
Abstract
In the adult rodent brain, the continuous production of new neurons by neural stem/progenitor cells (NSPCs) residing in specialized neurogenic niches and their subsequent integration into pre-existing cerebral circuitries supports odour discrimination, spatial learning, and contextual memory capabilities. Aging is recognized as the most potent negative regulator of adult neurogenesis. The neurogenic process markedly declines in the aged brain, due to the reduction of the NSPC pool and the functional impairment of the remaining NSPCs. This decline has been linked to the progressive cognitive deficits of elderly individuals and it may also be involved in the onset/progression of neurological disorders. Since the human lifespan has been dramatically extended, the incidence of age-associated neuropsychiatric conditions in the human population has increased. This has prompted efforts to shed light on the mechanisms underpinning the age-related decline of adult neurogenesis, whose knowledge may foster therapeutic approaches to prevent or delay cognitive alterations in elderly patients. In this review, we summarize recent progress in elucidating the molecular causes of neurogenic aging in the most abundant NSPC niche of the adult mouse brain: the subventricular zone (SVZ). We discuss the age-associated changes occurring both in the intrinsic NSPC molecular networks and in the extrinsic signalling pathways acting in the complex environment of the SVZ niche, and how all these changes may steer young NSPCs towards an aged phenotype.
Introduction
Contrary to what was previously believed, the generation of neurons in the rodent central nervous system (CNS) is a long lasting process, occurring not only during embryonic stages, but also throughout adulthood in restricted areas of the brain.1,2 This has raised great interest in understanding the role of adult neurogenesis in cognition and in exploring whether defects in this process may be involved in the onset/progression of neurological disorders.
In the adult mouse brain, neural stem/progenitor cells (NSPCs) are harboured both in the wall of the lateral ventricles (subventricular zone, SVZ) and in the hippocampus, at the border between the dentate gyrus (DG) and hilus, in a thin layer known as the subgranular zone (SGZ). 3 Although the origin of adult NSPCs is not fully known, two recent papers have employed genetic labelling of embryonic NSPCs in vivo to demonstrate that the majority of the neurogenic stem cells retained in the SVZ, known as B1 cells, are produced from radial glia cells during mid-embryonic development and remain quiescent until they are reactivated in the adult brain.4,5
NSPCs support the generation of new neurons throughout the lifespan of rodents, although tissue organization, niche composition, and the regulatory signals modulating neurogenesis are significantly different between the SVZ and the SGZ, which might affect the type of neurons that they produce. 6 B1 cells generate transient amplifying cells (C cells), which in turn give rise to neuroblasts (A cells) that leave the SVZ and travel through the rostral migratory stream (RMS) to reach the olfactory bulb (OB), where they differentiate into GABAergic and glutamatergic interneurons.7,8 In contrast, NSPCs in the SGZ produce neuroblasts that migrate at a short distance in the overlying granule cell layer, where they differentiate into excitatory glutamatergic granule cells and integrate in pre-existing circuitries. 11
The generation of new neurons in the adult brain is a regulated process, depending on external stimuli, which confers plasticity to the brain.2,9,11,12 Hippocampal neurogenesis is important for learning and memory, anxiety regulation, and social behaviour,13-17 while OB neurogenesis is involved in the structural integrity of the OB, in olfactory memory, in fear conditioning, and in behavioural responses to pheromones.18–20
Increasing evidence suggests the existence of a direct link between neurogenesis impairment and behavioural deficits in several neurological disorders. 21 Dissecting the mechanisms involved in the retention of functional NSPCs is a paramount issue to understand how generation of neurons in the adult brain is supported throughout life under normal and pathological conditions.
Adult Neurogenesis Undergoes an Age-Associated Decline
Neurogenesis in the adult brain is regulated by external stimuli, such as environment enrichment and voluntary exercise, and is sensitive to aging. The negative influence of aging on adult neurogenesis was proposed for the first time at the end of the last century in rodents.14,22–24 In the following years, many studies extended these assessments to other species, such as monkeys, 25 and provided evidence for the cellular and molecular mechanisms underpinning the age-related reduction of neuron generation. By acting on different phases of the neurogenic process, aging impairs the ability of the brain to generate new neurons throughout the entire life. In the hippocampus, aging affects NSPC activity,26–28 reduces the number of progenitors and newly formed neurons, 29 and causes changes in the fate of newly generated cells. Furthermore, Encinas and colleagues 30 showed that NSPC depauperation occurs in the DG of aging mice. They used lineage-tracing approaches to show that NSPCs undergo a limited number of divisions before terminally differentiating into non-neurogenic astrocytes, suggesting that the exhaustion of the stem cell pool acts as a major force driving the age-associated neurogenic decline. 31
In aged rodents, the cytoarchitecture of the SVZ, the area of the adult CNS where the largest pool of NSPCs is retained, is also profoundly altered. The SVZ gets thinner, ependymal and astrocytic cell morphology and functions undergo significant changes, 32 and the RMS tends to disappear, as a consequence of the progressive decline in the number of neurons produced.33–37 These changes correlate with the impairment of olfactory discrimination abilities.35,38
By means of in vivo analyses based on the staining of SVZ tissue sections of different ages with proliferation markers, several studies have firmly established that the pool of actively proliferating NSPCs becomes dramatically reduced during aging, indicating that the curtailed neurogenic output of the aged SVZ is linked to the depletion of the NSPC reservoir.33,35,39-43 The decrease in the amount of NSPCs in vivo is paralleled by a weaker ability of the aged SVZ to form neurospheres upon dissociation and in vitro culture compared with the young adult SVZ.33,38,43 This difference in neurosphere-forming ability was not detectable when the assay was performed with nestin-expressing cells, sorted from young adult and aged nestin:GFP mice, 33 indicating that spared NSPCs from aged mice retain the competence to respond to growth factors in vitro. 33 Other in vitro experiments, however, suggest that NSPC proliferative properties, besides their in vivo numbers, change with the age of the mice from which NSPC cultures originate. For example, a decline in bromodeoxyuridine (BrdU) incorporation was observed in neurosphere cultures of young adult (2 months old) NSPCs compared with newborn animals. Further impairment was found at middle age (15 months old). At 13-15 months, NSPCs also had lower survival rates and generated fewer neurons and more astrocytes, evidencing a generalized impairment of NSPC function at middle age. 38 These data connect the altered behavioural functions of aged mice with the impairment of NSPC functional properties such as survival, proliferation, and lineage fate decision.
To better dissect the changes occurring at the level of the different cell populations residing in the SVZ niche, fluorescent activated cell sorting was used to isolate specific cell subpopulations, including activated B1 cells that were identified through the expression of the cell surface markers carbohydrate Lewis-X and epidermal growth factor receptor (LeX/EGFR).44,45 This work suggests that the pool of LeX/EGFR-positive cells is stable at 6 months and may be retained until 12 months. Nonetheless, an increase in cell cycle length, mainly attributable to a lengthening of the G1 phase, was observed between 2 and 6 months when LeX/EGFR-positive cells were cultured in vitro. The same authors also found that the numbers of C and A cells in vivo, but not their proliferation in vitro, decreased constantly from 2to 12 months, suggesting that a precocious alteration in the proliferative capacity of activated B1 cells leads to the reduction of the progenitor/neuroblast cell populations and accounts for the early neurogenic decline.44–45
In vivo analyses on histological sections of the SVZ confirmed that the number of B1 cells contacting the lateral ventricles was unchanged between 3 and 6 months, but this was followed by a decrease across the entire lateral wall of the ventricle in 1- to 2-year-old mice. This study also found that within the reduced B1 cell population, the percentage of cells that were positive for mitotic markers increased in aged animals, suggesting increased activation of older B1 cells to compensate for the decrease in the NSPC pool. 37 Supporting the idea that age-related NSPC depletion may stimulate the proliferative activity of the residual NSPCs, another study used double labelling with thymidine analogues in vivo to show that dividing NSPCs are fewer in the aged SVZ, but they divide more frequently than young adult NSPCs. 42 Notably, a clonal lineage-tracing analysis of genetically labelled middle aged (12-14 months old) SVZ NSPCs in vivo has been recently reported. Although clone size and spatial distribution were similar in young adult and middle aged mice, mathematical models predict that in comparison with young adult mice, middle aged B1 cells have an increased probability of asymmetric divisions at the expense of symmetric differentiating divisions (ie, giving rise to two C cells), which is counterbalanced by a decreased rate of C cell expansion. According to these models, aging is also associated with an increased duration of B1 cell quiescence, due to reduced activation and inactivation rates. 46
Assessing the functional changes occurring in the mouse SVZ during a longer aging interval (2-22 months), recent work showed that in vivo, the abundance of NSPCs expressing Ascl1, a marker of C cells and a subset of active B1 cells, declined between 2 and 18 months, but it rose up again between 18 and 22 months. This may be due to a more rapid division cycle in the remaining proliferating Ascl1-positive cells, as detected at 22 months both in vitro and in vivo. 47 Despite a partial rescue of the Ascl1-positive population at 22 months, which might be interpreted as an attempt to compensate aging, neuroblast numbers steadily decreased between 2 and 22 months, indicating that lineage progression might be impaired in aged animals. 47 Aging is then a dynamically evolving process that needs to be analysed at different time points, and in specific subpopulations of the neurogenic niche, to be unmasked in its whole complexity.
In the attempt to understand how the NSPC pool is maintained in the adult niche, genetically modified mice have been used to trace NSPC clones in vivo. Tracing the progeny of individual NSPCs through the GlastCreERT2 mouse line (targeting B1 cells) crossed to the ‘Confetti’ multicolor Cre reporter, Calzolari and colleagues 48 showed that SVZ B1 cells undergo rapid rounds of cell divisions within 3 to 5 weeks before becoming consumed. A recent report has shown that B1 cells divide mainly symmetrically, opposite to what was observed in stem cells during embryonic development,49,50 or in the adult hippocampus. 30 Using multiple approaches, including ex vivo time-lapse imaging on organotypic whole-mount cultures, this work found that 70% to 80% of B1 cells went through symmetric differentiative divisions, implying that the majority of the activated B1 cells are exhausted by the generation of C cells; only 20% to 30% were symmetric self-renewing divisions. Such prevalence in consuming divisions leads to a consistent reduction in the number of B1 cells contacting the lateral ventricles, which is also paralleled by a significant age-associated neurogenic decline. 51 The progressive exhaustion of NSPCs in the SVZ, along with the proliferative and differentiative alterations of the residual NSPCs, may then contribute to the age-related reduction of adult neurogenesis, as also proposed by Encinas and colleagues 30 in the hippocampus. Using two different strategies to genetically label quiescent and active B1 cells in vivo and mathematical models, another recent study supports the hypothesis that B1 cells in the young adult SVZ mainly divide symmetrically, but provides evidence indicating that in between bursts of cell divisions, B1 cells can reversibly return to quiescence and maintain long-term self-renewal capacity, 52 similar to previous hypotheses for the behaviour of hippocampal stem cells. 53 Furthermore, this work also suggests cell extrinsic regulatory mechanisms within the SVZ niche, whereby the probability of B1 cell expansion is inversely correlated with the number of surrounding stem cells, 52 which can explain the increased tendency for B1 cell division observed in the stem cell-depleted aged SVZ niche. 37
In conclusion, there is now compelling evidence that the depauperation of the total NSPC population in the aged SVZ is accompanied by changes in the regulation of NSPC proliferation. These proliferative alterations appear to be, however, very complex, with distinct modifications taking place in different cell subtypes at different ages. Importantly, the available data suggest that although some of these functional changes may be intrinsically regulated and captured in vitro, extrinsic niche-based mechanisms play a major role and they may partially explain the discrepancies between in vitro and in vivo and analyses. In the following sections, we will discuss the current knowledge on the intrinsic and extrinsic molecular modifications in the aged SVZ niche that may underlie the above described functional alterations.
Intrinsic Molecular Changes Underlie the Functional Decline of SVZ NSPCs During Aging
Over the last 15 years, outstanding progress has been made in elucidating the intrinsic molecular features that distinguish SVZ NSPCs of different ages and the causal effects of these modifications on the regulation of NSPC proliferation and differentiation. Among them, differences in the regulation of the cell cycle machinery, especially of tumour suppressor pathways, appear to be critical. Recently, genome-wide molecular profiling of young and aged SVZ samples has suggested additional molecular events occurring during NSPC aging, which include changes in the expression levels of specific transcription factors and alterations in the intracellular handling of protein aggregates.
Tumour suppressor pathways are key regulators of SVZ NSPC aging
As described above, recent clonal labelling analyses in the mouse adult SVZ have shown that NSPCs can transit between active proliferation and extended periods of quiescence, thus retaining long-term self-renewal potential. 52 This could make them prone to genetic alterations resulting from errors occurring during DNA replication and/or exposure to various genotoxic agents. In the long run, one possible outcome of accumulating genetic damage in NSPCs could be their oncogenic transformation, as suggested by the hypothesis that severe forms of brain cancer, such as glioblastoma multiforme, may originate from transformed NSPCs. 54 Not surprisingly, tumour suppressor pathways lie at the core of the molecular networks controlling NSPC proliferation and changes in their activities are implicated in the functional alterations of aged NSPCs.
The tumour suppressor gene p16Ink4a encodes for an inhibitor of the cyclin-dependent kinases Cdk4 and Cdk6. These kinases promote cell cycle progression by phosphorylating retinoblastoma family proteins and preventing them from causing G1 cell cycle arrest. 55 The increase of p16Ink4a expression levels during the aging of several rodent tissues provided one of the first observations linking tumour suppressor pathways to aging processes. 55 In particular, in the mouse SVZ, p16Ink4a transcripts are undetectable in young adult (2 months old) mice, rise to detectable levels in middle aged (12 months old) mice and show a further increase in aged (24 months old) mice. 36 Although SVZ neurogenesis is not affected in young adult mice deficient for p16Ink4a, BrdU labelling in vivo and neurosphere assays in vitro indicate that the SVZ of aged mutant mice is enriched in NSPCs, which proliferate more efficiently and generate more OB neurons, in comparison with NSPC depletion and functional impairment in the wild-type aged SVZ. 36 Thus, the age-related increase in p16Ink4a expression levels appears to be functionally implicated in the neurogenic decline of the aged SVZ. Transcription of let7b microRNA in the aging SVZ increases in parallel to that of p16Ink4a, whereas Hmga2, encoding for a chromatin regulator, is downregulated. 56 Functional studies in knockout mice in vivo (BrdU staining) and in vitro (neurosphere cultures) show that opposite to p16Ink4a, Hmga2 is necessary to promote NSPC proliferation in the young adult, but not in the aged SVZ. At the molecular level, it has been proposed that let7b regulates Hmga2 translation by direct binding to its mRNA. Hmga2, in turn, could bind and repress the gene encoding for JunB, a transcriptional regulator known to promote p16Ink4a expression in stem cells 56 (Figure 1A). Notably, homologs of let7b are known to control developmental timing in worms. The mechanisms enhancing let7b transcription in aging mouse neural cells remain unclear.
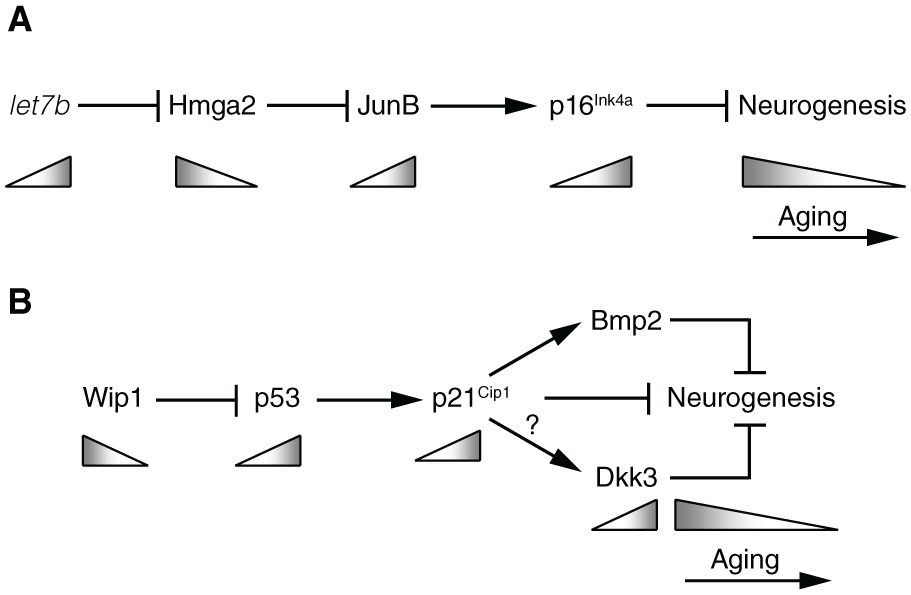
Tumour suppressor pathways implicated in the age-associated neurogenic decline of the mouse SVZ. (A) In SVZ NSPCs, the let7b microRNA represses translation of the Hmga2 protein, which act as a transcriptional repressor for the gene encoding for the JunB transcription factor. In turn, JunB promotes expression of the Cdk inhibitor p16Ink4a. During aging (grey triangles), let7b levels increase, leading to enhanced activity of p16Ink4a and to a decrease in neurogenesis. (B) In SVZ NSPCs, the Wip1 phosphatase acts as an inhibitor of p53, whose transcriptional activity promotes p21Cip1 expression. In turn, p21Cip1 can hinder neurogenesis through several mechanisms: by directly inhibiting NSPC proliferation due to its Cdk inhibitory activity, or by acting as a transcription factor to promote expression of the anti-neurogenic factors Bmp2 and Dkk3. During aging, Wip1 levels decrease, leading to enhanced p21Cip1 activity and a reduction of neurogenesis. The question mark indicates that the regulation of Dkk3 expression by the p53 pathway may occur independently of p21Cip1. Age-related changes in Bmp2 levels in the SVZ have not been described yet. See text for further details.
Another tumour suppressor playing a prominent role in SVZ neurogenesis and in its age-associated decline is p53.57,58 As expected from its anti-proliferative role in many cell types, p53 knockout stimulates the proliferation of SVZ NSPCs and increases their neurogenic output to the OB, whereas expression of a constitutively active p53 deletion mutant represses NSPC proliferation, as shown by staining of proliferating cells in vivo and neurosphere assays in vitro.43,57,58 Similar to p16Ink4a, the levels of active, phosphorylated p53 show an age-associated increase in the SVZ, which is dependent on the concomitant downregulation of Wip1, encoding for a protein phosphatase that can directly inhibit p53 through dephosphorylation. In turn, elevated p53 activity negatively affects NSPC proliferation through upregulation of p21Cip1, a direct transcriptional target gene of p53 encoding for a Cdk inhibitor. 43 In support of this model, adult Wip1 knockout mice show reduced NSPC proliferation (both in the SVZ in vivo and in SVZ cultures in vitro) and OB neurogenesis, which are rescued in Wip1/p53 compound mutants. 59 Moreover, the proliferative and neurogenic deficits of aged SVZ NSPCs in vivo are rescued in transgenic mice overexpressing Wip1. 43 One of the genes regulated downstream of Wip1 and p53 in NSPCs encodes for the Wnt antagonist Dkk3, whose expression levels increase in the aged SVZ. 43 Dkk3 knockout mice show enhanced neuronal production in the SVZ in vivo without a parallel increase in NSPC proliferation, and Dkk3 can antagonize the positive effect of Wnt ligands in the context of an in vitro neuronal differentiation assay. 43 These observations lead to an interesting model in which the Wip1-p53 pathway can perform parallel regulation of NSPC proliferation and differentiation in the adult and in the aging SVZ by acting on different sets of downstream genes. This model is also supported by a separate investigation of p21Cip1 target genes in SVZ NSPCs, which found that, besides its established role as a Cdk inhibitor, p21Cip1 acts as a transcriptional regulator to directly repress Sox2 and promote Bmp2 expression in SVZ NSPCs.60,61 The Sox2 transcription factor has a well-known function in supporting the proliferative state of NSPCs, whereas Bmp signalling promotes their differentiation into non-neurogenic astrocytes. Sox2 and Bmp2 are regulated by p21Cip1 through distinct transcriptional mechanisms and they also appear to act, at least in part, independently of each other, as suggested by results showing that Bmp antagonists can decrease astrocytic differentiation of p21Cip1-deficient NSPCs in vitro without affecting Sox2 levels.60,61 Although the observations resulting from different studies still need to be framed together in a comprehensive model, they suggest that the increased levels of p53, and its downstream effector p21Cip1, in aged NSPCs may curtail their neurogenic potential at different levels, by impairing NSPC proliferative and neuronal differentiation abilities and diverting them towards astrocytic differentiation through the parallel regulation of genes implicated in each of these processes (Figure 1B).
In spite of the negative action of increased p53 and p21Cip1 levels on SVZ neurogenesis, analysis of wild-type and p21Cip1 knockout mice shows that after a transient improvement of NSPC function in 2-month-old p21Cip1 mutants, the p21Cip1-deficient SVZ suffers a stronger age-associated decline in the number of proliferating NSPCs, in their proliferative activity and in neuronal production, based on both staining assays in vivo and neurosphere cultures in vitro. 62 Remarkably, when young adult NSPCs lacking p21Cip1 function are serially passaged in vitro in various experimental settings, they show similar trends of early enhancement of proliferation followed by subsequent deterioration of their ability to expand.60,62 The progressive functional impairment of p21Cip1-deficient NSPCs appears to be due to deregulated Sox2 expression and cell cycle progression, eventually leading to replicative stress, activation of the DNA damage response and inhibition of NSPC proliferation by p53 and p19Arf, 60 another tumour suppressor acting upstream of p53. 55 These results argue against the idea that counteracting the age-associated increase in tumour suppressor levels may be sufficient to prevent NSPC aging and suggest a much more complex scenario, in which a finely tuned increase of tumour suppressor activity in aging NSPCs is required to avoid the total exhaustion of the NSPC pool and preserve at least some of its initial activity in the aged SVZ. Supporting this hypothesis, aged (24 months old) mice that carry a transgene driving a moderate increase in the levels of p16Ink4a, p19Arf and p53 activities show a partial rescue of SVZ NSPC function compared with the wild-type aged SVZ, as indicated both by gene expression analyses in SVZ cells and by neurosphere assays in vitro. 63
Intrinsic changes associated with NSPC aging as revealed by molecular profiling of the young adult and the aged SVZ
Valuable insight into the molecular mechanisms implicated in the neurogenic decline of the aged SVZ has been obtained by means of recent, genome-wide studies of gene activity in SVZ samples of different ages. In particular, five studies have compared the transcriptome of young adult and aged samples by means of RNA sequencing,47,64,65,66 or microarrays analysis. 44 The results of these different investigations are difficult to compare, due to crucial differences in the type and the age of the analysed samples. In particular, two studies analysed specific subpopulations of freshly purified NSPCs, using similar, but not identical, purification protocols.44,64 Other studies employed NSPCs expanded in vitro for two passages 65 or freshly dissected SVZ explants. 47 Importantly, one study performed transcriptomic analyses of whole SVZ dissections, of single cells freshly isolated from dissociated SVZ explants and of in vitro cultured SVZ NSPCs. 66 The age-associated changes found in each of these samples were largely non-overlapping when globally compared, indicating that the biological nature of the SVZ samples used for the analysis significantly affects the final readout. Furthermore, samples of different ages were used for these studies, with young samples ranging between 1 and 4 months old, and aged samples between 6 and 24 months old. Crucially, one of these studies performed comparative analysis of 4 different ages (2, 6, 18, and 22 months old), finding that the majority of the age-related changes were non-monotonic and showed opposite trends before and after 18 months. 47 Therefore, sample age needs to be carefully taken into account when interpreting the results of these studies.
In spite of the critical influence of the chosen methodological approaches, these different investigations suggest some shared and potentially important conclusions. First, on a genome-wide scale, the gene expression profiles of SVZ NSPCs appear to be surprisingly preserved during aging. Although several hundred genes may be differentially expressed between 2 and 18-22 months in the SVZ as a whole, 47 the number of genes showing significant changes between these ages specifically in NSPCs appears to be more limited (roughly 150-250 depending on the study).64-66 This suggests that the functional changes observed in aged NSPCs may involve the altered regulation of specific genes playing key roles in NSPC proliferation and differentiation, rather than large-scale modifications of the NSPC gene expression programmes. This hypothesis is also supported by the analysis of genome-wide DNA methylation and histone methylation in aged SVZ NSPCs expanded in vitro for two passages, showing global similarities in the epigenomic profiles of young adult and aged NSPCs, although changes associated with specific genes were found. 65 Second, gene categories related to cell cycle and neuronal differentiation were represented among the differentially expressed genes in more than one study,44,47,64-66 thus supporting the hypothesis that altered regulation of the molecular pathways controlling NSPC proliferation and differentiation is a crucial event during NSPC aging. Third, other recurrent gene categories in the above studies were those related to the immune process, inflammation, and cell signalling,47,64-66 suggesting that extrinsic cues within the SVZ niche contribute to the molecular and functional changes taking place in aged NSPCs.
The unexpected observation that the expression changes distinguishing young and aged NSPCs appear to be limited to specific genes and molecular pathways leads to the straightforward planning of functional work to test the roles of the differentially expressed genes and pathways during NSPC aging. Some of the studies described above have followed up the results of transcriptomic analyses by means of functional assays. In one study, genes associated with lysosome function were found to be downregulated in aged NSPCs, which correlated with an effective decrease of lysosome levels and activity and an increased accumulation of protein aggregates in comparison with young adult NSPCs in vivo and in vitro. 64 Remarkably, enhancement of lysosomal function by genetic or pharmacological means stimulated the proliferative response of aged NSPCs to growth factors in vitro and increased the number of activated NSPCs in vivo, 64 thus suggesting that the reduced clearing of protein aggregates due to decreased lysosomal activity could be one of the underlying causes, rather than a consequence, of the functional alterations of aged NSPCs. Another study identified the homeobox gene Dbx2, previously known for its role in spinal cord development, 67 as a candidate regulator of neurogenic aging in the SVZ, based on its increased expression levels in cultured or freshly sorted aged (18 months old) SVZ NSPCs in comparison with adult (3-7 months old) samples. 65 As an initial approach to address the function of Dbx2 in this context, the gene was overexpressed in young adult NSPC cultures, which promoted an aged-like phenotype in vitro, including decreased NSPC expansion and transcriptional changes in a cohort of age-associated genes. 65 These initial results lend support to the hypothesis that changes in the expression levels of a few key regulatory genes may be sufficient to drive NSPC aging in the SVZ.
Alterations of Extrinsic Molecular Signals in the SVZ Niche Are Associated With its Age-Related Neurogenic Decline
It is believed that perturbations in the niche environment can also contribute to the progressive impairment of adult neurogenesis observed during aging, although it remains unclear how extracellular signals can affect intrinsic NSPC pathways associated with aging.
NSPCs are harboured in a specialized microenvironment, where they receive and integrate a plethora of converging extracellular factors, whose dynamically regulated production may tune NSPC proliferation and differentiation into neurons, even in the rather anti-neurogenic environment of the adult brain. 68 Remarkably, when transplanted outside of their niches, into intact areas of the brain, NSPCs preferentially differentiate into glial cells.6,69 The extracellular environment of the niche is influenced, by the cerebrospinal fluid (CSF) in the lateral ventricle on the apical side of the niche, by the interaction with the endothelial cells of the blood vessels running on the basal side of the niche, and by non-neurogenic cell types, such as astrocytes and microglia.
Noteworthy, systemic changes in circulating factors within the blood stream can also affect adult neurogenesis.70-73 A study based on plasma injection or heterochronic parabiosis, an experimental procedure that creates a shared circulatory system between two animals of different ages, demonstrated that circulating factors affect the NSPC niche. This study shows that age-related changes in the levels of several circulating molecules can affect brain function and adult neurogenesis 72 and suggests that acting on the circulating milieu might ameliorate the functional deficits associated with aging.70,73
In the following paragraphs, we summarize some of the main age-associated changes taking place in the extracellular environment of the SVZ niche and the functional effects that these modifications may exert on the regulation of the neurogenic process.
Opposite changes in the levels of EGFR and transforming growth factor β signalling are associated with the neurogenic decline of the aged SVZ
Activation of the signal transduction pathway mediated by the EGF receptor (EGFR) is key to promote the proliferation of SVZ NSPCs. 74 Crucially, the number of EGFR-positive cells and the total levels of the EGFR protein as well as those of transforming growth factor α (TGFα), the predominant EGFR ligand in the adult brain, decrease in the aged (24 months old) mouse SVZ when compared with the young adult (2 months old) SVZ. 35 Adult mice with mutations reducing the levels of TGFα show age-associated phenotypes, such as a reduction in the number of SVZ NSPCs, decreased OB neurogenesis and altered olfactory discrimination in comparison with wild-type mice. In contrast, infusions of EGF ligands in the lateral ventricle can increase the proliferating NSPC pool in the aged SVZ. 35 In support of these data, in transcriptomic analyses of single SVZ cells, putative NSPC clusters showed decreased expression levels of genes associated with the extracellular signal-regulated kinase (ERK) signalling pathway, which mediates intracellular signal transduction downstream of activated EGFR and other growth factor receptors. 66 Moreover, the number of NSPCs expressing the active phosphorylated form of ERK1/2 is reduced in the aged SVZ, in comparison with young adult samples, and treatments with ERK inhibitors can decrease NSPC proliferation in vitro. 66 Notably, a subpopulation of actively proliferating NSPCs in the young adult SVZ activates mammalian target of rapamycin (mTOR) signalling, but their number decreases during SVZ aging. Both this decrease and the reduction of the proliferating NSPC pool are counteracted by infusions of EGF in the lateral ventricle of 10-month-old mice, but the effects of EGF are abolished in the presence of mTOR inhibitors. 75 Altogether, these results suggest that the decreased levels of EGFR signalling in NSPCs, due to both receptor loss and reduced ligand availability, lead to altered levels of ERK and mTOR activation, which could play important roles in the neurogenic decline of the aged SVZ.
Highlighting the complexity in the regulation of adult neurogenesis, SVZ NSPCs also express receptors for anti-proliferative signals, such as TGFβ receptors. TGFβ1 ligands are produced by endothelial cells in SVZ microvessels and their levels markedly increase in aged brains (15-24 months old) as compared with young adult samples (3-6 months old). This increase is mirrored by the enhanced phosphorylation of the intracellular TGFβ effector Smad3 in aged NSPCs in vivo, indicating that they activate TGFβ signalling more strongly than young adult NSPCs. 76 Importantly, treatments of in vitro cultures of adult SVZ NSPCs with TGFβ1 slow down their cell cycle and decrease their viability, whereas in vivo delivery of inhibitors of TGFβ signalling can partially rescue the neurogenic activity of the aged SVZ.45,76
All in all, these results suggest that the proliferative activity of SVZ NSPCs is tuned by the balance between the levels of intracellular signalling pathways mediated by EGF and TGFβ receptors. In the aged SVZ, this balance appears to lean towards activation of TGFβ signalling, leading to altered NSPC function and neurogenic decline (Figure 2A and B).
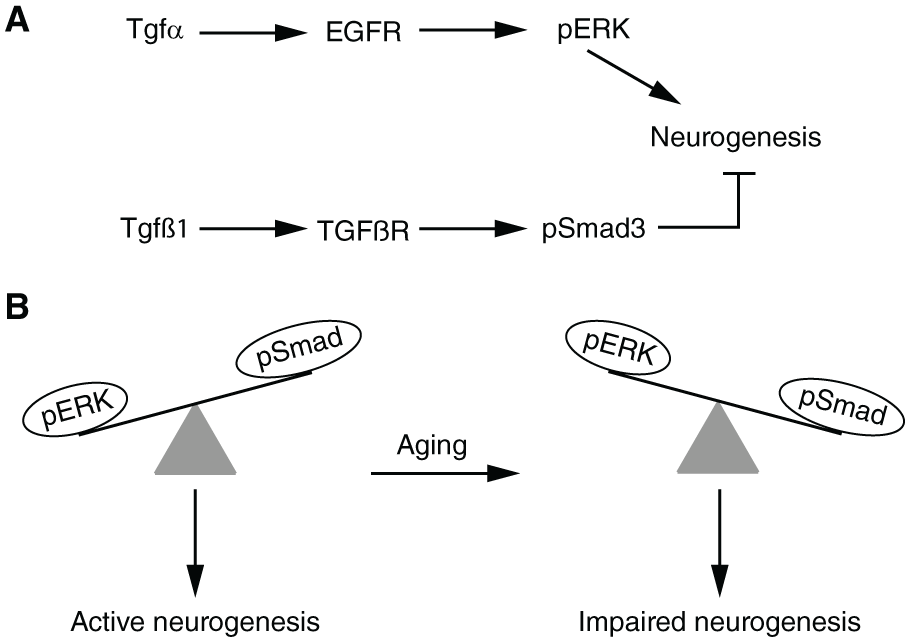
Opposite roles of EGFR-dependent signalling and TGFβ-dependent signalling in the age-associated decline of SVZ neurogenesis. (A) In SVZ NSPCs, TGFα promotes neurogenesis by activating EGFR and increasing the levels of phosphorylated ERK (pERK). In contrast, TGFβ1 hampers neurogenesis by activating TGFβ receptors and enhancing the levels of phosphorylated Smad3 (pSmad3). (B) In NSPCs of the young adult SVZ, ERK activation predominates over Smad3 activation and the neurogenic process is efficiently stimulated. During aging, the relative ratio of ERK and Smad3 activation is reversed, leading to a decline in neurogenesis. See text for further details. EGFR indicates epidermal growth factor receptor; TGF, transforming growth factor.
Age-related modifications in the extracellular signalling mediated by the choroid plexus, microglia and astrocytes affect the neurogenic activity of the aged SVZ
The lateral ventricle choroid plexus (LVCP), an interface between the brain and the circulation, critically contributes to the niche environment for SVZ NSPCs. 77 Conditioned medium from explants of the adult LVCP supported NSPC expansion in vitro, and LVCP conditioned medium increased the number of SVZ NSPCs in vivo when infused in the lateral ventricle of adult mice. Importantly, aged (18 months old) LVCP conditioned medium supported NSPC expansion less efficiently than medium from explants of young adult (2 months old) LVCP, and opposite effects on NSPC numbers were detected following infusions of aged or young adult LVCP medium in vivo. Transcriptome analyses of young adult and aged LVCP showed that gene categories related to immune response, inflammation, and the response to type I interferon (IFN-I) were enriched in the aged samples, opposite to the loss of type II interferon (IFN-II) activity during aging.77,78 Importantly, the number of newborn neurons increased in aged animal in which IFN-I activity was neutralized, whereas reduction of IFN-II signalling is associated with decreased neurogenesis. 78 Furthermore, proteomic analysis of LVCP conditioned medium revealed that the signalling molecules Igf1 and Bmp5, which can both stimulate NSPC expansion in vitro, were produced less efficiently by the LVCP of aged mice when compared with young adult mice and this might explain the reduced ability of the aged LVCP to support NSPC proliferation. 77
Glial cells, such as microglia and astrocytes, are integral cellular elements of the niche, acting as key players in the modulation of adult neurogenesis. 79 Previous work has demonstrated that quiescent microglia controls neurogenesis80,81 and exerts a pro- or an anti-neurogenic effect depending on its activation status.82–84 Aging induces a neuroinflammatory status in the brain, associated with a non-uniform, region-dependent modification of the microglial phenotype. 85 During aging, the relation of microglia with other niche elements in the SVZ is consistently altered; microglial cells become progressively activated and start producing pro-inflammatory mediators, such as TNFα, whose levels rise up with time. 86 For example, reactive microglia are more abundant and produce more TNFα, in the aged SVZ/striatum (22-24 months old) when compared with young adult mice (2-5 months old). 39 These changes appear to be functionally linked to altered regulation of AKT kinase activity in SVZ NSPCs, which is decreased by aged microglia-derived signals in comparison with young adult microglia, as found using in vitro co-cultures of NSPCs and microglia. In turn, AKT can inhibit the glycogen synthase kinase 3β (GSK3β) kinase, which promotes β-catenin degradation. Consequently, exposure of SVZ NSPCs to aged microglia causes an increase in GSK3β activity and a reduction in the levels of β-catenin, leading to the downregulation of transcriptional programmes associated with NSPC proliferation and neuronal differentiation. 39 Remarkably, some of the anti-neurogenic effects of the aged microglia are indirectly mediated through regulation of astrocyte-dependent signalling. Astrocytes in the aged SVZ/striatum express the gene encoding for the Wnt1 ligand less than young adult astrocytes, thus potentially contributing to the reduction in the levels of β-catenin in aged NSPCs. This decrease in Wnt1 transcription may be caused by astrocyte exposure to the aged microglia, as suggested by co-culture experiments of astrocytes and microglia 39 (Figure 3). Aged microglia-dependent signalling is also implicated in the reduced expression levels, in aged SVZ NSPCs, of Nrf2, a transcription factor mediating cellular responses to oxidative stress. Nrf2 promotes NSPC viability, proliferation, and neuronal differentiation, as indicated by gain- or loss-of-function assays in NSPC cultures and by in vivo analysis of Nrf2 knockout mice.38,39

Age-related changes in microglia-derived signals negatively affect neurogenic molecular pathways in SVZ NSPCs. In SVZ NSPCs, PI3 kinase (PI3K) promotes AKT kinase activity, which leads to inhibition of the GSK3β kinase. In turn, GSK3β represses the function of β-catenin, which is critical for the neurogenic process. The activities of PI3K and AKT are inhibited by signalling pathways stimulated by reactive microglial cells, which increase during aging. This causes a decrease in the levels of β-catenin and the consequent decline in neurogenesis. Wnt1 ligands released by astrocytes in or near the SVZ niche collaborate to inhibit GSK3β in NSPCs, but Wnt1 expression is negatively affected by reactive microglia-derived signals and hence decreases during aging, contributing to the reduction of neurogenesis. See text for further details.
Remarkably, the detrimental effects of aging can be partially reversed by physical exercise, 87 which may act, eg, through growth hormone-dependent signalling to activate NSPCs. 88 It is also known that physical exercise decreases the proportion of activated microglia and protects against inflammation-mediated reduction of neurogenesis in aged animals. This suggests that the beneficial effect of running in preventing and or reversing the age-related neurogenic decline relies on the anti-inflammatory mechanisms activated by physical exercise.89,90
Conclusions and Future Perspectives
During the last two decades, the mouse SVZ has established itself as a successful experimental paradigm to investigate the cellular and molecular mechanisms underlying the aging of an adult stem cell niche. It is now clear that the neurogenic activity of the SVZ becomes compromised during aging, and that this neurogenic decline involves intrinsic differences in the aged NSPC population when compared with younger NSPCs. Not only is the NSPC pool severely reduced in the aged niche, but the remaining NSPCs show alterations in their capacity for proliferation and differentiation. These functional changes are linked to distinct molecular modifications occurring within NSPCs, such as the increased activity of cell cycle inhibitors, the decreased ability to deal with intracellular protein aggregation, and the altered regulation of specific transcriptional networks. At the same time, other compartments of the niche, such as the choroid plexus, the vasculature, the microglial and astrocyte cell populations, show age-related modifications too, leading to changes in the extracellular signalling molecules acting within the niche that have a powerful influence on NSPC activity. Exposing young NSPCs to extrinsic cues of the aged niche can accelerate NSPC deterioration, but, excitingly, environmental factors of the young niche can significantly ameliorate the functional deficits of aged NSPCs. These observations raise the attractive hypothesis that aging, although a genetically driven process, does not irreversibly compromise NSPC function, as also suggested by the surprising preservation of aged NSPC transcriptomes and by the equally unexpected finding that many of the observed changes are non-monotonic, ie, they reverse their trend past a critical age. Nonetheless, several challenges lie ahead before it might become possible to translate our knowledge of the mechanisms driving neurogenic aging into therapeutic approaches. First, the molecular changes taking place in the aging SVZ will need to be characterized in much more detail, both at the temporal level, with a special focus on the early onset of aging, and at the cellular level, dissecting the changes occurring in individual cells of each niche cell type. Second, it will be crucial to extend the molecular profiling of aged SVZ cells by including the analysis of the non-coding RNA transcriptome and of the epigenome, given the crucial roles that non-coding RNAs and epigenetic modifications exert on neurogenesis. 91 Third, although in vitro studies will remain essential to dissect the roles of specific molecular pathways and cell types, increasing efforts will be needed to perform functional studies in vivo, using sophisticated genetic and imaging approaches to manipulate and track specific cells in specific time windows. Last but not least, a crucial knowledge gap will need to be filled to shed light onto the physiological role, if any, of the SVZ in adult humans and the possibility of stimulating its neurogenic activity for therapeutic purposes.
Footnotes
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
GL and EC wrote the manuscript. RG PSN SB critically revised the manuscript.