
Abstract
Aim:
We tested the hypothesis that short-term oral sodium nitrite supplementation would improve vascular dysfunction in obese, diabetic mice.
Methods and results:
Vascular function was determined in control mice and in db/db mice receiving drinking water with or without sodium nitrite (50 mg/L) for 5 weeks. Nitrite supplementation increased plasma nitrite concentrations in db/db mice (0.19±0.02 µM vs 0.80±0.26 µM; p < 0.05). Db/db mice had lower endothelium-dependent dilation (EDD) in response to increasing doses of acetylcholine versus heterozygous control mice (71.2% ± 14.3% vs 93% ± 7.0%; p < 0.05), and sodium nitrite supplementation restored endothelium-dependent dilation to control levels (92.9% ± 2.3% vs 93% ± 7.0%; p < 0.05). The improvement in endothelial function was accompanied by a reduction in intrinsic stiffness, but not by alterations in plasma or vascular markers of inflammation.
Conclusion:
These data suggest that sodium nitrite may be a novel therapy for treating diabetes-related vascular dysfunction; however, the mechanisms of improvement are unknown.
Introduction
Type II diabetes (T2D) is associated with increased cardiovascular disease (CVD) and mortality. 1 Two alterations that occur within the vasculature of type II diabetics, endothelial dysfunction and large artery stiffness, contribute to the development of CVD. 1
Endothelial dysfunction is mediated in large part by reduced bioavailability of the vascular protective molecule, nitric oxide (NO). Loss of NO bioavailability also contributes to the development of arterial stiffness. 2 Interventions that boost NO bioavailability have tremendous clinical relevance for treating vascular dysfunction in settings of obesity or T2D. 3
The oxidation of NO yields nitrite, which is now recognized as an important storage form of bioavailable NO. 4 Exogenous nitrite administration has been shown to protect against cardiovascular abnormalities, including vascular dysfunction in the settings of ageing and hypercholesterolaemia.5,6 In this study, we examined whether sodium nitrite supplementation would improve vascular dysfunction in a mouse model of T2D.
Methods
Animals and experimental design
C57BLKS/J mice (3–4 months of age) homozygous for a point mutation in the gene for the leptin receptor (db/db), and their heterozygous littermates (CON), were obtained from The Jackson Laboratory (Bar Harbor, ME, USA). Mice were randomly assigned to one of three weight-matched groups: (1) heterozygous controls (CON; n = 7–13), (2) homozygous controls (Db-C; n = 9–13), and (3) homozygous nitrite (Db-N; n = 4–13). CON and Db-C received regular drinking H2O, whereas Db-N received water supplemented with 50 mg/L sodium nitrite (NaNO2; Sigma-Aldrich, St. Louis, MO, USA) ad libitum for 5 weeks. Fresh NaNO2-supplemented water was measured and replaced every other day. All mice were allowed chow ad libitum. All procedures involving mice were reviewed and approved by the Colorado State University Institutional Animal Care and Use Committee.
Tissue collection
Mice were fasted for 4 h and then anaesthetized with isoflurane and euthanized via exsanguination. Trunk blood was collected from the area of decapitation. The aorta was excised and cleaned of surrounding tissue in ice cold physiologic saline solution, flash frozen and stored at −80°C for subsequent analyses. Livers were flash frozen and stored at −80°C.
Plasma nitrite
Plasma nitrite concentrations were determined as described previously 7 using ion chromatography (ENO20 Analyzer; Eicom, San Diego, CA, USA).
Carotid artery vasodilatory responses
Measurements of endothelium-dependent dilation (EDD) and endothelium-independent dilation (EID) were performed in carotid arteries as described previously. 5 Briefly, after equilibration in pressure myograph chambers (DMT, Inc., Atlanta, GA), arteries were constricted with phenylephrine (2 µM), and changes in luminal diameter were measured in response to acetylcholine (Ach: 1 × 10−9 − 1 × 10−4 M) and sodium nitroprusside (SNP: 1 × 10−9 − 1 × 10−4 M).
Aortic stiffness
Mechanical stiffness was determined as previously described. 8 A 2-mm segment of thoracic aorta was loaded onto pins in a wire myograph chamber (DMT, Inc.) and incrementally stretched ~10% every 3 min and force recorded. Another segment was sliced (20 µm) and used for measurement of diameter and wall thickness via ImageJ (Rasband, W.S., ImageJ; US National Institutes of Health, Bethesda, MD, USA).
RNA isolation and gene expression
Total RNA was isolated from aortic tissue using TRIzol reagent (Life Technologies, Carlsbad, CA, USA), and collection was enhanced with the addition of glycogen (1 µg/µL of RNA solution); quantitative real-time polymerase chain reaction (PCR) was performed for interleukin-6 (IL6) and reference gene (beta-2-microglobulin) as previously described. 9
Liver triglyceride content
Flash-frozen liver samples were weighed and digested in ethanolic potassium hydroxide, prior to several ethanol solvent purification steps. Triglyceride content was assayed via commercial kit (Triglyceride Colorimetric Assay Kit; Cayman, Ann Arbor, MI, USA).
Glucose concentrations
Blood glucose was measured from tail vein via hand-held glucometer (Freestyle Lite; Abbot Laboratories, Alameda, CA, USA).
Plasma and aortic cytokines
Plasma and aorta lysates were analysed in duplicate on a single plate to determine the concentration of common inflammatory markers (interleukins: 1α, 1β, 6, 12, 13; monocyte chemoattractant protein-1 (MCP-1) and tumor necrosis factor-α (TNF-α)) using a commercially available multiplex assay (MCYTMAG-70 K; EMD Millipore, Billerica, MA, USA). Intra-assay variability (<5%) was within the normal limits reported by the manufacturer.
Statistical analysis
Data are expressed as mean ± standard error of mean (SEM). Differences between groups were examined for statistical significance using one-way analysis of variance (ANOVA) with Tukey’s post hoc analysis using SPSS (V22). Differences were considered significant at p < 0.05.
Results
As expected, Db-C and Db-N mice displayed significantly higher body weights (Db-C, 44.3±1.7; Db-N, 45.8±1.5; CON, 31.3±0.7 g) and food intake (data not shown) compared to CON. Db-C and Db-N also displayed higher fasting plasma glucose levels (Db-C, 396.3±21.5; Db-N, 383.7±19.8; CON, 92.2±5.0 mg/dL; p < 0.05) and liver triacylglycerols (Db-C, 45.1 + 4.5; Db-N, 40.0 + 4.2; CON, 19.5 + 0.9 µmol/g; p < 0.05) compared to CON, with no effect of nitrite. As shown in Figure 1(a), plasma nitrite levels were significantly elevated in Db-N compared to both CON and Db-C (both p < 0.05). Plasma nitrate, a byproduct of nitrite oxidation, was unaffected by nitrite supplementation (data not shown).
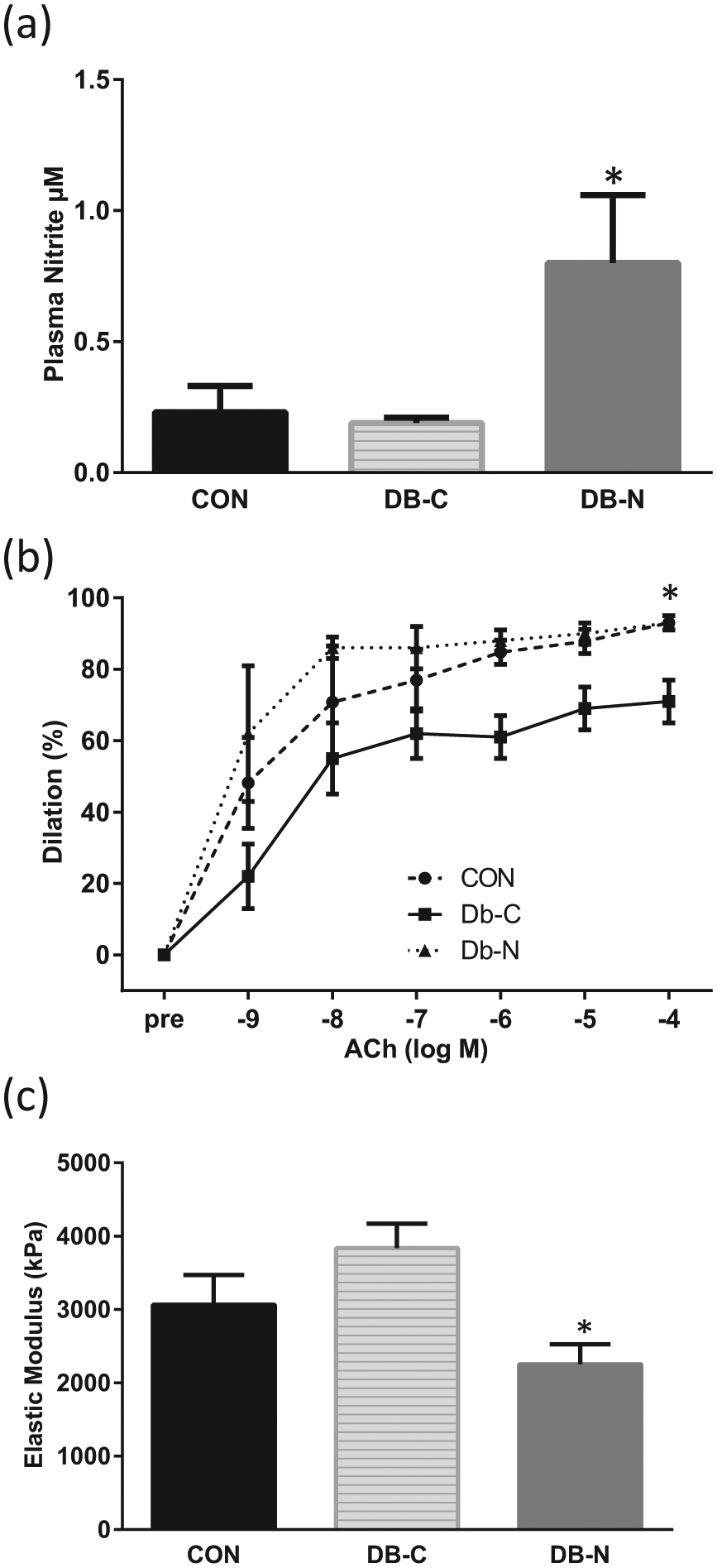
(a) Plasma nitrite levels in heterozygous control mice (CON), Db/Db mice without nitrite supplementation (DB-C) and Db/db mice with nitrite supplementation (DB-N); *p < 0.05 versus CON and Db-N. (b) Dose-responses to the endothelium-dependent dilator acetylcholine (Ach) in CON, DB-C and DB-N; *p < 0.05 versus Db-N and CON. (c) Intrinsic stiffness of aortic segments measured in CON, DB-C and DB-N; *p < 0.05 versus Db-N.
Ach-induced EDD was ~20% lower in Db-C versus CON, and sodium nitrite treatment restored EDD to control levels (Figure 1(b); p < 0.05). In contrast, EID to SNP did not differ among all groups (data not shown). Aortic stiffness was approximately 30% greater (p = 0.20) in Db-C mice compared to CON, and nitrite supplementation significantly reduced this stiffness back to control levels (p < 0.05, Db-C vs Db-N; Figure 1(c)).
To examine potential mechanisms of nitrite-induced improvements in vascular function, we first measured aortic gene expression of IL6, a pro-inflammatory cytokine implicated in the development of endothelial dysfunction. IL6 gene expression was significantly increased in Db-C compared to CON, and nitrite supplementation returned the expression to that of controls (Db-C, 1.4±0.2; Db-N, 0.8 + 0.2 fold changes from CON, p < 0.05). In light of these data, we used remaining aortic tissue to perform a broader analysis of circulating and aortic inflammatory markers (IL1α, IL1β, IL6, IL12, IL13; MCP-1 and TNF-α). Surprisingly, no consistent pattern of inflammation was observed in the plasma or aorta of db/db mice with or without nitrite supplementation (data not shown), suggesting that nitrite’s ability to improve vascular function may involve other factors besides systemic or local inflammation.
Discussion
The key finding of this study is that 5 weeks of oral sodium nitrite treatment improved endothelial function and aortic stiffness in obese, diabetic mice. As expected, db/db mice displayed increased body weight, plasma glucose concentrations and liver triglyceride content compared to heterozygous controls. Five weeks of sodium nitrite supplementation did not significantly affect these metabolic characteristics, suggesting that the beneficial vascular effects of nitrite are independent of alterations in these metabolic parameters.
Five weeks of oral nitrite supplementation significantly increased plasma nitrite concentrations, confirming the effectiveness of nitrite delivery via drinking water. Plasma levels of nitrate, a product of nitrite oxidation, were unchanged, lending support to the concept that plasma levels of nitrite, rather than nitrate, reflect endogenous NO production. 6
Endothelial function was reduced by ~20% in db/db mice versus heterozygous controls, similar to that observed in previous studies, 10 and sodium nitrite therapy restored endothelial function to that of control levels. These results extend previous studies demonstrating that short-term sodium nitrite treatment reverses impaired EDD in mouse models of ageing 5 and hypercholesterolaemia. 6 During the preparation of this manuscript, Jiang et al. reported that 4 weeks of oral nitrite supplementation trended towards improving endothelial function in the aorta of db/db mice. The authors concluded that a longer duration of treatment may be necessary to see functional improvements, which is supported by the 5-week supplementation in the current study. 7
Sodium nitrite therapy also significantly reduced aortic stiffness in db/db mice, similar to a previous report using pulse wave velocity in aged mice.5,8 Therefore, sodium nitrite supplementation has been shown to reduce two different expressions of large elastic artery stiffness in a variety of pathological conditions.
We were unable to identify the exact underlying mechanism(s) by which nitrite improved vascular function. A broad analysis of inflammatory markers in plasma and aorta yielded no consistent increase in diabetic mice, irrespective of nitrite treatment. These findings are in agreement with some 11 but not all 12 previous studies demonstrating inconsistent levels of inflammatory markers in db/db mice compared to controls. Future studies should examine whether other putative mechanisms, such as oxidative stress, mediate the beneficial vascular effects of nitrite in models of T2D. 5
In conclusion, oral sodium nitrite therapy improves vascular function in a mouse model of T2D. Although the mechanisms remain unidentified, our results support further exploration of nitrite as a potential therapeutic option in the treatment and prevention of vascular dysfunction in settings of obesity and T2D.
Footnotes
Acknowledgements
The authors would like to acknowledge Melanie Zigler and Stewart Weber for their technical assistance in acquiring vascular function data.
Author contributions
Amy L Sindler and Kimberly Cox-York contributed equally.
Declaration of conflicting interests
The authors declare that there is no conflict of interest.
Funding
This work was supported by the National Institutes of Health [DK087777 (C.L.G.)], [P30DK048520 (C.L.G.; principal investigator (PI): Hill, J.O.)], [NIH AG013038 (D.R.S.)], and [AG000279 (A.L.S.)]