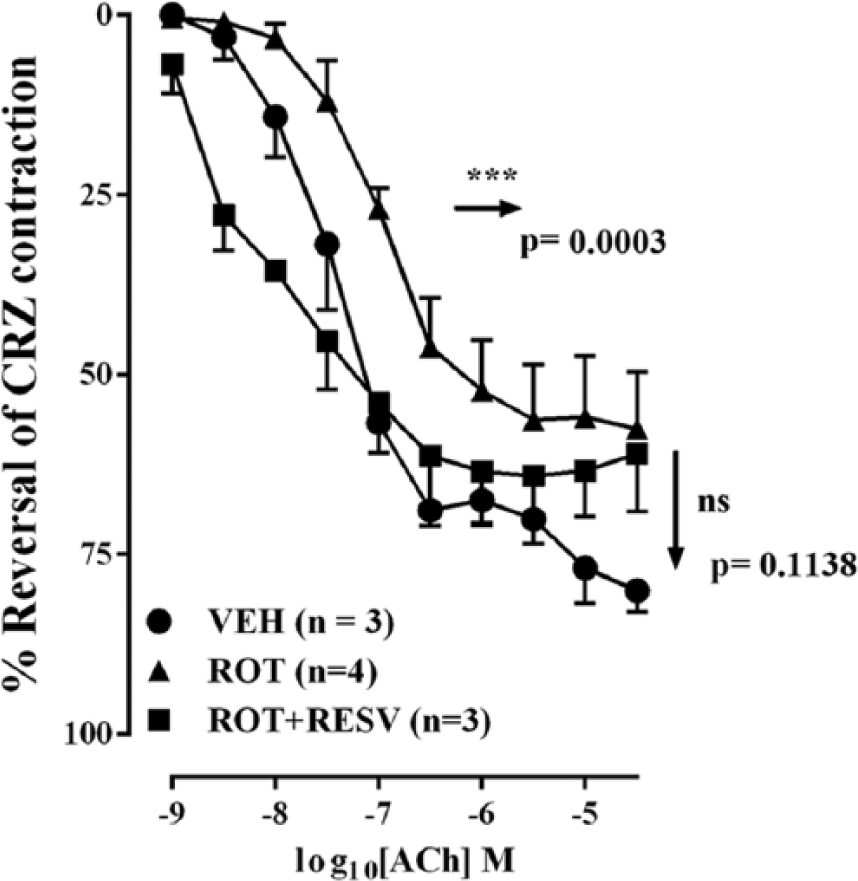
Endothelium-dependent isolated vessel function measured from mouse aortas. Inhibition of mitochondrial function with the complex I inhibitor, rotenone, significantly impaired the endothelium-dependent modulation of vascular tone by acetylcholine. Incubation of isolated vessels with resveratrol reverses the impairment caused by rotenone in this setting. All data are expressed as mean ± SEM for each group.
Mitochondrial dysfunction causes impaired endothelial proliferation
In addition to the pharmacologic consequences of abnormal mitochondrial function in ECs, we also aimed to test the possibility that endothelial mitochondrial dysfunction could impair EC proliferation and regeneration. Using the in vitro scratch assay, we demonstrated that inhibition of mitochondrial function by rotenone causes impaired EC migration. Similarly, EC migration was also significantly slower in the HG-treated group and the rotenone-treated group (Figure 4). Scratch width after 12 h was 21.1% ± 2.3% of the baseline width in NG, while repair was markedly attenuated (99.1% ± 6.3%) in HG (p < 0.05 vs control) and 93.19% ± 10.4% in the normal glucose + rotenone–treated group (p < 0.05 vs control). Resveratrol treatment in these groups significantly improved EC migration and scratch closure in 12 h post-scratch.
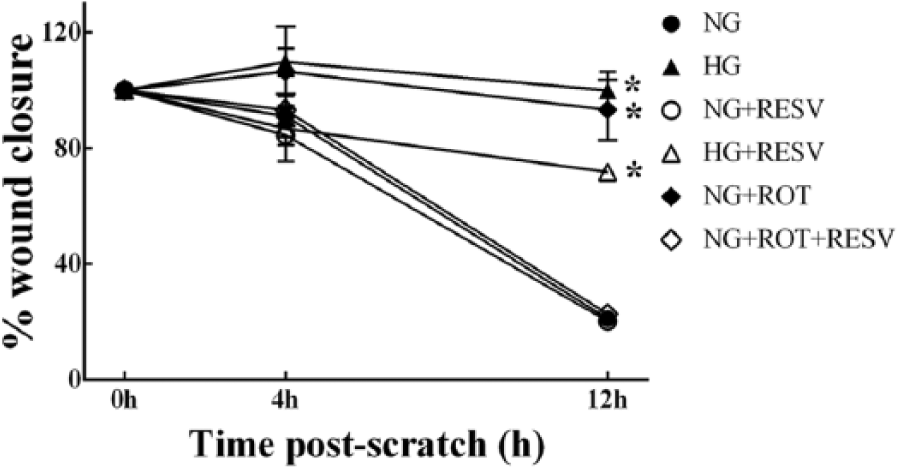
Effects of mitochondrial inhibition on endothelial cell proliferation or migration were measured using the in vitro scratch assay. These data demonstrate that inhibition of mitochondrial function by rotenone causes impaired endothelial cell proliferation and/or migration. ‘Wound closure’ was significantly slower in the high glucose–treated group and the rotenone-treated group. Resveratrol treatment in the high glucose–treated and rotenone-treated groups partially improved endothelial cell migration and scratch closure in 12 h post-scratch. All data are expressed as mean ± SEM in three replicates for each group.
Reduced coronary perfusion in diabetic animals
In order to extend the above observations to the in vivo situation, we next aimed to investigate whether ECs exposed to chronic hyperglycaemia would exhibit abnormal function. In this study, we examined coronary flow in control and diabetic animals. In the diabetic animals, the coronary flow rate was significantly lower than in the control animals, and this was significant reversed by treatment with resveratrol (Figure 5).
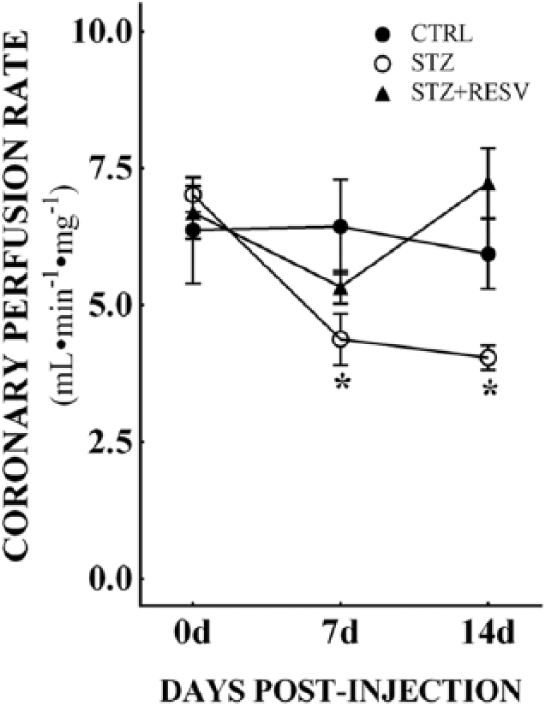
Coronary perfusion changes in diabetic animals were measured using Langendorff isolated heart preparation. Coronary flow was measured in control animals, streptozotocin (STZ)-induced diabetic animals and diabetic animals treated with resveratrol. Coronary flow was measured at 7 and 14 days post-injection. Coronary flow was significantly reduced in diabetic animals (n = 6) when compared to control animals (n = 7). Resveratrol treatment in diabetic animals significantly improved coronary perfusion in these animals (n = 6). All data are expressed as mean ± SEM.
Early and sustained contractile dysfunction in diabetic hearts is alleviated by resveratrol treatment
Having demonstrated abnormal coronary flow in the diabetic rat heart, we also examined effects on mechanical function. STZ-treated animals showed contractile deficits as early as 7 days post-STZ, with a 26% reduction in LV dP/dtmax compared to the vehicle control group (Figure 6(a)). Concomitantly, diastolic function (LV dP/dtmin) was also impaired in the diabetic animals by 21% at 7 days (Figure 6(b)). Daily s.c. injection with resveratrol (10 mg/kg) alleviated the contractile dysfunction observed in diabetic animals. Impaired contractile function was also evident in the 14-day treatment group, again with significant improvement after resveratrol treatment. These data demonstrate that cardiac functional deficits detected at 7 days post-STZ are sustained during progression of diabetes and are alleviated by resveratrol treatment. In addition to the contractile effects, the QRS and QTc intervals were also abnormal in diabetic animals. Resveratrol treatment normalized both QRS (Figure 6(c)) and QTc intervals (Figure 6(d)). Notably, there was a significant (p = 0.03) correlation between the degree of impairment in coronary flow and contractile function (Figure 7) consistent with the in vitro studies of the effects of mitochondrial oxidative stress on EC function.
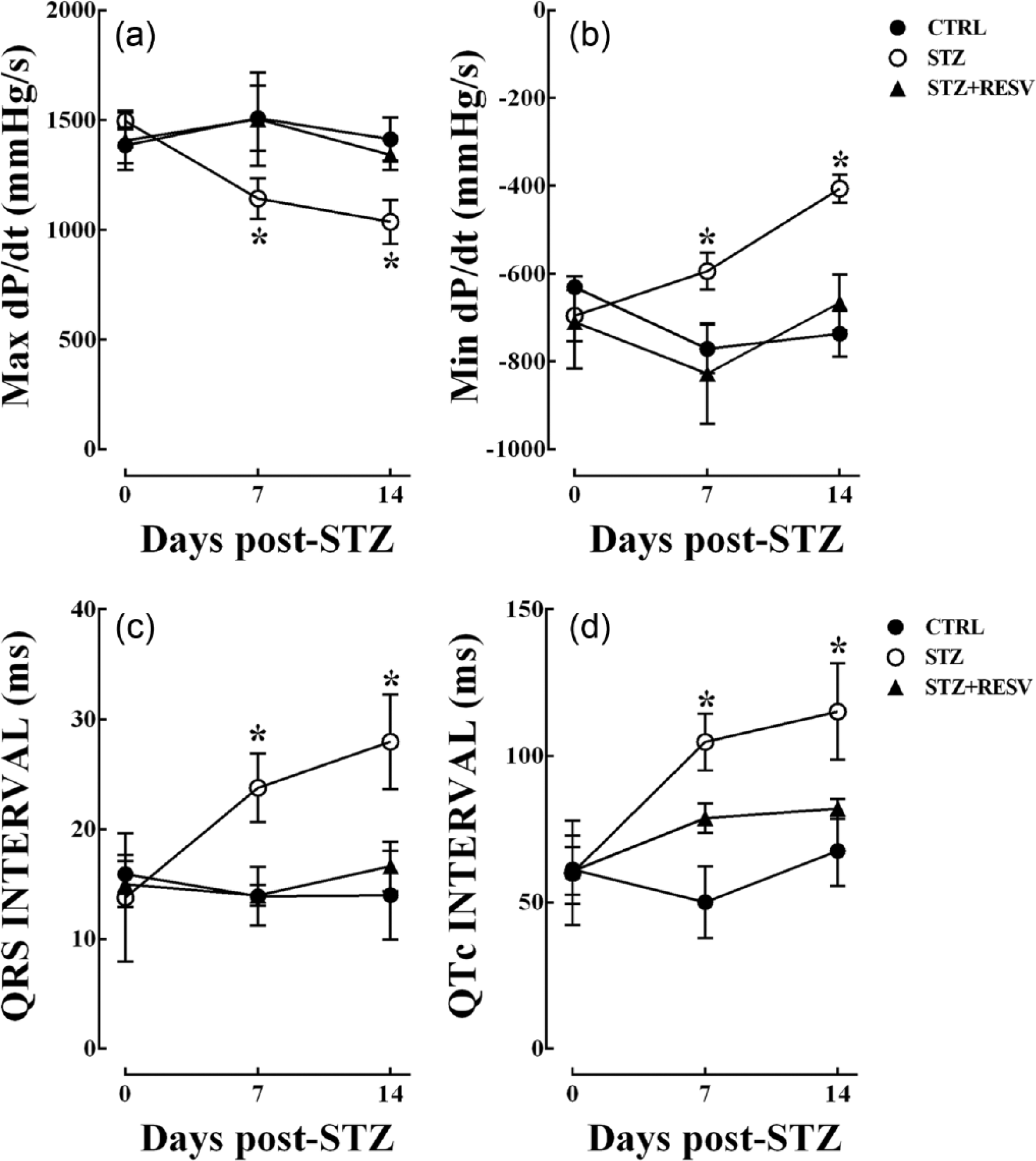
Cardiac contractile and electrophysiological function measured in isolated rat hearts. We observed early and sustained cardiac dysfunction in diabetic hearts as early as 7 days post-STZ. STZ-treated animals showed contractile deficits with impaired (a) systolic and (b) diastolic functions. Daily resveratrol treatment alleviated the contractile dysfunction observed in diabetic animals. Additionally both (c) QRS and (d) QTc intervals were elongated in diabetic animals, suggesting that electrophysiological disturbances occur early in diabetes progression. Daily injection of resveratrol normalized the cardiac electrophysiological parameters. All of these impairments were observed at 14 days post-injection, indicating that these abnormalities are sustained in diabetes. Resveratrol treatment alleviated these impairments in cardiac function at both time points measured. n = 6–7 per group, and all data are expressed as mean ± SEM.
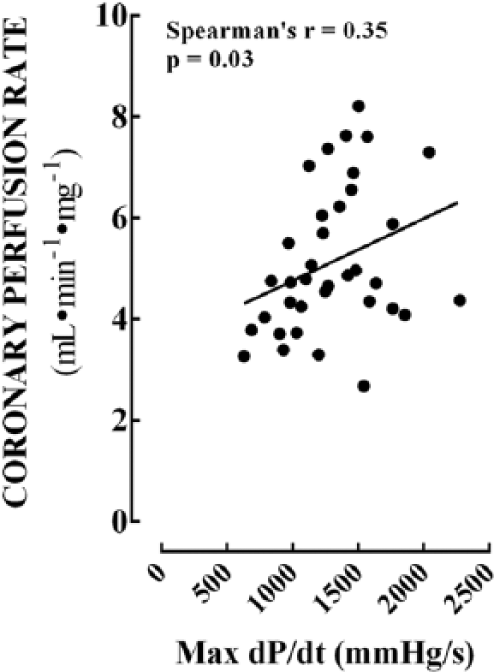
Association between coronary perfusion and cardiac contractile function. We demonstrated that a significant (p = 0.03) correlation exists between the degree of impairment in coronary flow and contractile function. These data indicate that endothelial dysfunction mediated via mitochondrial impairments may directly affect cardiac contractility during diabetic cardiovascular complications.
Discussion
In this study, we examined the influence of mitochondrial dysfunction on a range of fundamental CV properties modulated by the vascular endothelium. Although mitochondria only occupy an estimated 5% of EC volume and ECs preferentially utilize non-mitochondrial pathways to generate ATP, it is increasingly accepted that endothelial mitochondria contribute to other important aspects of endothelial biology. Among these, emerging evidence is accumulating to suggest that endothelial mitochondria contribute vascular homeostasis via modulation of nitric oxide (NO), ROS generation and intracellular Ca2+ levels. 7 In the context of diabetes, previous studies have indicated that hyperglycaemia can elevate mitochondrial ROS production. 10 We show in this study that hyperglycaemia also impairs mitochondrial energy metabolism as reflected by reduced basal oxygen consumption, ATP synthesis and maximal respiratory capacity.
Mitochondrial control of cellular metabolic demands is centrally coordinated by the proliferator-activated receptor gamma co-activator 1α (PGC-1α), which is regulated by sirtuin-1 (SIRT1) via deacetylation. 11 SIRT1 is the most widely studied sirtuin homologue and is shown to play an important role in biological functions ranging from DNA repair to longevity. 12 It is widely recognized that SIRT1 plays a role in cellular energy metabolism. 13 Studies have also shown that SIRT1 promotes endothelium-dependent relaxation via stimulation of nitric oxide synthase 3 (NOS3) and NO. 14 SIRT1 is also known to play a critical role in angiogenesis with upregulation of SIRT1 leading to increased expression of vascular endothelial growth factor (VEGF) and NOS3. 15 Thus, SIRT1-mediated signalling may affect the mitochondrial function and may be responsible for endothelial dysfunction observed in diabetic vasculature. Resveratrol is a known SIRT1 activator which prevents or delays the onset of many diseases such as cancer, ischaemic heart disease and other disease models.16 –18 Resveratrol-mediated SIRT1 activation is known to regulate PGC-1α deacetylation and thus enhance mitochondrial biogenesis and function. 19 Alternatively, SIRT1 is also recently shown to influence mitochondrial biogenesis as well as OXPHOS via PGC-1α-independent pathway which involves mitochondrial transcription factor A (TFAM)-mediated signalling. 20 These resveratrol-activated SIRT1 pathways may directly affect mitochondrial function in response to hyperglycaemia. Accordingly, we examined the influence of resveratrol on the hyperglycaemia-induced inhibition of mitochondrial function in ECs. Consistent with the above paradigm, resveratrol was able to reverse the effects of hyperglycaemia on mitochondrial function in ECs.
Having demonstrated the depressive effects of hyperglycaemia on endothelial mitochondrial function, we examined the impact of mitochondrial dysfunction upon control of vascular tone and endothelial proliferation. In isolated aortic rings, we showed that the complex I inhibitor impaired responses to the endothelium-dependent vasodilator acetylcholine but not to sodium nitroprusside. Endothelium-dependent nitric oxide (NO) production is the primary mediator of ACh-mediated responses. Differences in ACh responses after rotenone treatment indicate that endothelial NO production may be diminished. Responses to single nucleotide polymorphism (SNP) were not altered by rotenone alone or rotenone + resveratrol, suggesting that smooth muscle responses are not altered in this setting. While we did not specifically measure levels of oxidative stress in this preparation, resveratrol was able to reverse the effects of rotenone, consistent with the possibility that the inhibitory effect of rotenone on endothelial vasodilator function was mediated via mitochondrial oxidant stress. Some prior studies have demonstrated that resveratrol can promote vascular relaxation via potassium channels. However, these responses are observed at higher concentration of resveratrol than the concentration used in our studies. 21 Hence, it is highly unlikely that the protective effects observed in our studies are due to direct vessel relaxation by resveratrol. In a complimentary manner, we also showed that both high glucose and rotenone markedly impaired the endothelial proliferation response in the wound scratch model. We did not specifically test for the presence of apoptosis; however, it is known that hyperglycaemia can induce apoptosis.22,23 Here, we did not specifically measure apoptotic cell or tissue status as previous studies have demonstrated that rotenone-induced apoptosis is primarily evident at 24–36 h post-treatment.24 –26 In our in vitro studies, we have exposed the cells up to only 12 h of rotenone treatment. Hence, it is less likely that the effects observed in the scratch assay are due to apoptosis in these studies. Future studies with longer duration of rotenone treatment and simultaneous measurement of cellular apoptosis are warranted. Our study provides new insights into the link between mitochondrial dysfunction and apoptosis, highlighting a potential central role for mitochondrially generated ROS given the protective actions of resveratrol.
Although a number of studies provide indirect evidence for altered mitochondrial function in diabetic patients, the relative contributions of abnormal mitochondrial function in cardiomyocytes and ECs to the overall physiology of the diabetic heart have not been examined. Similarly, while a reduction in diastolic function that is correlated with the magnitude of the reduction in the cardiac phosphocreatine–ATP ratio 27 is consistent with mitochondrial dysfunction, it is possible that this may be only one measure of the effect of mitochondrial dysfunction. Moreover, prior studies have linked mitochondrial dysfunction to the pathogenesis of diabetic vascular disease via mitochondrial production of ROS, impaired biogenesis and increased markers of inflammation. 28 Our data indicate that diabetes induced by STZ is accompanied by significant impairments in systolic and diastolic functions together with coronary flow. Consistent with the above studies, resveratrol reversed the myocardial and coronary vascular phenotype, suggesting that abnormal cardiomyocyte and endothelial mitochondrial function may contribute to the diabetic cardiomyopathy phenotype.
Taken together, our data demonstrate that resveratrol treatment has beneficial effects on myocardial function, coronary perfusion, EC function and vascular tone. As such, these studies suggest that SIRT1 activation by resveratrol may be beneficial for reducing CV abnormalities observed in diabetic heart and thus reduce the risk of diabetic CV complications. In addition to the direct regulation of mitochondrial energetics, it is also evident that mitochondrial biogenesis is a critical mediator of cellular respiration including that seen in ECs. 29 Fragmentation of mitochondrial networks has been shown after exposure to hyperglycaemia and oxidants. 30 Thus, alterations in dynamic mitochondrial regulation due to hyperglycaemia and oxidative stress may lead to vascular dysfunction. Interventions such as resveratrol, directed towards restoring normal dynamics, via PGC-1α-SIRT1 signalling may have significant therapeutic benefits in this setting. Based on our studies, further studies to investigate the role of EC-specific mitochondrial changes as a mediator of microvascular abnormalities in diabetic CVD are warranted.
The annual incidence of type 1 diabetes is increasing worldwide at a rate of approximately 3%–5%, particularly in children under the age of 15 years, 31 and early onset type II diabetes is also increasingly recognized. Early onset increases the likelihood of long-term complications including vascular disease and mortality. Vascular complications including myocardial infarction, stroke and peripheral vascular disease account for a substantial proportion of the life-threatening events experienced by patients living with both type I and type II diabetes. Conventional risk factors for the development of vascular complications including poor disease control, dyslipidemia, hypertension and smoking are well known; however, despite the development of improved strategies for glucose and lipid control, the risk of vascular complication remains unacceptably high. Mechanistically, heightened oxidative stress and endothelial dysfunction have been proposed to be major contributors to the development of micro- and macrovascular complications, although the basis for the transition of endothelial dysfunction to vascular pathology remains unclear. Our studies demonstrate that CV dysfunction is evident in the early phase of diabetes. These effects observed within 7 days post-STZ are sustained over the 14-day period in this study. Contractile as well as electrophysiological abnormalities are seen in the diabetic animals. Resveratrol treatment of the diabetic animals demonstrated significant improvement in functional parameters studies. Early studies of cardiac biopsy samples showed significantly higher incidence of small vessel disease in diabetic patients. 32 The microvasculature is dynamically regulated by a combination of endogenous pro- versus anti-angiogenic forces,33 –35 including EC impairment, decreased local presence of survival factors and increased local presence of anti-angiogenic factors.33 –36 Decreased coronary reserve is a well-recognized contributor to the early incidence of cardiac abnormalities in diabetic patients. 37 Microvascular changes may lead to reduced perfusion and mismatch of myocardial supply and demand. 38 Indeed, our data demonstrate that coronary perfusion is reduced in diabetic animals. These changes were in the absence of any changes in heart weight and significantly correlated to impaired cardiac contractility.
Thus, our studies demonstrate that coronary microvascular dysfunction is likely to occur in the early stages of diabetes. These changes are likely to be mediated by mitochondrial dysfunction. Currently, very limited clinical data are available on the early cardiac and vascular impairments soon after diabetes diagnosis. Based on our preclinical studies, it is reasonable to believe that it would be highly beneficial in a clinical setting to carry out preliminary non-invasive CV function testing soon after diagnosis. Further studies into the potential of resveratrol as a possible therapeutic agent to delay and/or prevent these CV impairments and to normalize mitochondrial function are warranted.
Footnotes
Declaration of conflicting interests
The authors declare that there is no conflict of interest.
Funding
This research was supported by funding from the National Health and Medical Research Council of Australia (to D.M.K.) and by a Victorian Government OIS Infrastructure Grant to Baker IDI.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.