
Abstract
Introduction:
Aldosterone can induce changes in the expression or activity of Na+/H+ exchanger isoform 1 (NHE–1) in vascular smooth muscle cells. We aimed to clarify whether chronic mineralocorticoid receptor activation exerts an effect on the activity of NHE–1 in the aorta of mineralocorticoid-induced hypertensive rats.
Methods:
Uninephrectomized male Sprague-Dawley rats received subcutaneously 10 mg/week of desoxycorticosterone (DOCA) with or without 20 mg/kg of spironolactone, or vehicle alone (n = 20). After four weeks of treatment, the animals were sacrificed; the aorta was excised for subsequent studies, including histological analysis, RT-PCR, Western blot, measurement of NHE-1 activity and vascular contractility in the presence or absence of the selective NHE-1 inhibitor ethyl-isopropyl amiloride (EIPA).
Results:
Chronic DOCA treatment increased the NHE-1 activity, systolic and diastolic blood pressure, and aortic wall thickness. All these effects were prevented by co-treatment with Spironolactone (p < 0.05). Phenylephrine-induced vascular contractility was significantly reduced in the DOCA group when EIPA was added in the media (p < 0.05). No significant differences in NHE-1 mRNA or protein levels were detected between groups.
Conclusions:
Chronic DOCA administration induced functional and morphological alterations in the rat aorta that are partially explained by enhanced NHE-1 activity and prevented by spironolactone. However, we did not observe changes in the NHE-1 transcript or protein levels, suggesting that the effect may be due to post-transcriptional modifications induced by mineralocorticoid receptor activation.
Keywords
Introduction
Elevated aldosterone concentrations are highly prevalent in secondary hypertensive patients and have been associated with the development of several cardiovascular diseases.1,2 Current evidence supports that aldosterone contributes to the pathogenesis of hypertension through induction of long-term modifications in vascular tissue including structural remodeling, tone modulation and changes in electrolyte composition of the arterial wall.3,4 The deleterious cardiovascular effects of aldosterone are accentuated in the presence of higher NaCl concentrations, independent from blood pressure or angiotensin II levels.3,4 This suggests that sodium transporters might participate in mineralocorticoid-induced vascular dysfunctions.5–7
The Na+/H+ exchanger isoform 1 (NHE-1) is involved in several physiological vascular processes including regulation of the intracellular pH (pHi), intracellular volume, proliferation and contractility of smooth muscle cells.8,9 NHE-1 activity is increased in hypertensive patients and it has been linked to vascular remodeling and increased peripheral vascular resistance. 10 Non-genomic activation of NHE-1 has been described in response to aldosterone in cultured vascular smooth muscle cells (VSMCs) and in vascular tissue.11,12 However, little is known about the effect of long-lasting hyperaldosteronism on the expression and activity of NHE-1 and its possible impact on the vascular tone. In this study we evaluated the activity and expression of this ion exchanger in the aorta of desoxycorticosterone acetate (DOCA)-salt rats in the presence or absence of spironolactone.
Methods
Experimental model
Uninephrectomized male Sprague-Dawley rats (150 g) were separated into three groups: 1) DOCA: the rats were injected twice per week with recommended DOCA doses, 13 5 mg/kg for one month and received water supplemented with 1.0% NaCl and 0.2% KCl; 2) DOCA-Sp: the rats were injected with DOCA and treated with recommended spironolactone doses, 14 20 mg/kg provided in the food; 3) sham: the rats were injected with vehicle only twice per week for one month and received water supplemented with 1.0% NaCl and 0.2% KCl. After four weeks, blood pressure was measured through the tail-cuff method and the animals were euthanized. The aortas were excised for morphological studies and/or further experiments. The ethics committee of Universidad de los Andes according to the National Institutes of Health Guide approved the experimental protocol for the Care and Use of Laboratory Animals.
Determination of gene expression by real-time polymerase chain reaction (PCR)
Real-time quantitative reverse transcription PCR (qRT-PCR) was performed as described previously. 13 The primers used for NHE-1 qRT-PCR were: 5′-GTCTTTGTGCAGGGCATGACCATT-3′ and 5′-GGTTGAGCTTGTCCTTCCAGTGAT-3′; and 18S (housekeeping) 5′-TTAGAGTGTTCAAAGCAGGCCCGA-3′ and 5′-TCTTGGCAAATGCTTTCGCTCTGG-3′. NHE-1 messenger RNA (mRNA) levels were expressed as relative to the amount of 18S signal.
Determination of protein expression by Western blot
Thoracic aortas were homogenized as described previously. 5 The immunoblot was performed according to a standard protocol. 15 Primary antibodies were rabbit anti-NHE-1 (Abcam) or rabbit anti-tubulin (Cell Signalling) and secondary antibodies were Alexa fluorophore-conjugated. Protein signals were detected using the quantitative fluorescent image system ODYSSEY CLX (Li-Cor).
NHE-1 activity in the aorta
Activity was evaluated through measurement of pHi with the membrane-permeable fluorescent dye acetoxymethyl ester of 2’, 7′-bis (2-carboxyethyl)-5,6-carboxyfluorescein (BECECF-AM), as described previously. 14 Briefly, the wavelengths used for excitation were 495 and 440 nm, while emission was 530 nm. The ratio 495/440 nm was used for pHi determinations. The slope of the curve describing the sodium-dependent pHi recovery was considered as NHE-1 activity.
Vascular reactivity
After the animal’s sacrifice, thoracic aortas were quickly extirpated and placed in cold physiological Krebs-Ringer bicarbonate (KRB) buffer (4.2 mM KCl, 1.19 mM KH2PO4, 120 mM NaCl, 25 mM Na2HCO3, 1.2 mM MgSO4, 1.3 mM CaCl2 and 5 mM D-glucose (pH 7.4)). Two 4 mm-thick aortic rings were prepared after removing the peri-aortic fat and adventitial tissue, taking special care to avoid endothelium damage. Standard isometric tension measurements were performed, as previously described. 16
Aortic wall morphometry
Formalin-fixed, paraffin-embedded aorta fragments were sectioned at 6–8 µm intervals and stained with a conventional hematoxylin and eosin method and with Masson’s trichrome stain, as previously described. 17 Slides were digitalized using an Aperio ScanScope scanner at 20× magnification and tissue measurements were performed using the ImageScope package.
Statistical analysis
Data are expressed as mean ± standard deviation (SD); N denotes number of animals studied. Differences between mean values were assessed by Student t-test. For multiple comparisons, one-way analysis of variance (ANOVA) with Bonferroni correction was performed. Statistical significance level was p < 0.05. Analyses were performed with IBM SPSS Statistics 19.
Results
Biometric characteristics
Uninephrectomized male Sprague-Dawley rats were used as models of hypertension with cardiovascular remodeling, characterized by hypertrophy, fibrosis, and endothelial dysfunction. There were no differences in body mass among the groups (Figure 1(a)). As expected, the blood pressure and the relative left ventricular mass (mg of tissue/100 grams of body mass) were significantly higher in DOCA-treated animals than in controls (Figure 1(b)–(d)). A comparable increase in the aortic wall thickness was found (Figure 2(a) and (b)). In addition, a tendency for enlargement of the collagenous adventitia was also apparent (measured by Masson’s trichrome stain) but did not reach statistical significance (Figure 2(a)–(c)). The hypertensive and hypertrophic effects of DOCA were prominently attenuated by co-administration of spironolactone, as shown in the DOCA-Sp animals (Figures 1 and 2).

Effect of DOCA and DOCA-spironolactone on morphological characteristics, blood pressure, and cardiovascular parameters. (a) Body weight. (b) Systolic blood pressure. (c) Diastolic blood pressure. (d) Relative left ventricular mass (mg of ventricle/100 grams of body weight). Bar graph represents mean ± SD. *<0.05 compared to sham. **<0.05 compared to DOCA, n = 15 per group. DOCA: desoxycorticosterone.

(a) Masson’s trichrome-stained aorta from sham, DOCA, and DOCA-Sp animals. Blue, collagenous material; red/pink, smooth muscle; Green label is the aortic wall thickness measurement. (b) Aortic wall thickness. (c) Collagenous adventitia thickness of aorta, measured by Masson’s trichrome-stain. DOCA: desoxycorticosterone; DOCA-Sp: rats injected with DOCA and treated with recommended spironolactone doses.
Effect of DOCA on NHE-1 in aortic tissue
As shown in Figure 3(a) and (b), there were no statistically significant differences among the experimental groups in the mRNA or protein expression for NHE-1 in the aortic tissue. However, in aortic tissue the DOCA group showed an increased NHE-1 activity compared to the controls (p < 0.05). Spironolactone inhibits the NHE-1 activation induced by DOCA to the same levels as controls (Figure 3(c)).
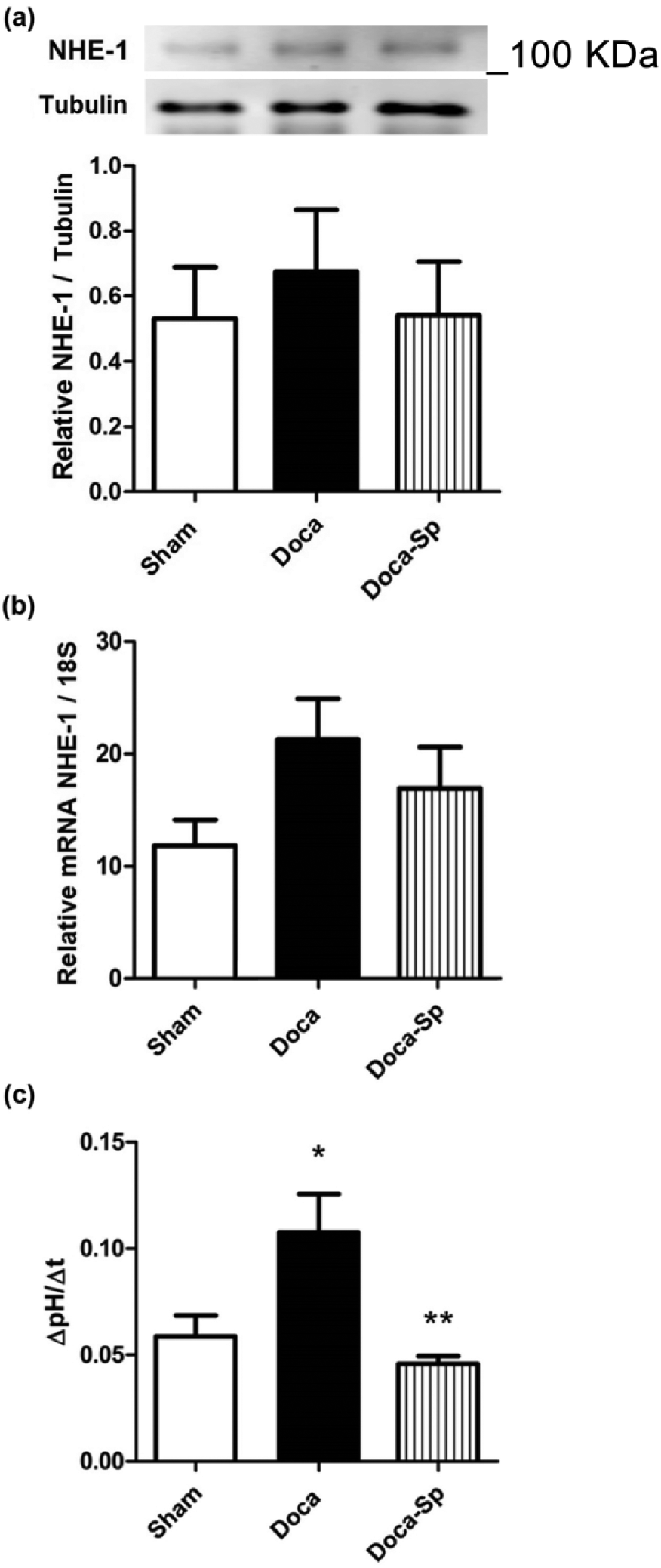
Measurements of NHE-1 abundance and activity in the thoracic aorta of the experimental groups. (a) Relative NHE-1 protein abundance determined by Western blot. A representative picture is shown in the upper section (n = 5 per group). (b) NHE-1 mRNA abundance measured by qRT-PCR (n = 4 for sham, n = 6 for DOCA and DOCA-Sp). (c) NHE activity calculated as change of pHi per time frame (ΔpHi/Δt). Bar graph represents mean ± SD. *<0.05 compared to sham. **<0.05 compared to DOCA, (n = 5 per group). NHE-1: Na+/H+ exchanger isoform 1; mRNA: messenger RNA; qRT-PCR: quantitative reverse transcription polymerase chain reaction; DOCA: desoxycorticosterone; DOCA-Sp: desoxycorticosterone with spironolactone; pHi: intracellular pH.
Vascular contractility
We studied the role of NHE-1 in the thoracic aortas’ contractility in the DOCA-treated animals. The NHE-1 inhibition (ethyl-isopropyl amiloride (EIPA)) did not affect the contractility in the control group (Figure 4(a)). However, the aortic contractility curve was significantly inhibited by EIPA in the DOCA group (Figure 4(b)) starting at 10–7 M of phenylephrine (mean difference 0.231 ± 0.168 g, p < 0.05) up to a peak at 10−5 (mean difference 0.368 ± 0.180 g, p < 0.05). The spironolactone suppressed the EIPA dependence in vascular contractility observed by the DOCA treatment (Figure 4(c)). Consequently, phenylephrine-induced contractile response in hyperaldosteronemic rat aortas was partially dependent on NHE-1 activity (up to 53 ± 44% of the total response).

Aortic contractile response curves to phenylephrine in presence of ethyl-isopropil amiloride (EIPA) or vehicle (dimethyl sulfoxide (DMSO)). (a) Sham. (b) DOCA. (c) DOCA-spironolactone. Markers represent Mean ± SD.*<0.05 EIPA compared to vehicle, n = 5 per group. DOCA: desoxycorticosterone; DOCA-Sp: desoxycorticosterone with spironolactone.
Discussion
Previous studies reported the non-genomic stimulation of NHE-1 in response to aldosterone in cultured VSMCs and in murine vascular tissue.11,12 Furthermore, VSMCs derived from mineralocorticoid receptor (MR) knockout mice also exhibit rapid activation of NHE-1 in response to aldosterone, suggesting that stimulation is independent from the MR-related signaling. 18 Our study indicates that prolonged MR activation enhances the activity of NHE-1 in the aorta of mineralocorticoid-induced hypertensive rats. Additionally, our results show that spironolactone inhibits the NHE-1 hyperactivation, supporting a role for the MR in this response. Hyperactivation of NHE-1 has been described to occur in several ways, including increased transporter expression and/or decreased turnover rate/degradation, changes in pH sensitivity of NHE-1 and/or post-translational modifications such as phosphorylation of either the protein itself or regulatory proteins.19,20 We found no significant differences in the mRNA or protein levels of NHE-1 in aortic homogenized tissue between experimental groups, in contrast with data reported by other authors.21,22 Interestingly, we found a very small tendency toward NHE-1 mRNA and protein increasing in the DOCA-treated group. NHE-1 protein induction was previously described in the myocardium of DOCA-salt rats. 23 To our knowledge, however, no other studies have found changes in NHE-1 protein attributable to hyperaldosteronism in the vasculature. Miyata et al. have suggested an increase in the number of transporters induced by aldosterone in VSMCs, 12 although this assumption was based on the interpretation of NHE-1 activity and not on protein measurements. Based on our results and those reported by others,24–26 we propose that increased NHE-1 activity possibly through post-translational modifications could account for the observations of Miyata et al. 12
NHE-1 is regulated by various other factors at different levels. 20 Intracellular acidosis appears to be the most powerful activating stimulus for this exchanger, which rises dramatically as pHi decreases.27,28 Additionally, the existence of a complex transcriptional regulation though the phosphorylation and interaction of protein kinases with the intracellular C-terminal portion of NHE-1 can modify its activity both in vitro and in vivo. 29 Aldosterone and DOCA can stimulate NHE-1 using mechanisms that would involve inositol trisphosphate (IP3), diacylglycerol, increases of intracellular Ca2+ and activation of protein kinase C (PKC). 30 Moreover, aldosterone could enhance myocardial NHE-1 activity via transactivation of the epidermal growth factor receptor (EGFR) and formation of reactive oxygen species.31,32 Considering that spironolactone inhibits the activity of NHE-1, implying an MR-dependent effect over its function, transactivation of EGFR seems a plausible mechanism through which mineralocorticoids can induce vascular NHE-1 hyperactivity. This mechanism has not yet been proven in vascular tissue and should be attained in further studies.
In our study, NHE-1 made a substantial contribution to vasoconstriction in the aorta of mineralocorticoid-induced hypertensive rats thatwas not detected in the presence of spironolactone. Consistent with our results, vasoconstriction has been previously associated with NHE-1 activation in other vascular territories.16,33 The mechanism through which NHE-1 regulates contractility seems to involve the elevation in intracellular Na+ concentration and the subsequent influx of Ca2+ through a reverse mode of the Na+/Ca2+ exchanger. 32 Interestingly, this pathway has been implicated in the development of cardiac hypertrophy and pathological remodeling, 32 which is observed in hyperaldosteronemic conditions and diminished by the use of MR blockers. 34
We conclude that chronic DOCA administration induced functional and morphological alterations in the rat aorta, an effect mediated partially by NHE-1 activity and prevented by spironolactone. NHE-1 transcript or protein levels were not changed by DOCA, suggesting post-transcriptional modifications induced by MR activation.
Footnotes
Conflict of interest
None declared.
Funding
This work was supported by FAI-MED 004-09 and FAI-MED 001-11 (Research Funds, Universidad de los Andes), and CORFO- N° 14IDL2-30168.