
Abstract
The MHC (major histocompatibility complex) is a group of genes that play a crucial role in immune recognition and in tolerance of tissue grafting. The MHC has also been found to influence body odors, body odor preferences, and mate choice in mice and humans. Here we test whether verbal descriptions of human body odors can be linked to the MHC. We asked 45 male students to live as odor neutral as possible for two consecutive days and to wear a T-shirt during the nights. The odors of these T-shirts were then described by five evaluators: two professional perfumers and three laymen. One of the perfumers was able to describe the T-shirt odors in such a way that some of the allelic specificity of the MHC was significantly revealed (after Bonferroni correction for multiple testing). This shows that, although difficult, some people are able to describe MHC-correlated body odor components.
Introduction
Odors are important components of our emotional life, and although the role of odors in human mate choice and sexual behavior is not as well studied as it is, for example, in rodents, it is clear that such a connection exists and that it is a complex one (Stoddart, 1990). A group of genes within the MHC (major histocompatibility complex) has been demonstrated to be linked to body odors and odor preferences (reviews in Penn and Potts, 1999; Yamazaki and Beauchamp, 2005; Ziegler, Kentenich, and Uchanska-Ziegler, 2005). The MHC is also one of the most polymorphic regions of the genome and plays a central role in controlling immunological self and non-self recognition (Apanius, Penn, Slev, Ruff, and Potts, 1997).
Human noses can distinguish between two congenic inbred mouse strains that differ only in their MHC (Gilbert, Yamazaki, Beauchamp and Thomas, 1986), and rodents seem to be able to recognize human MHC-types (Ferstl, Eggert, Westphal, Zavazava, and Müller-Ruchholtz, 1992). Wedekind, Seebeck, Bettens, and Paepke (1995) found that women's preference for male odors correlated negatively with the degree of similarity between their own and the men's MHC type. T-shirt odors were judged as more pleasant when they were worn by men whose MHC genotype was different from that of the judging woman. This finding is analogous to findings in mice (Egid and Brown, 1989; Potts, Manning, and Wakeland, 1991; Penn and Potts, 1998a; Roberts and Gosling, 2003; Yamazaki, et al., 1976; Yamazaki, et al., 1988).
Furthermore, the odors of MHC-dissimilar men more frequently reminded the women of their present or a former mate than did the odors of MHC-similar men. These memory associations suggest that the MHC or linked genes still influence human mate choice. A link between MHC alleles and body odor intensities could not be demonstrated (Wedekind, Seebeck, Bettens, and Paepke, 2006). In a second set of experiments with new combinations of T-shirt wearers and smellers, Wedekind and Füri (1997) found again that dissimilar MHC types smell on average more pleasant than similar MHC types, and that again according to the reported memory associations the MHC or linked genes influence mate choice today. Moreover, when men and women sniffed male and female odors, there was no significant effect of gender in the correlation between pleasantness and MHC similarity. Ober, et al. (1997) then demonstrated in American Hutterites that married couples were less likely to share MHC loci than expected by chance, even after incest taboos were statistically controlled for, but the result could not confirmed in two other human populations (Hedrick and Black, 1997; Ihara, Aoki, Tokunaga, Takahashi, and Juji, 2000).
Montag, et al., (2001) demonstrated that an “electronic nose”, i.e. a chemical sensor system combined with pattern recognition software, is able to detect MHC-dependent odor components in mice and humans. Later T-shirt experiments confirmed a link between MHC dissimilarity and the perception of odor pleasantness in humans (Santos, Schinemann, Gabardo, and Bicalho, 2005; Thornhill et al., 2003), and a recent study on romantic couples at the University of New Mexico suggests that as the degree of MHC sharing increased, women's sexual responsivity to their partners decreased (Garver-Apgar, Gangestad, Thornhill, Miller, and Olp, 2006). Here we test whether humans can describe body odors in such a way that the link to the MHC is revealed.
Materials and Methods
We asked 45 male students (average age: 24.6 years, SD = 2.6), which we had typed for their HLA-A, -B and -DR (methods in Wedekind, et al. 1995, see also Appendix), to live as odor-neutrally as possible for two consecutive days, i.e. to use only perfume-free soap that we had provided, to use only clothes and bedclothes that had been washed with perfume-free detergent, to avoid a list of odor-producing food, to refrain from drinking alcohol or smoking tobacco, and to avoid any activities that could potentially produce disturbing smells (e.g., staying in smelly rooms, sexual activity, etc.). During the two nights, they were asked to wear a T-shirt (100% untreated cotton) and to keep it in an open plastic bag in between (we provided the bags). On the third day, these T-shirts were wrapped one by one in a new plastic bag (first layer) and aluminum foil (second layer) and stored at −78°C for later use.
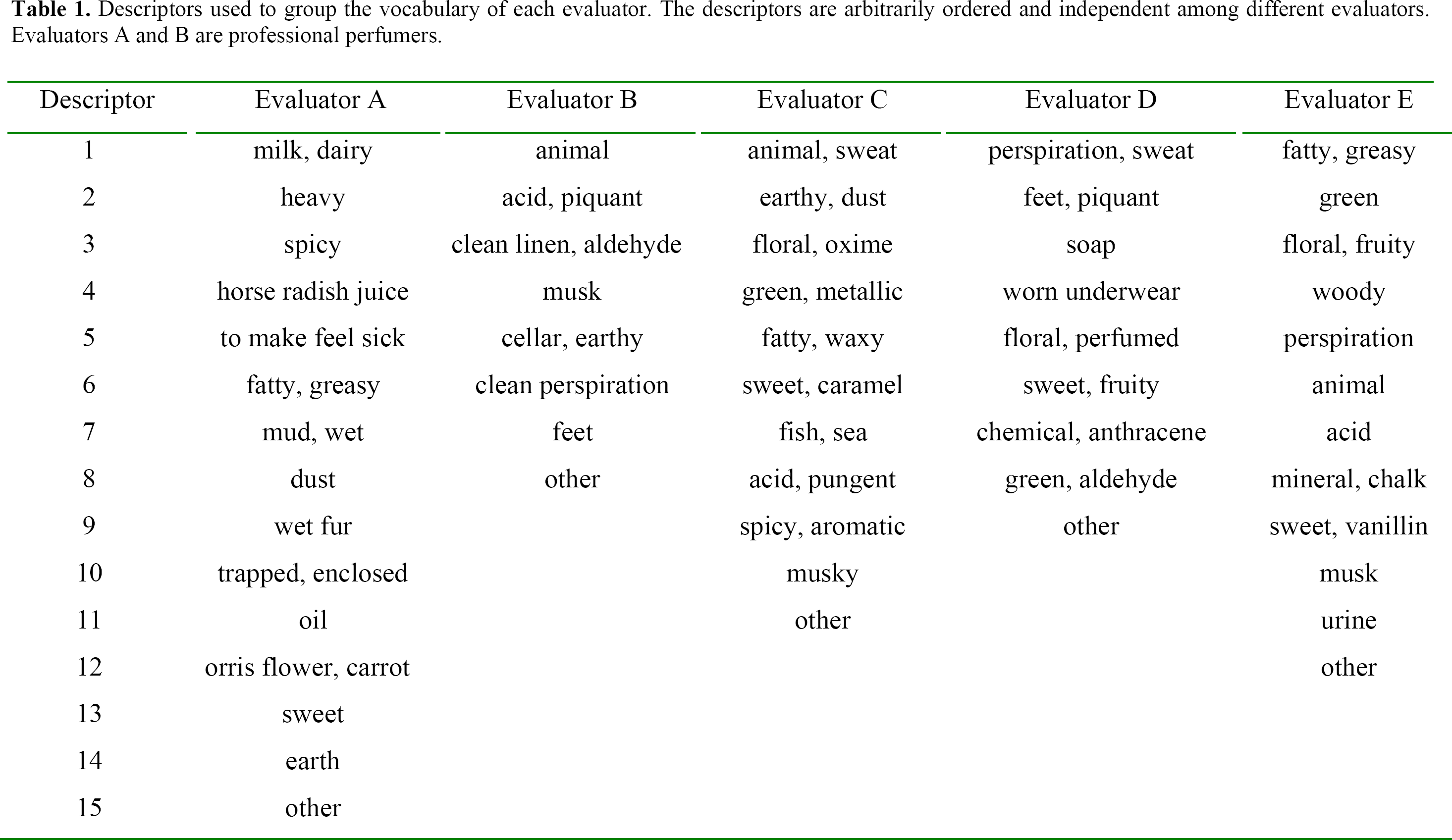
Before the tests, the 45 T-shirts were thawed in 1.5 liter glass containers equipped with a lid. Five evaluators (A, B, C, D and E), two of them professional perfumers (evaluators A and B), then described the odors of the 45 T-shirts by opening these containers and sniffing the content. The evaluators had no information about the MHC-types of the odor donor, and they described the odors independently of each other using their own personal vocabulary. Their evaluations were classified into groups of similar descriptors by a person who was unaware of the MHC-types, with the help of the respective evaluator (the descriptors are listed in Table 1; inconsistencies between the different lists of odor descriptors reveal different kind of odor perception and/or subjective differences in verbalizing odor components). The T-shirts were stored at −30°C in between the days of evaluation.
Descriptors used to group the vocabulary of each evaluator. The descriptors are arbitrarily ordered and independent among different evaluators. Evaluators A and B are professional perfumers.
In order to test whether the odor descriptions of a given evaluator correlated with the MHC of the T-shirt wearers, we used the following statistical procedure: We recorded for every possible pairwise comparison of two T-shirt wearers each (= 990 pairs) whether their odors shared at least one descriptor (i.e. odor similarity for each pair was recorded as 0 or 1), and whether the T-shirt wearers shared more or less MHC antigens than expected from the overall average (i.e. MHC similarity for each pair was also recorded as 0 or 1, the average antigen sharing among all possible pairs was 1.3). To test whether the resulting χ 2 is significantly different from random, we compared it to a distribution of χ 2 that was derived from randomly reassigned data (randomization test with 5000 permutations). The test was done for each evaluator separately. Odor descriptions that were used only once by an evaluator were excluded from this analysis. We calculated directed p-values because if there is a significant link between odor descriptors and MHC-antigens, the χ 2 is expected to be positive (see also Rice and Gaines, 1994, for a discussion of directed p-values). We reduced the critical α-value to 0.01 according to the number of tests we did to account for multiple testing (Bonferroni correction).
Results
The evaluators differed in their average number of odor descriptors per T-shirt (Fig. 1a). Evaluator A (a professional perfumer) provided the most detailed and therefore the most useful description of odors for the further analyses (Fig. 1a). Similarities in odor descriptions given by this evaluator correlated with similarities in the MHC of the T-shirt wearer (Fig. 1b) even after correcting for the fact that we had looked for such a correlation in all five evaluators (Bonferroni correction). No significant overall connection between odor description and MHC could be found in the other four evaluators. We therefore concentrate on the descriptions of evaluator A only for all further analyses.

a) The number of odor descriptors per T-shirt of evaluator A to E (mean±SE; ANOVA: F4 = 32.3, P ≪ 0.001). b) In order to test for a link between odor descriptors and MHC-antigens, we recorded for every possible pair of two T-shirt wearer (990 pairs) both the similarity in odor description as sharing at least one descriptor or none, and the MHC similarity as sharing more or less antigens than the overall average of 1.3. The correlation between these two binary variables is described in a χ 2 for each evaluator. To test whether the observed χ 2 (arrows) are significant different from 0, the null-expectancies (the frequency distributions in the figure) were estimated for every evaluator by repeatedly calculating the χ 2 between randomly reassigned data. “N.s.” means non-significant, p=0.001 depicts a significant deviation from the null-expectancy after correction for multiple testing (Bonferroni correction).
Figure 2 lists the odor descriptors of evaluator A in relation to their tendency to correlate with the MHC, i.e. to the average number of MHC-antigens shared by the T-shirt wearers whose odors were described with one of these descriptors. It appears that the descriptors “spicy” and “mud, wet” correlated best to the MHC of the T-shirt wearers, and the category “other” (that summarizes all descriptions that were used only once and could not be attributed to one of the chosen descriptors) appeared to correlate worst to the MHC (Fig. 2).

Odor descriptors of perfumer A relative to the average number of MHC-antigens shared by the T-shirt wearers whose odors were described with one of these descriptors. The figure shows the mean deviation (± SE) from the average number of MHC-antigens shared by all T-shirt wearers (= 1.3 MHC antigens), the width of the boxes corresponds to the number of odors that were described with the respective descriptor (e.g. “spicy”: n=13, “other”: n=6). Black boxes depict strong deviations from the null expectancy (one-sample tests, “spicy”: t = 3.56, p = 0.0004; “mud, wet”: t = 2.71, p = 0.006, directed).
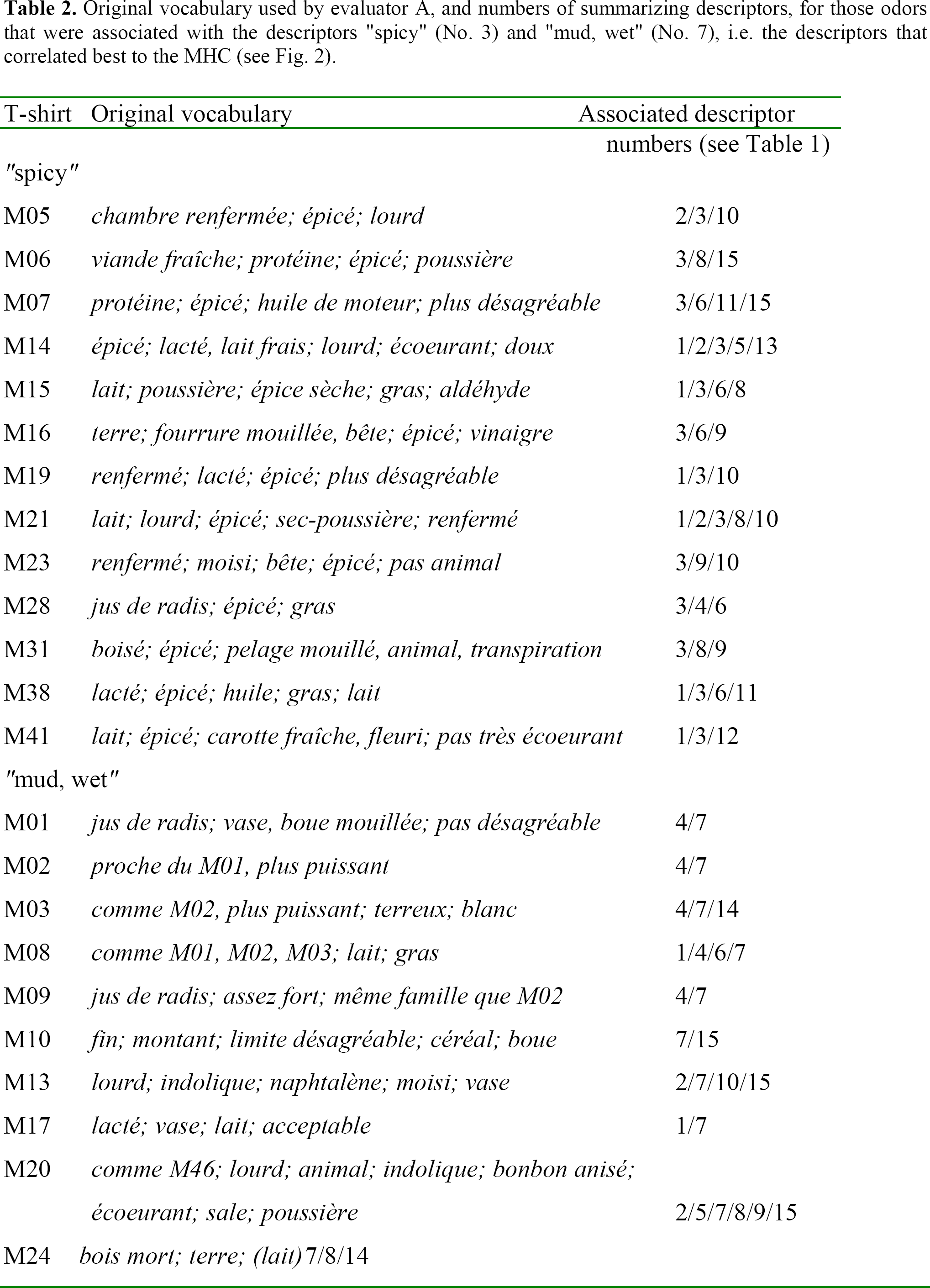
Table 2 lists the original vocabulary (in French) that the evaluator used and that was associated with the descriptors “spicy” and “mud, wet”, respectively. The table reveals that these descriptors assemble quite a heterogeneous group of odors, although the global analysis in Figure 1 detected a general association between these descriptors and the MHC. None of the 13 men whose odor was described as “spicy” possessed the antigen HLA-A1, a common antigen in the study population. However, these men carried the antigen HLA-A2 (another very common antigen) more often than expected by chance (Fig. 3). It further appears that men with HLA-B14, a relatively rare antigen, smelled like “mud, wet” to the evaluator. These and all further comparisons between observed and expected frequencies of the most common HLA-A, -B, and -DR antigens are given for the two descriptors “spicy” and “mud, wet” in Figure 3. In this figure, Z-values are used to weight sample size and actual differences between observations and null-expectancies. These Z-values were calculated using the formula
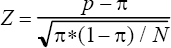
where p is the observed frequency, p the expected frequency under the null-expectancy, and N the number of subjects in a group (Glass and Hopkins, 1984). This more detailed antigen-specific analysis also indicates how much of the variance in the MHC is not, or only weakly, connected to the odor descriptors: most Z-values are small and < |2|.

Exploring possible links between common MHC antigens and the two odor descriptors “spicy” or “mud, wet” of evaluator A. In order to weight the sample size and the actual difference between observation and null-expectancy, the figure gives the Z-values of a comparison between observed and expected frequencies of HLA-A, -B, and -DR antigens in men whose odor was described as “spicy” (above) or “mud, wet” (below). See text for the calculations of Z-values. Only antigens that could be found in at least five T-shirt wearers are plotted (bold: n > 10). Positive Z-values indicate that an antigen may be more often present than expected by chance, negative values indicate that the antigen may be less often present than expected by chance.
Original vocabulary used by evaluator A, and numbers of summarizing descriptors, for those odors that were associated with the descriptors “spicy” (No. 3) and “mud, wet” (No. 7), i.e. the descriptors that correlated best to the MHC (see Fig. 2).
Discussion
The evaluators reported huge difficulties in verbalizing the variation in human body odors. This may explain why the three laymen and even one of two professional perfumers who are used to evaluate and describe fragrances could not find enough odor descriptors that would enable us to do a sensitive statistical analysis. Nevertheless, one evaluator found on average more than three different descriptors per T-shirt. It turned out that these descriptors provided enough information about the odors to link them significantly to the MHC type of the T-shirt wearers (after Bonferroni correction for multiple testing). Although the link between MHC and odor description was only significant in one of five evaluators, our findings show that the association is possible, i.e. odors can, in principle, be described in a way that the description is statistically linked to the MHC. Our more detailed analysis suggests that some descriptors may be more closely linked to certain MHC antigens than others. Such first observations offer testable predictions. However, most of the variation in odor description remains still unexplained.
It seems obvious that the MHC cannot be the only correlate to human body odors (Engen, 1982; Stoddart, 1990; Bestmann, Haberkorn, Vostrowsky, Eggert and Ferstl, 1994; Sommerville, et al., 1994). Even an experimental procedure that controls for disturbing odors of garlic, tobacco, etc. cannot get rid of, for example, potential age effects, of components that reveal developmental stability (Gangestad and Thornhill, 1998; Rikowski and Grammer, 1999; Thornhill and Gangestad, 1999), or of volatile steroid components that may not be linked to the MHC (Singer, Beauchamp, and Yamazaki, 1997). When the influence of many potentially confounding variables is experimentally reduced, up to 23 % of the variance in pleasantness can be explained by the degree of similarity at the loci of the MHC between T-shirt wearer and smeller (Wedekind and Füri 1997). In mice, the estimates are even higher and reach up to 50% (Ziegler, Kentenich, and Uchanska-Ziegler, 2005). However, the pleasantness of one and the same body odor can be perceived very differently by different individuals. In Wedekind and Füri (1997), when 121 students rated the same six odors for intensity, pleasantness, and sexiness, all the odors received nearly all possible scores from very unpleasant to very pleasant and from very weak to very intense. The perception of human body odors therefore seems to vary enormously, i.e. the descriptors used by evaluator A in the present study may therefore not work for other evaluators.
Ehlers et al. (2000) and Younger et al. (2001) found a gene cluster that contains 36 olfactory receptor genes, of which two belong to the vomeronasal family. This cluster is located at the telomeric end of the MHC complex. Thirteen of these genes were tested and found to be polymorphic. Although the physiology of MHC-correlated body odors and odor preferences is not well understood yet (Penn and Potts, 1998b; Ziegler et al., 2005), this polymorphism, and the proximity of such a cluster of olfactory receptor genes to the MHC, suggests that the olfactory receptor genes could somehow be involved in MHC-related odor preferences. If so, the allelic diversity on these loci could partly be responsible for individual differences in odor description.
Milinski and Wedekind (2001) tested in two experiments whether individual preferences for perfume ingredients correlate with a person's MHC-genotype. They found a significant correlation between the MHC and the scorings of scents in both tests. Different people prefer different fragrances, and a statistically significant part of this variation is linked to variation on the MHC and/or MHC-linked genes like (possibly) the olfactory receptor genes described by Ehlers et al. (2000) and Younger et al. (2001). It should be stressed, however, that the effect found in Milinski and Wedekind (2001) was a weak one, i.e it is probably only detectable in large sample sizes. Also, there are obvious further facets of the psychology of fragrance selection besides MHC-correlated odor preferences (Van Toller and Dodd, 1991; Ohloff, 1992).
In conclusion, we found that MHC-linked odor components can be verbally described, but that it appears to be very difficult to describe odors in sufficient detail to link the description to the allelic specificity of the MHC. Moreover, there are good reasons to assume that the perception of body odors differs between different evaluators. Descriptors that work for one evaluator may therefore not work for another. Our results may help to identify MHC-linked odor components that may then be chemically analyzed. A better understanding of the chemistry of MHC-linked body odor components will eventually contribute to an understanding of the physiology of MHC-linked odor production and odor perception.
Footnotes
Acknowledgements:
We thank the participants for their cooperation and interest in this study. Many thanks also to D. Dobbelaere, S. Füri, M. Perlen, B. Streb, K. Viragh for various kind of support, and T. Bakker, M. Frischknecht, L. Keller, F. Näf, C. Margot, M. Milinski, S. Platek, L. Schärer, T. Seebeck, and the anonymous reviewers for discussion and/or constructive comments on the manuscript. This project was supported by the Roche Foundation and the Swiss National Science Foundation.
APPENDIX
The HLA phenotypes of the T-shirt wearer and the simplified descriptors (see Table 1) used by evaluator A.