
Abstract
Visceral obesity is a chronic, low-grade inflammatory disease that predisposes people to the metabolic syndrome, type 2 diabetes and its cardiovascular complications. Adipose tissue is not a passive storehouse for fat, but an endocrine organ synthesizing and releasing a variety of bioactive molecules, some of which are produced by infiltrated immune-inflammatory cells including macrophages. Two different subpopulations of macrophages have been identified in adipose tissue: pro-inflammatory ‘classical’ M1 and anti-inflammatory ‘alternative’ M2 macrophages, and their ratio is suggested to influence the metabolic complications of obesity. These macrophages derive primarily from peripheral blood mononuclear cells (PBMCs). We hypothesised that obesity and the metabolic syndrome modulate PBMC functions. Therefore, alteration of the monocyte response, and more specifically their ability to differentiate toward alternative anti-inflammatory macrophages, was assessed in PBMCs isolated from lean and obese subjects with or without alterations in glucose homeostasis. Our results indicate that PBMCs from obese subjects have an altered expression of M2 markers and that their monocytes are less susceptible to differentiate toward an alternative phenotype. Thus PBMCs in obesity are programmed, which may contribute to the inflammatory dysregulation and increased susceptibility to inflammatory diseases in these patients.
Introduction
Obesity is characterised by an excess of adipose tissue, which predisposes individuals to co-morbidities such as diabetes, atherosclerosis, liver disease and cancer. Obesity is associated with an inflammatory state: plasma concentrations of inflammatory markers including interleukin (IL)-6, tumour necrosis factor (TNF)α, soluble TNFα-receptor and C-reactive protein (CRP) are significantly elevated in obese patients.1,2 Therapies targeting obesity-associated immune and inflammatory defects are thus of potential interest. Most prior research on the cellular mediators of inflammation in obesity has focused on intrahepatic and adipose tissue immune cell function. 3 However, these inflammatory cells derive mainly from circulating precursors, such as peripheral blood mononuclear cells (PBMCs), whose function in obesity-associated immune and inflammatory defects has been less studied. Circulating PBMCs are exposed to the systemic environment, including metabolic factors, such as dyslipidaemia, and inflammatory molecules produced by other organs and tissues, and may thus be programmed and directly involved in the low-grade chronic inflammation related to obesity. PBMCs are composed of lymphocytes and monocytes, the latter being the precursors of macrophages. Macrophages play a key role in inflammatory responses, by infiltrating adipose tissue of obese patients, as well as the vascular wall during development of atherosclerosis.4,5
Macrophages are a heterogeneous cell population, which adapts and responds to a large variety of micro-environmental signals. 6 Th1 cytokines, such as interferon γ (IFNγ), interleukin 1-β (IL-1β) or lipopolysaccharide (LPS), induce a ‘classical’ activation profile (M1). M1 macrophages produce primarily pro-inflammatory cytokines, such as TNFα, IL-6 and IL-12. 7 On the other hand, Th2 cytokines, such as IL-4 and IL-13, induce an ‘alternative’ activation programme (M2) in macrophages. The M2 macrophages dampen inflammatory and adaptive Th1 responses by producing anti-inflammatory factors – IL-10, transforming growth factor β (TGF-β) and IL-1 receptor antagonist (IL-1Ra) – by scavenging debris and by promoting angiogenesis, tissue remodelling and repair.7,8 Moreover, macrophages are plastic cells because they can switch from an activated M1 state back to M2, and vice versa, upon specific signals. 9 Human adipose tissue-infiltrated macrophages (ATMs) are characterised by a mixed M1–M2 phenotype, since ATMs express at their surface specific M2 markers but secrete/release M1 pro-inflammatory factors.10,11 No data are currently available regarding the consequences of obesity and its co-morbidities on the capacity of monocytes to differentiate into alternative macrophages. We thus tested whether obesity and associated alterations in glucose homeostasis affects monocyte differentiation into alternative anti-inflammatory M2 macrophages.
Methods
Research subjects
Subjects were patients operated on at the Department of Endocrine Surgery, Lille University Hospital, France, and enrolled in the ABOS (Atlas Biologique de l’Obésité Sévère) study. Morbidly obese female patients (body mass index (BMI) >35) undergoing bariatric surgery were divided into different groups based on their metabolic characteristics viz. normoglycaemic (n = 10): fasting blood glucose ≤ 6 mmol/l and/or ≤ 7.8 mmol/l at 120 minutes after oral glucose loading; glucose intolerant (IFG/IGT) (n = 10): fasting blood glucose ≥ 6 and ≤ 7 mmol/l and/or ≥ 7.8 and ≤ 11.1 mmol/l at 120 minutes after glucose loading; and diabetic (n = 13): fasting blood glucose ≥ 7 mmol/l fasting and/or ≥ 11.1 mmol/l at 120 minutes after glucose loading. A group of lean, normoglycaemic individuals operated on for benign and non-inflammatory diseases (n = 10, BMI ≤ 27, glycaemia ≤ 6 mmol/l fasting and/or ≤ 7.8 mmol/l at 120 minutes after glucose loading) was used as the control (Table 1). None of the patients had any clinical symptoms of systemic inflammation. This study was approved by the institutional review board and all patients gave written informed consent.
Clinical and biochemical characteristics of subjects.a
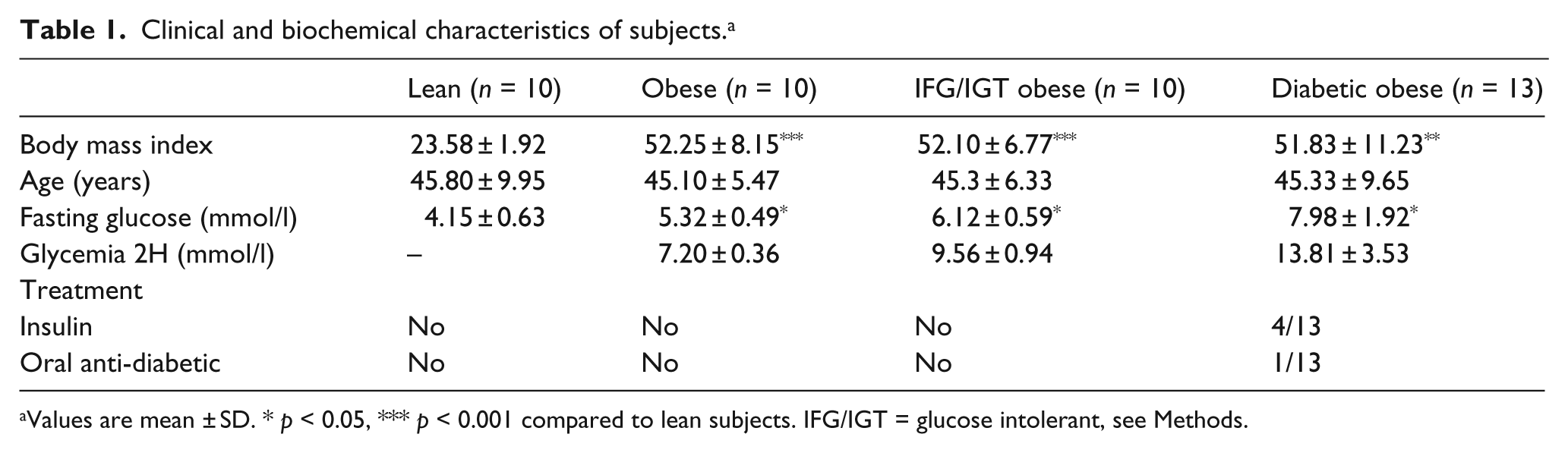
Values are mean ± SD. * p < 0.05, *** p < 0.001 compared to lean subjects. IFG/IGT = glucose intolerant, see Methods.
Cell culture
Peripheral blood was drawn from all patients during the pre-operative clinic visit and from lean control subjects. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll density gradient centrifugation. 12 The isolated monocytes were suspended in RPMI 1640 medium supplemented with 10% human pooled serum (Abcys, France), containing gentamicin (40 µg/ml), glutamine (2 mM) (Sigma) and seeded at density of 1 × 106 cells/well in Primaria 24-well plastic culture dishes (Polylabo, Strasbourg, France) for 2 hours at 37°C. After this period non-adherent lymphocytes were removed by phosphate-buffered saline washes. Differentiation of monocytes into macrophages occurs spontaneously by adhesion of cells to the culture dishes in the presence of RPMI medium containing gentamicin, L-glutamine and 10% pooled human serum (Abcys, France) for 7 days. 13 Monocyte differentiation into alternative macrophages (M2) was assessed by stimulating monocytes with recombinant human IL-4 (15 ng/ml) (Promocell, Germany) added at the beginning of differentiation and for 7 days. 13
RNA extraction and analysis
Total cellular RNA was isolated from PBMCs or differentiated macrophages using Trizol (Invitrogen) and reverse transcribed; the cDNAs were quantified by quantitative polymerase chain reaction (Q-PCR) on a MX4000 apparatus (Stratagene) using specific primers (Table 2). The relative expression of each gene was calculated by the ΔCt method, where ΔCt is the value obtained by subtracting the Ct (threshold cycle) value of cyclophilin mRNA from the Ct value of the target gene. The amount of target relative to the cyclophilin mRNA was expressed as 2–(ΔCt).
Sequences of primers used.
Statistical analysis
Statistically, differences between groups were analysed by Kruskal Wallis and Mann Whitney tests and were considered significant when p <0.05.
Results
PBMCs from obese subjects display a pro-inflammatory phenotype
In order to characterise the phenotype of circulating PBMCs (a cell population including circulating monocytes and lymphocytes), blood was drawn from lean or obese patients with or without alterations in glucose homeostasis, and PBMCs were isolated by gradient centrifugation. Q-PCR analysis of inflammatory genes revealed that PBMCs derived from obese subjects presented a higher gene expression of TNFα, IFNγ and IL-1β, indicative of a pro-inflammatory profile. Interestingly, while the expression of TNFα and IL-1β were most markedly increased in obese diabetic subjects, obesity per se conferred high expression of IFNγ (Figure 1).
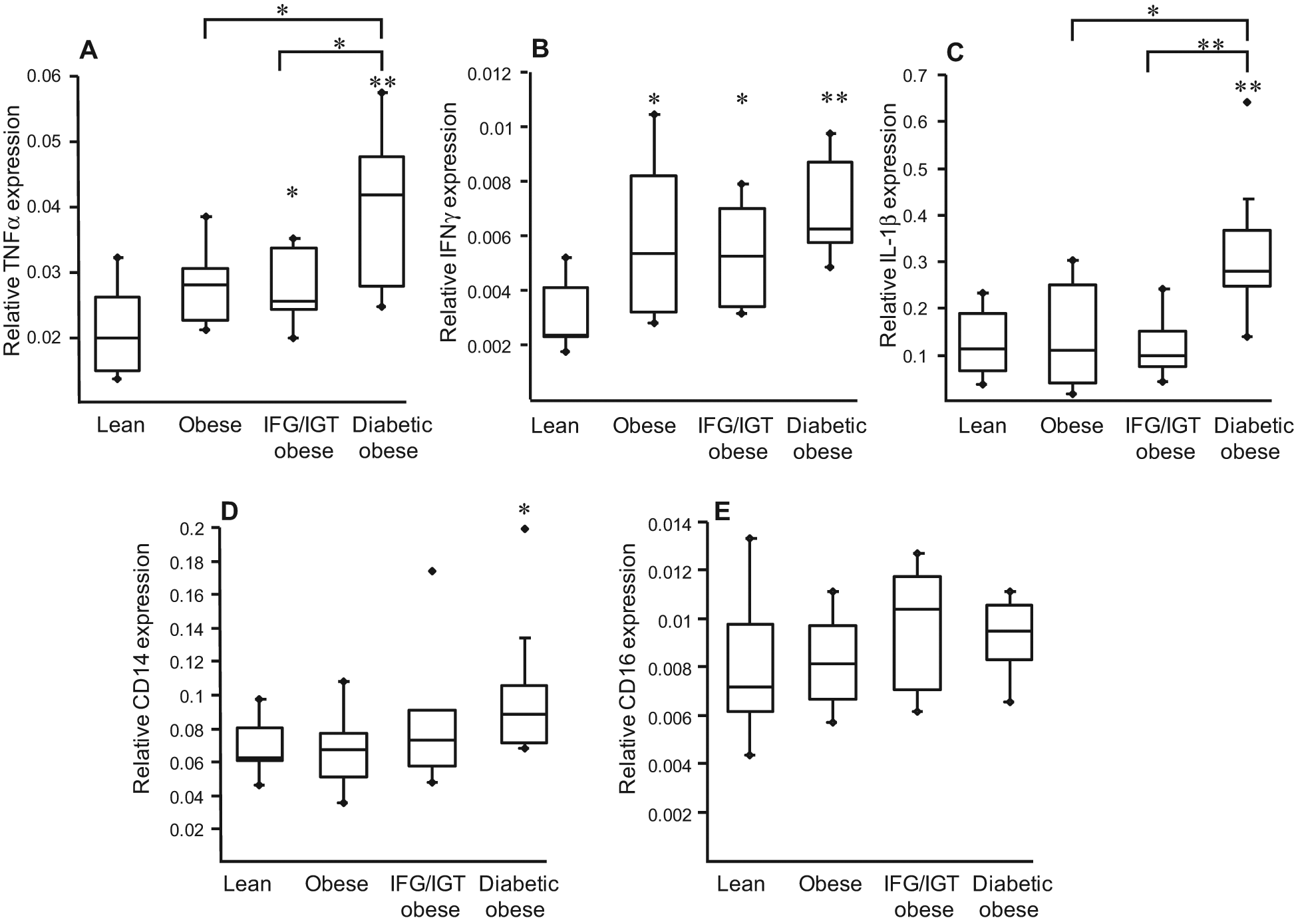
Peripheral blood mononuclear cells (PBMCs) from obese subjects display a pro-inflammatory profile. PBMCs were isolated from lean and obese subjects. The mRNA levels of TNFα, IFNγ, IL-1β, CD14 and CD16 were measured by Q-PCR. The relative expression of each gene was calculated as described above and represented in Boxplots indicating the median and the lower and the upper quartiles. Statistically significant differences are indicated (* p < 0.05, ** p < 0.01). IFG/IGT = glucose intolerant, see Methods.
PBMCs from obese subjects display altered expression of alternative differentiation markers
No major difference was observed in the expression of the lymphocyte markers (CD3, CD4 and CD8) with the exception of CD4 which was slightly decreased in PBMCs from obese subjects (Figure 2).
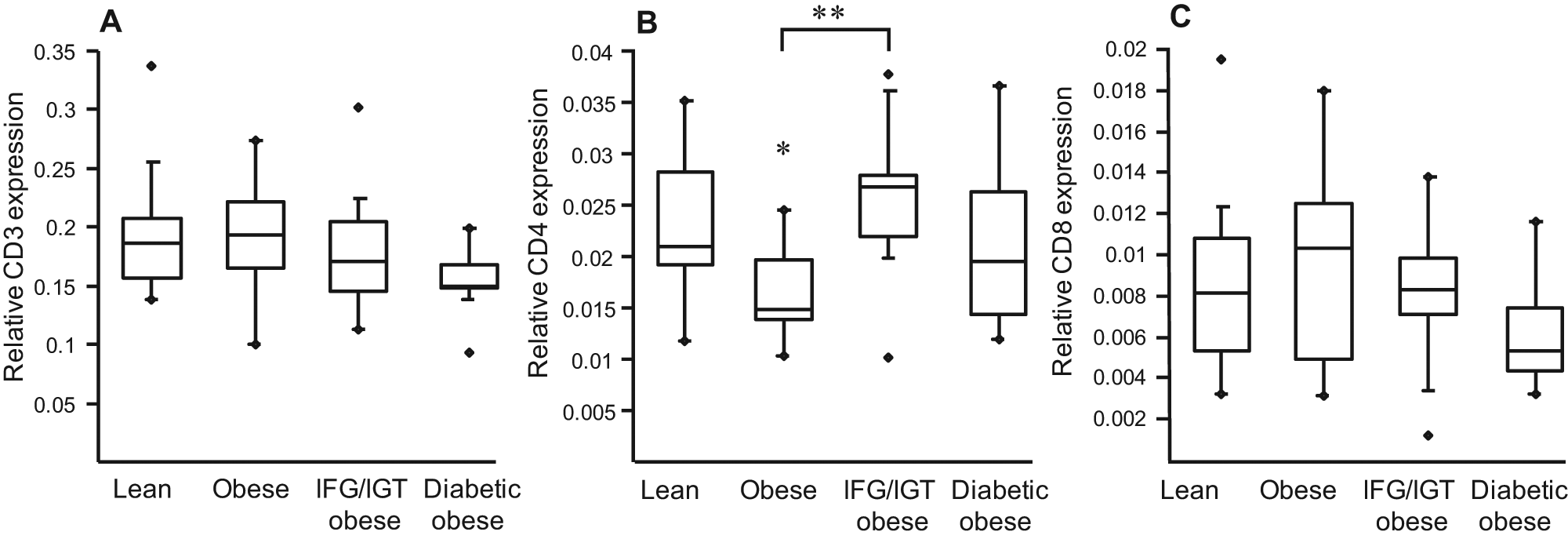
Expression of lymphocyte markers is similar in PBMCs from lean and obese subjects.
To determine whether markers of alternative macrophage differentiation are altered in PBMC from these subject groups, MR, CD163 and CD200R mRNA levels were measured. The expression of other alternative macrophage markers (F13A1, AMAC1) was not detectable in circulating PBMCs (data not shown). Expression of general monocyte markers, such as CD14, was slightly higher in obese diabetics, with no difference for CD16 (Figure 1), whereas MR, CD163 and CD200R were lower in PBMC from obese subjects compared with lean individuals, while no significant differences were observed between obese individuals with different metabolic complications (Figure 3). These results suggest that precursor mononuclear cells from obese subjects already display an altered expression of markers of alternative macrophage differentiation.
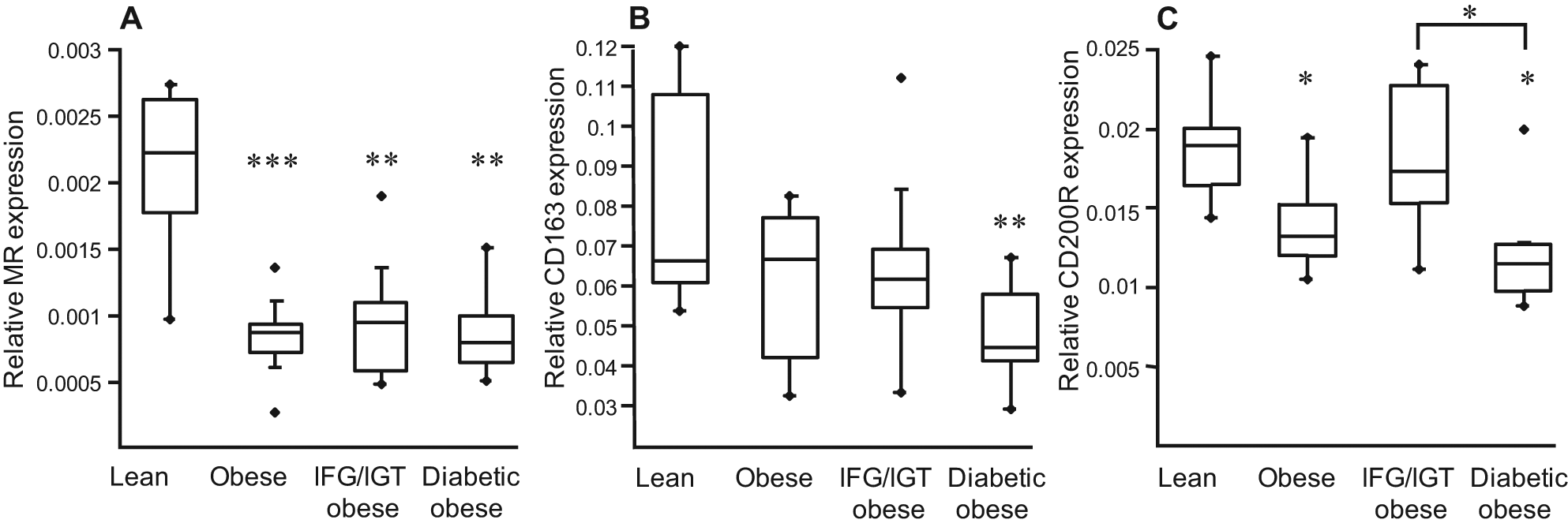
Expression of markers of alternative differentiation is altered in PBMCs from obese subjects. PBMCs were isolated from lean and obese subjects. The mRNA levels of MR, CD163 and CD200R were measured by Q-PCR. The relative expression of each gene was calculated as described above and represented in Boxplots indicating the median and the lower and the upper quartiles. Statistically significant differences are indicated (** p < 0.01, *** p < 0.001). IFG/IGT = glucose intolerant, see Methods.
Monocytes from obese patients display a reduced capacity to differentiate toward an alternative phenotype
To determine the influence of donor phenotype on the ability of monocytes to differentiate toward alternative anti-inflammatory macrophages, monocytes from lean and obese donors (normoglycaemic, glucose-intolerant and diabetic) were differentiated for 7 days in the presence of IL-4, a commonly-used potent inducer of the macrophage alternative phenotype. 13 Our results demonstrated that the gene expression of CD200R, F13A1, TGFβ, AMAC1, CD163 and MR – all established markers for the alternative differentiation – was drastically lower in the alternative macrophages derived from monocytes of obese subjects compared with those isolated from lean individuals (Figure 4). Interestingly, no significant differences were observed between the three groups of obese patients, indicating that obesity alone is sufficient to cause altered monocyte responses (Figure 4).
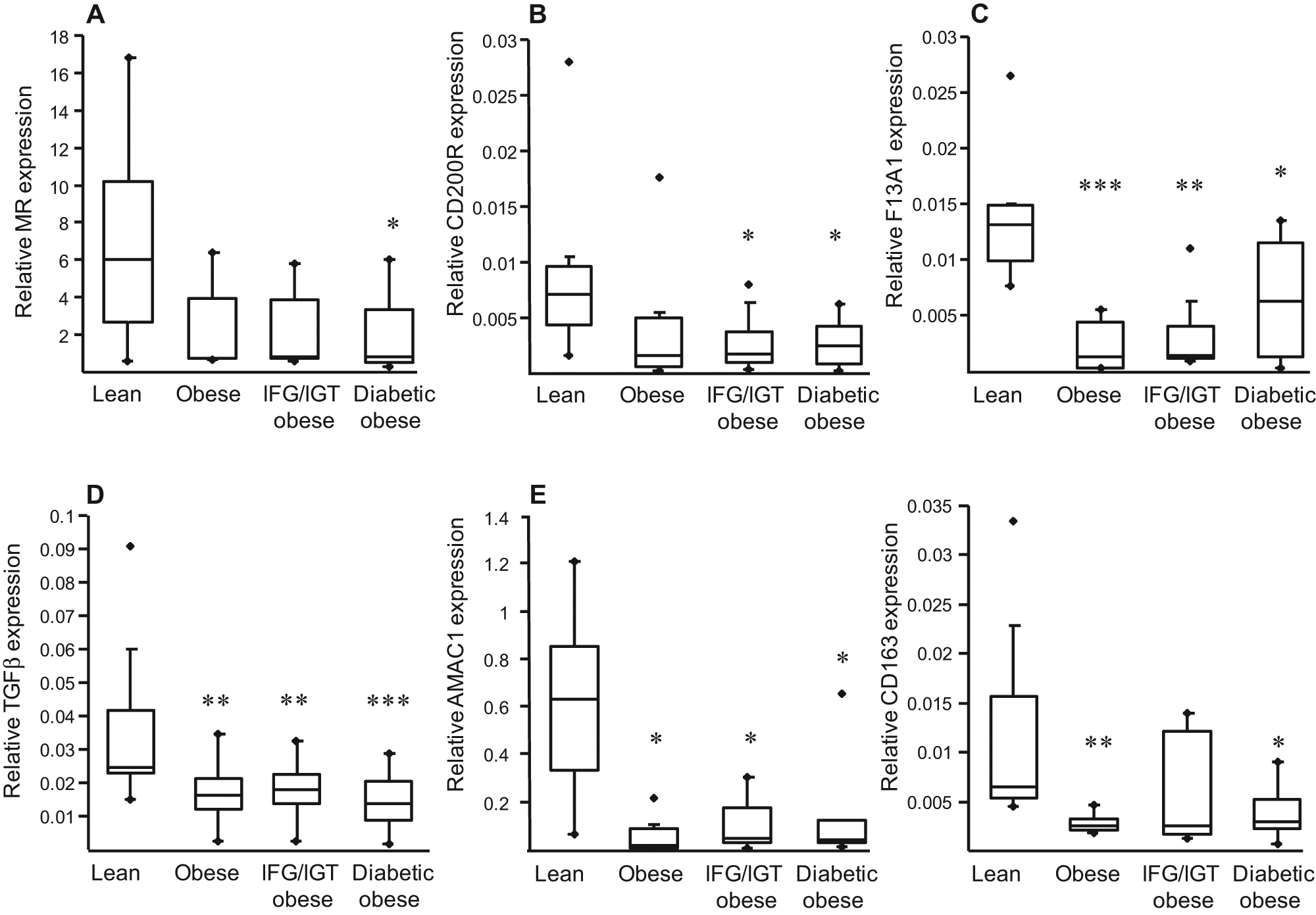
Monocytes from obese subjects display an altered capacity to differentiate into IL4-driven alternative macrophages. Monocytes from lean or obese subjects were differentiated into M2 macrophages in the presence of IL-4 (15 ng/ml) for 7 days. At the end of differentiation, mRNA was extracted and expression of markers of alternative differentiation (MR, CD200R, F13A1, TGFβ, AMAC1 and CD163) analysed by Q-PCR. The relative expression of each gene was calculated as described above and represented in Boxplots indicating the median and the lower and the upper quartiles. Statistically significant differences are indicated (* p < 0.05, ** p < 0.01 and *** p < 0.001). IFG/IGT = glucose intolerant, see Methods.
Discussion
Obesity is a major underlying risk factor for cardiovascular disease due to atherosclerosis and through its association/promotion of other major disease risk factors, namely dyslipidaemia, hypertension and hyperglycaemia. Recent studies have provided substantial evidence that obesity is a chronic low-grade inflammatory disease. Adipose tissue is not a passive storehouse for fat, but an endocrine organ synthesising and releasing a variety of bioactive molecules that may modulate the PBMC phenotype and monocyte functions.
PBMCs are biosensors whose gene expression is influenced by circulating molecules produced by all organs as well as environmental factors, such as diet and exercise. As such, blood cell gene expression patterns might reflect the pathological processes developing in the organism. Available studies indicate that the gene expression profile of circulating PBMCs can be a useful biomarker in the identification and diagnosis of the disease, both in prediction of outcome and in monitoring of the response to treatment in patient follow-up. 14
During the last years, several studies have reported increased inflammatory mediators during obesity. Plasma concentrations of inflammatory markers such as IL-6, TNFα and CRP are significantly elevated in obese subjects.1,2 In addition, the expression of alternative macrophage differentiation markers IL-10 and CD163 was significantly decreased in monocytes from obese diabetic patients relative to obese non-diabetic patients. 15 In our study, we did not confirm a difference in the expression of the alternative markers MR, CD200R and CD163 between the three groups of obese patients with different metabolic disorders. However, a strong reduction in their expression was observed when obese subjects were compared to lean individuals. Our results indicate that obesity itself is sufficient to alter the expression of alternative macrophage differentiation markers in human circulating precursor cells.
The influence of obesity and associated alterations in glucose homeostasis on the ability of monocytes to differentiate toward an alternative macrophage phenotype has not yet been evaluated in humans. Interestingly, the altered gene expression profile observed in PBMCs from obese individuals was conserved in monocyte-derived alternative macrophages differentiated in vitro for 7 days in the presence of IL-4, 7 indicating that these cells are refractory to acquiring an M2 phenotype. We did not observe any difference in the expression of alternative differentiation markers in patients with glucose intolerance or diabetes, indicating that obesity per se is the major cause of the observed phenotype. These results indicate that the precursor cells are already programmed and maintain their phenotype once cultured in vitro, suggesting a potential involvement of epigenetic mechanisms.
The fact that monocytes from obese individuals are impaired to a stimulus promoting alternative macrophages can have more general consequences. Alternative macrophages are resident in adipose tissue as well as in atherosclerotic plaques, where they can exert local effects. 16 In adipose tissue, macrophages produce the anti-inflammatory cytokines IL-10 and IL-1Ra, but can also secrete pro-inflammatory cytokines such as TNF, IL-6 and IL-1β, supporting a role of macrophages in obesity-induced adipocyte dysfunction and metabolic disorders. 10 Thus, obesity may be associated not only with a sustained pro-inflammatory reaction, but also with a failure of anti-inflammatory control mechanisms. Understanding the consequences of obesity-associated pathological conditions on PBMC and monocyte functions can represent a promising novel strategy to elucidate the complexity of obesity-related inflammation.
Footnotes
Acknowledgements
We are grateful to the patients participating in this study. Members of F. Pattou’s surgery team, particularly MF. Six, C. Eberle are acknowledged for their skilled contribution. We are grateful to P. Gelé (Centre d’Investigation Clinique, CHRU, Lille) for blood collection.
Funding
The research leading to these results has received funding from the COST Action BM0602 and the European Community’s 7th Framework Programme (FP7/2007-2013) under grant agreement no. 201608). G. Chinetti-Gbaguidi is a recipient of a Contrat d’Interface from the CHRU de Lille.
Conflict of interest statement
None declared.