
Abstract
Background:
Antibiotic resistance (ABR) poses a critical global health challenge, necessitating its surveillance across both human and animal health sectors. This study evaluated ABR in bacteria harboured in reared inland fishes sold in Accra and the pond water from which they originated.
Method:
The study was cross-sectional, involving fishes and water sampled from 80 ponds. The gastrointestinal organs of the fishes were homogenised and cultured for bacteria, as were the water samples. The bacteria were identified using matrix-assisted laser desorption/ionisation time of flight mass spectrometry (MALDI-TOF-MS). Antimicrobial susceptibility test was done using the Kirby-Bauer method. Multidrug-resistant (MDR) bacteria were selected for further testing. The double disc diffusion method was used to detect extended-spectrum beta-lactamase (ESBL) production in isolates that were resistant to third-generation cephalosporins. Whole genome sequencing was performed on the ESBL-positive isolates using the Illumina Miseq platform.
Results:
In total, 39 different bacterial species, with their individual numbers totalling 391, were isolated. The bacteria were predominantly Escherichia coli (17%), Aeromonas veronii (11%), Citrobacter freundii (8%), Bacillus cereus (5%), and Klebsiella pneumoniae (5%). The overall ABR rates were cefotaxime (32%), gentamicin (1%), ciprofloxacin (4%), chloramphenicol (19%), tetracycline (37%), meropenem (0%), and ertapenem (0%). Overall MDR and ESBL bacteria prevalence were 13.6% and 1.3%, respectively. The sequence types of the ESBL isolates were ST4684 (80%, n = 4) and ST2005 (20%, n = 1), and the serotypes were H34:09 (80%, n = 4) and H7 (20%, n = 1); the ABR genes were blaCTX-M-15, fosA7, and qnrS1.
Conclusion:
The fishes and the pond water were contaminated with a diverse range of bacteria, mainly Escherichia coli and Aeromonas veronii. The ABR, MDR, and ESBL rates were low to moderate. Moreover, the main sequence type and serotype of the ESBL isolates were ST4684 and H34:09, respectively, and the ABR genes were blaCTX-M-15, fosA7, and qnrS1.
Introduction
Antibiotic resistance (ABR) poses a significant and urgent global health threat and was implicated in the deaths of 4.95 million individuals in 2019. 1 If effective measures are not taken to address antibiotic resistance by 2050, 10 million people could die annually, and the gross domestic product (GDP) could decline by 2% to 3.5%, resulting in a total cost of US$100 trillion. 2 Sub-Saharan Africa, which is disproportionately burdened with antibiotic resistance, is expected to experience a relatively higher impact, with a GDP reduction of US$2895 billion, equivalent to 20% of the region’s total economic output.1,2
Aquaculture is a significant contributor to the problem of antibiotic resistance due to its extensive use of antibiotics.3,4 Concerningly, around 70% to 80% of the antibiotics utilised in aquaculture are released into water systems; fish, being ineffective at metabolising these substances, predominantly excrete them into the environment.3,4 Moreover, antibiotics present in fish feed have the potential to persist in aquatic environments for extended periods, rapidly spreading through water systems and exerting selective pressure on ecosystems, thereby driving the development of antibiotic resistance. 5
To help tackle the antibiotic resistance menace efficiently, it is important to continuously conduct surveillance, targeting interphases that hold high relevance regarding the selection and transmission of antibiotic-resistant organisms among wildlife, humans, and the environment, such as aquaculture ecosystems. However, most antibiotic resistance surveillance studies, particularly, those conducted in sub-Saharan Africa, have mainly focused on humans and rarely on animals and the environment.6 -14 The few that have been conducted on animals have usually been on poultry, cattle, and pigs, with very little attention given to the aquaculture sector.6,7,15 -20 In Ghana, the minimal antibiotic resistance surveillance efforts in the aquaculture sector have mainly been on phenotypic characterisation of antibiotic resistance.15,16,19,20 However, data regarding genotypic characterisation of antibiotic resistance in Ghanaian aquaculture settings are generally sparse. 20 Also, on the whole, there is a paucity of data on the patterns and trends of antibiotic resistance in the aquaculture environment, making data on antibiotic resistance surveillance less comprehensive. To help fill this important knowledge gap, this study evaluated antibiotic resistance in bacteria harboured in reared fishes sold in Accra and the pond water from which they originated, employing a combination of phenotypic and genotypic techniques.
Materials and Methods
Study design and site
This was a cross-sectional study in which farm-reared fishes and pond water samples from which the fishes originated were collected, via random sampling. The farmers raised their fishes in intensive farming system, including earthen pond, concrete tanks, plastic tank, and tarpaulin system at Nii Ashley, Gbentaana, Mammomo, Adjringanor, Manhee, Amanfrom, Maledjor, Frafraha, and New Legon, all of which are situated in the Adenta District of the Greater Accra Region of Ghana (Figure 1). The Adenta Municipal Assembly, which has Adenta as its Central Business District, can be found 10 kilometres to the northeast of Accra. Its precise coordinates are 5°43’ north latitude and 0°09’ west longitude. Approximately 928.4 square kilometres of land make up the municipality. In the east and north, it shares borders with the Ashaiman Municipal Assembly and the Kpong Akatamanso District Assembly. In the west and south, it shares boundaries with the La Nkwantanang Municipal Assembly.
A map of the Adenta Municipality (Source: Ghana Statistical Service, GIS).
Sample collection
The procedures carried out on the fish samples were in accordance with the animal welfare policy of the Veterinary Directorate of the Ministry of Food and Agriculture in Ghana, as well as the sets of animal care and use recommendations. 21 The water and the farmed fishes were both aseptically sampled with the cooperation of the farms’ management teams, and the sampling procedures were carried out in accordance with EPA 22 recommendations. On each day of sampling, the collection of samples was carried out between the hours of 7:00 a.m. and 11:00 a.m. First-hand information on the management techniques followed by the farmers, including the use of antibiotics, was obtained. In total, fishes and pond water from 56 catfish ponds and 24 tilapia ponds were sampled across 12 fish farms. For each pond, five healthy-looking fish were randomly collected from scoop net and casting catches into uniquely-labelled sterile zip-lock bags, following aseptic procedures. Also, 500 ml water samples (in five 100 ml volumes) were collected at five different well-spaced points of the originating ponds using sterile bottles, at a depth of 50 cm below the surface of the water. These samples were transported on ice to the research laboratory of the Department of Medical Microbiology, University of Ghana Medical School.
Laboratory analysis
Bacterial isolation and identification
For each pond, the individual water samples were put together and completely mixed by turning the container slowly and carefully about ten times. A 10 ml portion of each mixed water was placed in a beaker and filled up to the 100 ml mark. The external surface of each fish sample was sterilised with 70% ethanol, and an aseptic incision made across the peritoneal cavity of the fish using a sterile scalpel blade. For the fish from each pond, the intestines and gastric-related organs were carefully removed and separated from the non-gastrointestinal organs, after which the gastrointestinal parts were completely homogenised. This homogenised content (10%) was added to 90% of buffered peptone water and incubated at 37°C for 18 to 24 hours.
After incubation, a loopful of the homogenised mixture was inoculated on blood agar, MacConkey agar, and thiosulfate-citrate-bile salts-sucrose agar (Oxoid, Basingstoke, Hants, UK). For modified semisolid Rappaport Vassiliadis agar, three drops of the mixture were placed on a plate and incubated at 37°C for 18 to 24 hours. From the mixed-growth culture, using the aseptic technique, each group of isolates was obtained using parallel overlapping strokes of the streak method of inoculation. The entire process was repeated for each distinct representative bacterial colony observed in the mixed-growth culture plate, except the modified semisolid Rappaport Vassiliadis agar which was placed on xylose lysine deoxycholate agar and incubated for 18 to 24 hours at 37°C.
Isolates obtained after using the streaking method were identified using Gram staining, and their identities were confirmed using matrix-assisted laser desorption/ionisation time of flight mass spectrometry (MALDI -TOF-MS).
Antimicrobial susceptibility testing
The disc diffusion technique was used to determine the susceptibility of the bacterial isolates using the following antibiotics: ciprofloxacin (5 μg), gentamicin (10 μg), tetracycline (30 μg), meropenem (10 μg), ertapenem (10 μg), chloramphenicol (30 μg), and cefotaxime (30 μg) (Oxoid, Basingstoke, Hants, UK). The criteria of the Clinical Laboratory Standard Institute (CLSI) were followed while conducting the test and interpreting its results. 23
A 0.5 McFarland standard suspension of each bacterial isolate was produced with sterile normal saline and used to inoculate the surface of a sterile Mueller-Hinton agar plate, after which antibiotic discs were carefully placed on the surfaces of the plates. The agar plates were then incubated at 37°C for 16 to 20 hours. After that, diameter measurements of the zones of inhibition were recorded using a pair of callipers, and converted from millimetres to percentages using the CLSI guidelines for specific pathogens and antibiotics. 23
The double disc diffusion method was employed in detecting extended-spectrum beta-lactamase (ESBL) production in isolates that were resistant to third-generation cephalosporins, using cefotaxime (30 μg) and amoxicillin-clavulanate (20/10 μg) or ceftazidime (30 μg) and amoxicillin-clavulanate (20/10 μg) antibiotic discs (Oxoid, Basingstoke, Hants, UK).
Whole-genome sequence analysis
ESBL-producing isolates were sequenced at the Noguchi Memorial Institute for Medical Research, University of Ghana. DNA of each freshly cultured isolate was extracted using the Qiagen DNA MiniAmp kit, following the manufacturer’s instructions. Extracted DNA was quantified using a Qubit 4.0 Fluorometer assay kit (Thermo Fisher Scientific, MA), followed by library preparation with Illumina DNA prep, per the manufacturer’s instructions. DNA concentration between 100 and 500 ng in 30 µl was used as starting material for library preparation. The Beaded-Linked Transposomes (BLT), which included fragmenting and tagging the DNA with adapter sequences, were used to tag the DNA. Following this procedure, a post-tagmentation clean-up was carried out to halt tagmentation and wash the adapter-tagged DNA prior to PCR amplification. The tagmented DNA was subsequently amplified by PCR, and paired index adapters and sequences needed for cluster formation on the flow-cell during sequencing were also added. The amplified libraries were then cleaned up using a post-PCR clean-up procedure to get rid of extra unbound index adapters. The quality and concentration of fragmented DNA were assessed with the 2100 bioanalyser system (Agilent) and qPCR (Kapa Sybr Fast qPCR kit), respectively. Using the fragment sizes generated by the bioanalyser, and the concentration of individual libraries from the qPCR, libraries were then normalised, pooled, and loaded on Illumina 2 × 300 cycle cartridges for sequencing on the Miseq platform (Illumina Inc., San Diego, CA).
Raw sequenced reads were quality-controlled, and adapters were trimmed using FASTQC (http://www.bioinformaticsbabraham.ac.uk/projects/fastqc/) and Trimmomatic (http://www.usadellab.org/cms/index.php?page=trimmomatic), respectively, with a minimum quality set at Q20.24,25 Trimmed reads were de novo-assembled with SPAdes genome assembler optimised with Unicycler V0.4.9 from which only contigs greater than 200 bp were used for further analyses.
Assembled files were uploaded to ResFinder (https://cge.food.dtu.dk/servicesResFinder/), a tool available on the Centre for Genomic Epidemiology (CGE) (http://www.genomicepidemiology.org/) platform, to detect resistance genes present in the sequenced isolates using an identity threshold of 90% and a minimum length of 60%. Prediction of bacterial species and acquired virulence genes were done using KmerFinder (https://cge.food.dtu.dk/services/KmerFinder/) and VirulenceFinder (https://cge.food.dtu.dk/servicesVirulenceFinder/), respectively. Plasmids and mobile genetic elements were predicted using Plasmidfinder (https://cge.food.dtu.dk/services/PlasmidFinder/) and MGEfinder (https://cge.food.dtu.dk/services/MobileElementFinder/), respectively. The sequence types of the isolates were predicted using MLSTFinder (https://cge.food.dtu.dk/services/MLST/). Distinctively, predicting serotypes of the E. coli isolates was done using the SeroTypeFinder (https://cge.food.dtu.dk/services/SerotypeFinder/) at default settings on the CGE platform.
Ethical approval
Ethical approval was obtained from the Ethical and Protocol Review Committee of the University of Ghana’s College of Health Sciences (ID: CHS-Et/M.2-P 4.6/2021-2022).
Results
Characteristics of the sampled ponds
Table 1 summarises the pond characteristics. The ponds on the 12 farms were mainly used in rearing catfish (70.0%, n = 56). Also, the most used holding facility type was tarpaulin (47.5%, n = 38).
Pond characteristics.

Five fish were randomly sampled from each pond.
Distribution of the isolated bacteria in the fishes and pond water
In total, 391 bacteria (of 39 different species) were isolated from the fishes and pond water. In the fish, a total of 201 bacteria, of 38 different species, were isolated. Conversely, the bacterial isolates from the pond water were 190 in total, from 34 species. As a composite, the top five bacteria were Escherichia coli (17%), Aeromonas veronii (11%), Citrobacter freundii (8%), Bacillus cereus (5%), and Klebsiella pneumoniae (5%). In the fishes, the top five bacteria were Escherichia coli (15.4%), Aeromonas veronii (12.4%), Citrobacter freundii (9.5%), Edwardsiella tarda (5.0%), and Klebsiella pneumoniae (4.5%). In the pond water, the top five bacteria were Escherichia coli (19.5%), Aeromonas veronii (8.9%), Bacillus cereus (5.8%), Citrobacter freundii (5.8%), and Proteus mirabilis (5.3%). Table 2 presents the details of the distribution of the bacteria in the fishes and pond water.
Distribution of the bacteria harboured in the fishes and pond water.

Antibiotic resistance of the bacterial isolates
The resistance of all the isolated bacteria to the tested antibiotics, stratified by sample type, are displayed in Figure 2, and in Tables 3 to 5, the distributions of the antibiotic resistance rates across the individual bacteria are presented. Across each of the fishes, pond water, and fishes and pond water combined, the antibiotic resistance rates were highest for tetracycline (31% to 42%), cefotaxime (29% to 35%), and chloramphenicol (19%). No resistance was recorded against either meropenem or ertapenem.
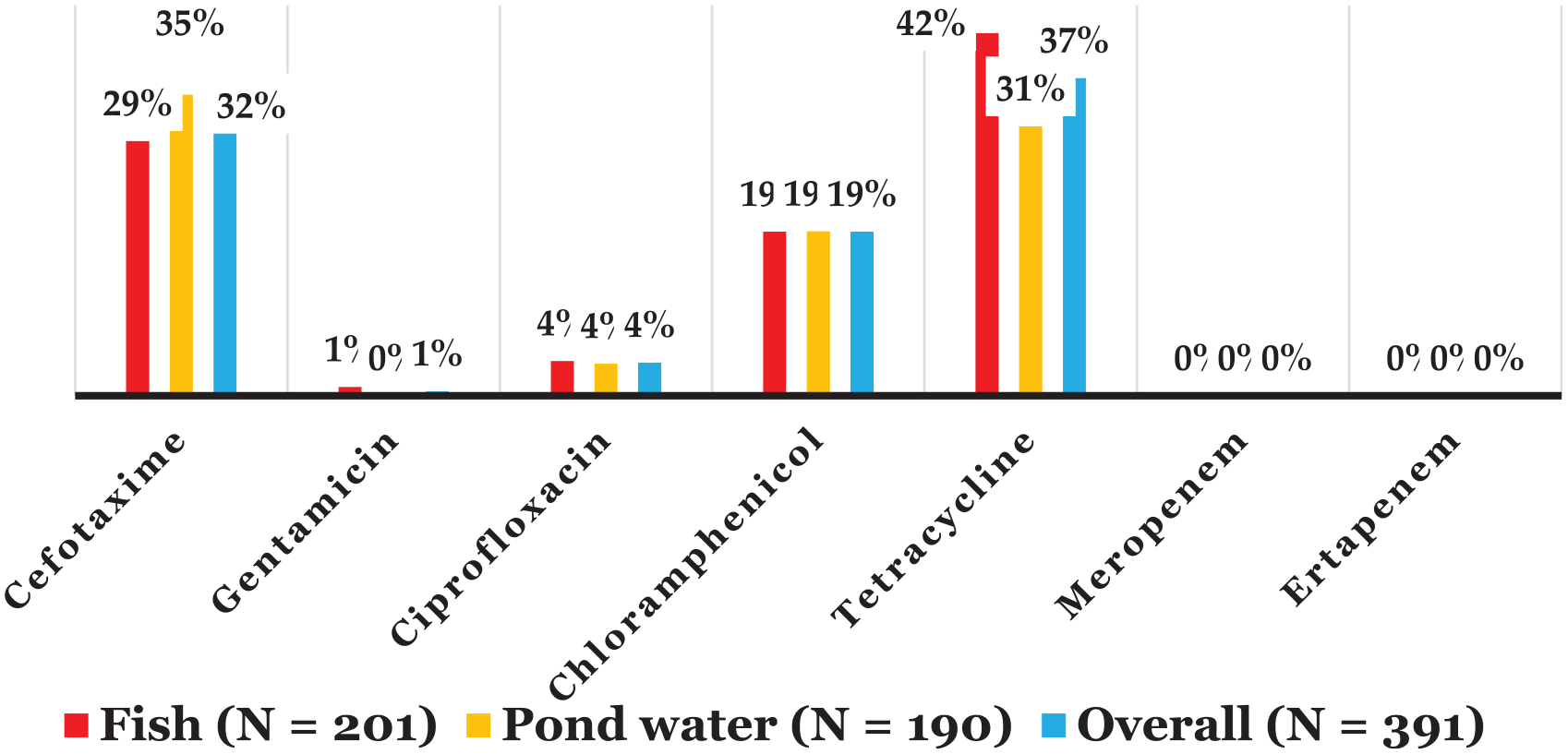
Antibiotic resistance patterns of the bacterial isolates.
Antibiotic resistance among the bacteria isolated from both the fishes and pond water.

CTX, Cefotaxime; GM, Gentamicin; CIP = , Ciprofloxacin; C, Chloramphenicol; TE, Tetracycline; MEM, Meropenem; ERT, Ertapenem.
Antibiotic resistance among the bacteria isolated from the fishes.

CTX, Cefotaxime; GM = Gentamicin; CIP, Ciprofloxacin; C, Chloramphenicol; TE, Tetracycline; MEM, Meropenem; ERT, Ertapenem.
Antibiotic resistance among the bacteria isolated from the pond water.

CTX, Cefotaxime; GM, Gentamicin; CIP, Ciprofloxacin; C, Chloramphenicol; TE, Tetracycline; MEM, Meropenem; ERT, Ertapenem.
Regarding E. coli, the tetracycline resistance rate was 50% in the fishes and pond water combined, 54.8% in the fishes, and 45.9% in the pond water. Chloramphenicol resistance was 37.8% in the fishes and pond water combined, 35.5% in the fishes, and 37.8% in the pond water. Cefotaxime resistance was 38.2% in the fishes and pond water combined, 41.9% in the fishes, and 35.1% in the pond water.
In Edwardsiella tarda, the tetracycline resistance was as high as 80% in the fishes and pond water combined, while chloramphenicol resistance was noted at 26.7%. In the fishes exclusively, the resistance to tetracycline was 90%, accompanied by a chloramphenicol resistance rate of 30%. In the pond water only, the resistance rates were 60% for tetracycline, 20% for chloramphenicol, and an additional 20% for cefotaxime.
For Klebsiella pneumoniae, in the combined fishes and pond water, resistance rates were recorded at 66.7% for cefotaxime and 38.9% for tetracycline. In the fishes alone, rates stood at 66.7% for cefotaxime and 33.3% for tetracycline. In the pond water only, the resistance rates were 66.7% for cefotaxime and 44.4% for tetracycline.
Resistance rates in Klebsiella variicola were notable at 50% for chloramphenicol and 100% for tetracycline in combined fishes and pond water. Similarly, in the fishes alone, resistance persisted with rates of 50% for chloramphenicol and a maximum resistance of 100% for tetracycline.
Citrobacter freundii, in the combined fishes and pond water, showed resistance of 30% for cefotaxime, 43.3% for chloramphenicol, and 40% for tetracycline. In the fishes alone, the resistance rates were 31.6% for cefotaxime, 42.1% for chloramphenicol, and 52.6% for tetracycline. In the pond water only, the resistance was 27.3% for cefotaxime, 45.5% for chloramphenicol, and 18.2% for tetracycline.
Citrobacter braakii demonstrated tetracycline resistance of 33.3% in the combined fishes and pond water and a higher resistance of 66.7% in the fishes alone. In the case of Kurthia gibsonii, in the combined fishes and pond water, it showed 25% resistance to ciprofloxacin, while in the fishes alone, resistance rates were 37.5% for ciprofloxacin, 25% for gentamicin, and 50% for tetracycline.
For Aeromonas veronii, cefotaxime resistance was 21.4% in the combined fishes and pond water, with a slightly elevated rate of 24% in the fishes only. In the pond water only, the resistance rates were 17.6% for cefotaxime and 23.5% for ciprofloxacin. Aeromonas jandaei, in the combined fishes and pond water, exhibited a tetracycline resistance of 30.8%, whereas in the fishes only and pond water only, the resistance rates were 28.6% and 33.3%, respectively. Similarly, Aeromonas caviae had various resistance in the combined fishes and pond water, with rates of 20% for cefotaxime and 50% for tetracycline, while in the fishes only, there was tetracycline resistance of 50%. In the pond water only, cefotaxime, chloramphenicol, and tetracycline resistance were observed to be 50%, 25%, and 50%, respectively.
Regarding Enterobacter bugandensis, in the combined fishes and pond water, the resistance rates were 22.2% for cefotaxime, 44.4% for chloramphenicol, and 55.6% for tetracycline. In the fishes only, there was a high resistance to chloramphenicol (66.7%) and tetracycline (66.7%). Also, in the pond water alone, the resistance rate was 33.3% for cefotaxime, 33.3% for chloramphenicol, and 50% for tetracycline.
Enterobacter kobei displayed significant tetracycline resistance, with rates of 83.3% in the combined fishes and pond water, 33.3% in the fishes only, and 66.7% in the pond water. Likewise, Enterobacter cloacae exhibited a notable 50% cefotaxime resistance in the combined fishes and pond water, while in the pond water alone, cefotaxime resistance increased to 75%, accompanied by a chloramphenicol resistance of 25%.
In the combined fishes and pond water, Enterobacter asburiae showed resistance rates of 25% for cefotaxime, 75% for chloramphenicol, and 50% for tetracycline. In the fishes only, the resistance was 33.3% for cefotaxime, 100% for chloramphenicol, and 66.7% for tetracycline. For Enterobacter hirae, high resistance rates were found in both the combined fishes and pond water, as well as in the fishes only, with 100% resistance to cefotaxime, chloramphenicol, and tetracycline.
Proteus mirabilis demonstrated resistance in the combined fishes and pond water at rates of 31.3% for cefotaxime and 100% for tetracycline. In the fishes only, the rates were 33.3% for cefotaxime and 100% for tetracycline. In the pond water only, the rates were 30% for cefotaxime and 100% for tetracycline.
In the case of Plesiomonas shigelloides, in the combined fishes and pond water, it exhibited resistance of 46.2% for cefotaxime and 38.5% for tetracycline. In the fishes only, the resistance was 50% for cefotaxime and 37.5% for tetracycline, while in pond water only, the resistance was 40% for both cefotaxime and tetracycline.
Vibrio albensis displayed various resistance in the combined fishes and pond water environments, with rates of 30% for cefotaxime, 30% for ciprofloxacin, and 50% for chloramphenicol. In the fishes, the resistance rates were 33.3% each for cefotaxime, ciprofloxacin, chloramphenicol, and tetracycline. In the pond water, the resistance included 28.6% each for cefotaxime and ciprofloxacin, and 57.1% for chloramphenicol.
Furthermore, in Bacillus cereus, in the combined fishes and pond water, resistance rates were 63.2% for cefotaxime and 42.1% for tetracycline. In the fishes alone, the resistance was 75% for cefotaxime and 37.5% for tetracycline. In the pond water, the resistance was 54.5% for cefotaxime and 45.5% for tetracycline.
Acinetobacter soli demonstrated 20% cefotaxime resistance in the combined fishes and pond water, while in fishes only, there was a higher cefotaxime resistance of 50%.
Moreover, Lactococcus latis displayed high cefotaxime resistance in the combined fishes and pond water (71.4%), with 50% resistance in the fishes only and 100% in the pond water only. Similarly, Pseudomonas putida showed cefotaxime resistance of 83.3% in the combined fishes and pond water, 100% in the fishes only, and 75% in the pond water. In like manner, Pseudomonas aeruginosa exhibited 85.7% cefotaxime resistance in the combined fishes and pond water, 100% in the fishes only, and 75% in the pond water only.
Lysinibacillus sphaericus demonstrated complete resistance (100%) to chloramphenicol and tetracycline in the combined fishes and pond water, as well as in the pond water alone, while in the fishes, the only 100% resistance was to cefotaxime.
Comamonas aquatica, Comamonas kerstersii, and Shewanella algae all displayed cefotaxime resistance, with rates of 100% in all settings – the combined fishes and pond water, the fishes only, and the pond water only.
Salmonella spp. exhibited equal resistance rates of 33% to cefotaxime, chloramphenicol, and tetracycline in the combined fishes and pond water, while in the pond water alone, the resistance increased to 50% for cefotaxime, chloramphenicol, and tetracycline.
Providencia alcalifaciens also displayed complete resistance to tetracycline in both the combined fishes and pond water and in the fishes only.
In terms of the prevalence of multidrug resistant (MDR) and ESBL-producing bacteria isolated, 13.6% and 1.3% of the total bacteria were MDR and ESBL producers, respectively (Table 6).
MDR and ESBL prevalence among the bacteria.

Categorically, 13.4% and 1.0% of the bacteria isolated from the fishes were MDR and ESBL producers, respectively. Similarly, bacteria isolated from the pond water recorded 13.7% and 1.6% for MDR and ESBL-producing bacteria, respectively. Among all the bacteria isolated, 20.6% and 7.4% of E. coli were MDR and ESBL-producing, with 19.6% and 6.5% isolated from the fishes and pond water, respectively. The overall MDR rate for A. veronii was 11.9%, while none was ESBL-producing. The percentage of MDR-A. veronii in the fishes and pond water were 8.0% and 17.6%, respectively.
Molecular epidemiology of the ESBL-producing isolates (sequence types, serotypes, resistance genes, and virulence genes)
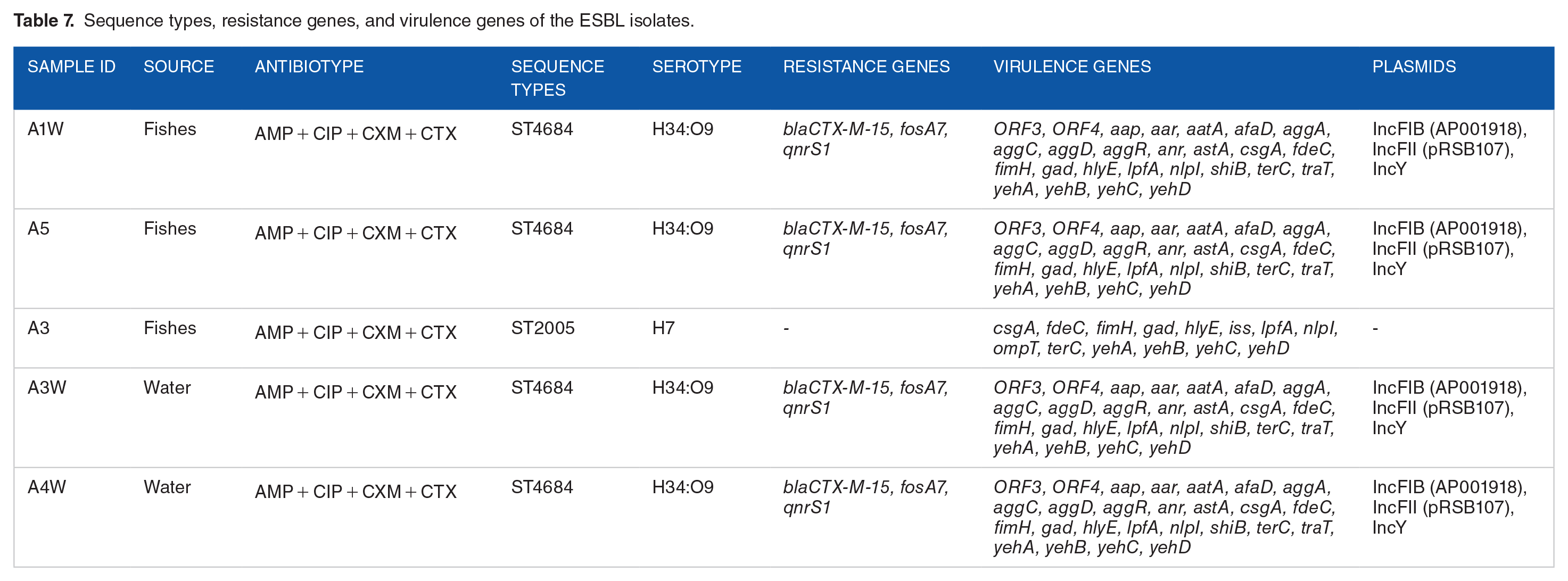
Table 7 shows the molecular epidemiology of the ESBL-producing isolates. These bacterial isolates (A1W, A5, A3, A3W, A4W), which were from the fishes and pond water, exhibited resistance to ampicillin, ciprofloxacin, cefuroxime, and cefotaxime, with specific genetic profiles including resistance genes blaCTX-M-15, fosA7, and qnrS1. Additionally, they carried virulence genes responsible for various functions, such as adhesive aggregative proteins (aap), aggregative adherence regulators (aar), Type VI secretion system effectors (aatA), Dr fimbrial adhesins (afaD), aggregation proteins and regulators (aggA, aggC, aggD, aggR), anaerobic regulation proteins (anr), heat-stable enterotoxins (astA), curli subunit proteins (csgA), ferrous iron dicitrate transport proteins (fdeC), Type 1 fimbrial proteins (fimH), glutamate decarboxylases (gad), haemolysin E proteins (hlyE), long polar fimbrial proteins (lpfA), lipoproteins (nlpI), outer membrane proteins (shiB, yehA, yehB, yehC, yehD), tellurite resistance proteins (terC), and serum resistance-associated proteins (traT). Notably, Isolate A3, originating from fishes, lacks serotype information but exhibits a distinct sequence type (ST2005). Samples A3W and A4W, sourced from water, share the same sequence type (ST4684) and display similar genetic characteristics.
Sequence types, resistance genes, and virulence genes of the ESBL isolates.
Discussion
The goal of this study was to evaluate antibiotic resistance in bacteria harboured in reared fishes in Accra and the pond water from which they originated, focusing on distribution of the harboured bacteria and their antibiotic resistance patterns, as well as their ESBL phenotypes, genotypes, and sequence types. The rich bacterial diversity in the fishes and pond water could possibly be accounted for by the different types of fish holding facilities used, namely concrete, earthen, plastic, and tarpaulin. It could also be attributed to the bioaccumulation of microorganisms within the fish. 19 This process allows for the concentration of bacteria and other microorganisms in the tissues of the fish, often in the gut, as well as their presence in the surrounding pond water. This bioaccumulation is particularly significant when considering the diet and feeding habits of the fishes – fishes consume various materials, including plankton, known to harbour high levels of bacteria. Notably, E. coli was prominent in the fishes and pond water, constituting 17% of the total isolates, signaling faecal contamination, 26 and raising concerns about water quality and hygiene practices in the farms. Adinortey et al., 19 in their report, pointed out that under stressful conditions, such as high faecal coliform levels in breeding ponds, bacteria may penetrate fish muscles, with the likelihood of muscle invasion increasing the longer fishes are exposed to polluted water. This observation is critical, as it suggests that not only external, but also internal tissues of fish can harbour diverse pathogenic bacteria.
Essentially, the highlighted concerns about the microbial safety of reared fishes are especially validated by similar observations on frequent occurrence of E. coli in aquaculture systems.27 -29 The presence of E. coli in both the fish and pond water samples underscores its wide distribution and potential for contamination throughout the aquaculture environment. Also, its high prevalence suggests the importance of implementing effective sanitation practices and water quality management to minimise the risk of faecal contamination and potential transmission of harmful strains. Enforcement of this recommendation is especially crucial, given the fact that these fishes (tilapia and catfish) are widely consumed in Ghana, and patronage of dishes from these grilled fishes is commonplace in the country. Thus, substandard cooking practices involving these fish could potentially endanger consumers.
To buttress the concerns about the microbial safety of the fishes and the potential risks they pose, Aeromonas hydrophila, a key opportunisitc pathogen, was the second most prevalent bacterium, accounting for 3% of the total isolates. It was primarily found in the fish samples, indicating its association with the fish microbiota. This finding is consistent with previous studies that have identified Aeromonas spp. as common inhabitants of reared fish.27,30 The higher prevalence of A. hydrophila in the fishes compared to the pond water samples suggests its potential transmission from the aquatic environment to the fish. The presence of Aeromonas spp. in the fishes raises concerns about their pathogenicity and their potential to cause opportunistic infections, particularly in stressed or immunocompromised fish. Proper monitoring and management strategies, including water quality control and stress reduction, are essential to minimise the risk of Aeromonas-related infections in aquaculture systems, as well as potential transmission to humans.
Also, Vibrio albensis was detected in 5% and 4% of the isolates, for the fish and pond water samples, respectively. Vibrio species are commonly associated with aquatic environments and have been previously reported in aquaculture settings.31,32 The presence of Vibrio albensis in this study highlights the importance of monitoring and controlling these pathogens to minimise potential health risks. However, it is crucial to note that the strains identified in this study may not necessarily be the toxigenic ones responsible for human cholera cases. Nonetheless, the presence of Vibrio species emphasises the need for proper hygiene practices, water treatment, and appropriate management strategies to reduce the risk of Vibrio-related infections in both fishes and humans.
Among the bacterial isolates, Klebsiella pneumoniae were predominantly found in the pond water samples rather than in the fish samples. This suggests that these bacteria may have a stronger association with the pond water environment. The higher prevalence of Klebsiella pneumoniae in pond samples aligns with the findings of Boulares et al., 27 who reported the presence of Klebsiella spp. in aquaculture systems. These bacteria may interact with the pond water, sediment, or associated microorganisms, highlighting the complexity of the microbial community dynamics in aquaculture systems.
Other bacterial species, such as Acinetobacter baumannii, Vibrio spp., Enterobacter cloacae, and Pseudomonas luteola, showed varying distributions between fish and pond water samples. This indicates their potential presence both in the fishes and their surrounding environment. The variations in distribution patterns among these bacteria highlight the intricate interplay between aquacultured organisms and their environment, which can influence the microbial composition and potential transmission of bacterial pathogens.
It is evident that the distribution and prevalence of bacterial species in aquaculture systems can vary depending on the specific environment, culture practices, and geographical location. Factors such as water source, feeding regimes, water quality management, and biosecurity measures can significantly influence the microbial ecology of aquaculture systems. Therefore, it is essential to consider these factors when interpreting and extrapolating results from different studies.
The presence of other pathogenic bacteria, such as Aeromonas hydrophila, Vibrio albensis, and Salmonella spp., among the isolates emphasises the importance of implementing appropriate management practices to mitigate the risk of bacterial contamination and potential transmission of antibiotic resistance genes. Aquaculture-associated bacterial pathogens can pose risks not only to fish health but also to human consumers through the consumption of contaminated seafood. Therefore, strategies such as regular monitoring of water quality, proper sanitation, adequate biosecurity measures, and responsible use of antibiotics are crucial for maintaining a safe and sustainable aquaculture industry.
Furthermore, the implications of antibiotic resistance in aquaculture cannot be overlooked. The presence of antibiotic-resistant bacteria, such as Acinetobacter baumannii and Klebsiella pneumoniae, in this study raises concerns about the potential transfer of antibiotic resistance genes to human pathogens. Antibiotic resistance is a global health issue, and the misuse or overuse of antibiotics in aquaculture can contribute to the emergence and dissemination of resistant strains. It is important to implement prudent and responsible use of antibiotics in aquaculture practices, in line with regulatory guidelines and best management practices, to minimise the selection and spread of antibiotic resistance.
It is worth noting that this study focused on a specific aquaculture system and a limited set of bacterial species. Further research is needed to explore the microbial ecology of different aquaculture systems, including marine, brackish, and freshwater environments, and to investigate a broader range of bacterial species. Additionally, the application of advanced molecular techniques, such as metagenomics and metatranscriptomics, can provide a more comprehensive understanding of the microbial community dynamics and functional interactions within aquaculture environments.
Generally, the antibiotic resistance rates were low to moderate, and were highest for tetracycline (31% to 42%), cefotaxime (29% to 35%), and chloramphenicol (19%), while no resistance was recorded against either meropenem or ertapenem. These results align with the global concern of widespread resistance to these antibiotics in various bacterial species. The high levels of resistance to tetracycline are consistent with the findings reported by Adinortey et al. 19 and Ginovyan et al. 33 in fish farms in Ghana and Armenia, respectively. The substantial resistance to cefotaxime highlights the need for careful consideration when using these antibiotics in aquaculture.
The prevalence variations between the fishes and pond water highlight the need for a more targeted investigation into the factors driving resistance in specific bacterial strains. This finding aligns with previous studies by Adinortey et al. 19 and Ginovyan et al., 33 which reported high levels of antibiotic resistance in coliform bacteria, including E. coli. Adinortey et al. 19 found that a significant proportion of coliform bacteria in fish farms in Ghana exhibited resistance to multiple antibiotics, including ampicillin. Similarly, Ginovyan et al. 33 highlighted the high resistance to ampicillin and other commonly used antibiotics among E. coli strains. The analysis of A. veronii revealed a moderate MDR prevalence (11.9%), with notable differences between the fishes (8.0%) and the pond water (17.6%). This emphasises the niche-specific nature of antibiotic resistance and underscores the importance of considering individual bacterial species when assessing resistance in aquatic environments.
This study provides evidence on the need to inculcate the One Health approach in managing antibiotic resistance in aquaculture. It can be seen from the results that the prevalence of antibiotic-resistant pathogens, antibiotic resistance genes, and virulence genes, in the gastrointestinal of the sampled fishes and the water within which the fishes were kept are determining factors for exacerbating AMR in the environment, in humans after their ingestion of improperly cooked infected fish, and that which is transmitted between and among fishes. However, a comprehensive study including the presence of the different type of antibiotics present in the soil, pond, and gut of fishes will be necessary to buttress this study, and to establish convincing evidence for the need for using One Health approach in tackling this aquaculture menace.
In line with the observed antibiotic resistance traits highlighted above, various antibiotic resistance-conferring genes were observed in this study, as has been the case of aquatic sediments in previous reports, showcasing the intricate nature of the issue. 34 Notably, this study identified the resistance genes blaCTX-M-15, fosA7, and qnrS1, in both fish and pond water samples. Among these, blaCTX-M-15 stood out as a widespread gene, encoding extended-spectrum β-lactamases with hydrolysing activity against cephalosporins.35,36 The fosA7 gene, on the other hand, encodes an enzyme that catalyses a chemical modification of fosfomycin. 37 This modification typically involves the addition of a chemical group to the antibiotic molecule, rendering it unable to inhibit its target enzyme effectively. As a result, bacteria carrying the fosA7 gene can neutralise the antibacterial effects of fosfomycin, leading to resistance.
Mechanisms of quinolone resistance frequently involve plasmid-encoded qnr proteins. In this study, the qnrs1 gene, a plasmid-mediated quinolone resistance gene was identified.38,39 Globally, there have been detections of plasmid-mediated quinolone resistance in bacteria associated with fish farms. 40 For example, in Chile, Buschmann et al. 41 reported the presence of qnrS in sediments impacted by fish farming. Also, in China, Jiang et al. 42 observed a high prevalence of qnrS genes in Escherichia coli from farmed fish, while Han et al. 43 found only a few strains with qnrS1-like genes in Aeromonas spp. isolated from diseased fish and water samples. Further studies on the mutagenesis of qnrS1 have shown that specific amino acid substitutions can affect its ability to confer resistance to antibiotics like ciprofloxacin. 44 The detection of these resistance genes in both fish and water samples points to a complex transmission network of antibiotic resistance within aquaculture systems, aligning with existing literature that highlight water bodies as conduits for antibiotic resistance spread, not only within aquaculture settings but also to surrounding environments.45,46
Two distinct serotypes, H7 and H34:O9, were identified in this study, with both serotypes present in fish samples, while only H34:O9 was found in the water samples. Serotype differentiation, based on specific flagellar (H) and somatic (O) antigens, reveals the diversity of bacterial strains in these environments. Specifically, serotype H34:O9 expresses the 34th flagellar antigen and the ninth somatic antigen, whereas serotype H7 only expresses the seventh flagellar antigen. 47 Although there is limited information on H34:O9 specifically, serogroup O9 was classified by Navarro et al. 48 as commensal strains containing diarrhoeagenic E. coli genes. Also, Ghilardi et al. 49 identified O142:H34 as one of the most frequent serogroups, also prevalent in diarrhoeagenic E. coli. Furthermore, the study by Onmaz et al. 50 reported an incidence of E. coli O157:H7 contamination in fish samples obtained from different fish farms and retail markets. These findings, however, contradict those of Fernandes et al., 51 who did not identify E. coli O157:H7 in 240 catfish fillet samples despite immunological testing with the 3 m Petri film HEC test kit and subsequent culture on 3 m Petri film E. coli plates.
The identified prevalence rates of antibiotic-resistant bacteria in aquatic environments have potential implications for public health. Also, the transfer of resistance genes from aquatic bacteria to human pathogens, through the food chain or waterborne routes, could contribute to the dissemination of antibiotic resistance in clinical settings. Furthermore, the discernible differences in resistance patterns between the fishes and pond water suggest that the aquatic habitat plays a role in shaping the antibiotic resistance landscape. Moreover, the slightly elevated prevalence of MDR in fishes compared to the overall sample underscores the potential influence of ecological factors on resistance dynamics within specific niches. In addition, the observed patterns of antibiotic resistance in the fish and pond water bacteria underscore the need for prudent antibiotic use in aquaculture. Plus, the resistance of bacteria to certain antibiotics, such as cefotaxime, highlights the potential challenges in disease management within this aquatic ecosystem. Besides, the effectiveness of gentamicin against a high percentage of bacteria isolated from both the fishes and pond water is promising and emphasises the importance of selecting antibiotics carefully in aquaculture practices.
Overall, this study has revealed a highly diverse microbial composition in aquaculture systems in Accra, Ghana, as well as patterns of antibiotic resistance genotypes and phenotypes. The presence of resistance genes and virulence factors suggests a complex interplay between genetic traits and selective pressures in the aquatic environment. It is noted, though, that the whole-genome sequence analysis was limited to the five ESBL-producing isolates recovered from the fishes and pond water in this study, all of which happened to be E. coli. Thus, antibiotic resistance traits in the isolates that were resistant to third-generation cephalosporins, but had no evidence of ESBL-producing capacity, may be mediated by other determinants, such as AmpC or other beta-lactamases, or even changes in outer membrane permeability. 52 Hence interpretation of the distribution of antibiotic resistance genes, as well as other insights revealed in the whole-genome sequence analysis ought to be interpreted in that limited context. Admittedly, understanding the molecular epidemiology of antibiotic-resistant bacteria is crucial for developing targeted strategies to control their spread and maintain the efficacy of antimicrobial treatments in aquaculture. It is, therefore, important that future antibiotic resistance surveillance efforts in aquaculture systems expand the scope of organisms subjected to the molecular characterisation.
Conclusions
The fishes and the pond water were contaminated with a broad range of bacteria, mainly Escherichia coli and Aeromonas veronii. The antibiotic resistance, MDR, and ESBL rates were low to moderate. Moreover, the main sequence type and serotype of the ESBL isolates were ST4684 and H34:09, respectively, and the antibiotic resistance genes were blaCTX-M-15, fosA7, and qnrS1.
Further research is needed to explore the microbial ecology of different aquaculture systems, including marine, brackish, and freshwater environments, and to investigate a broader range of bacterial species. Additionally, the application of advanced molecular techniques, such as metagenomics and metatranscriptomics, can provide a more comprehensive understanding of the microbial community dynamics and functional interactions within aquaculture environments. Furthermore, it is recommended that regulatory bodies enforce microbial food safety polices in the fish production chain and sensitise consumers on the need to cook their fish well to destroy contaminating bacteria.
Footnotes
Acknowledgements
We are grateful to the technical staff of the Department of Medical Microbiology, University of Ghana Medical School (UGMS), as well as those of the Bacteriology Department, Noguchi Memorial Institute for Medical Research (NMIMR), University of Ghana. We also appreciate the funding support from the SeQ-Africa (FF RGR2 FF25) Project which enabled the sequencing of the ESBL isolates.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Whole genome sequencing of the ESBL isolates was supported by the SeqAfrica Project (FF RGR2 FF25), funded by the Department of Health and Social Care’s Fleming Fund using UK Aid. The views expressed in this publication are those of the authors and not necessarily those of the UK Department of Health and Social Care or its Management Agent, Mott MacDonald.
Declaration Of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
Conceptualisation, N.T.K.D.D.; methodology, N.T.K.D.D., F.C.N.K., F.N.N.N.-M., B.E., B.K.A.T., A.D.D., P.B.T.-Q., and E.S.D.; software, B.K.A.T. and P.H.; validation, N.T.K.D.D., B.E., F.C.N.K., A.D.D., P.B.T.-Q., and E.S.D.; formal analysis, N.T.K.D.D., F.C.N.K., F.N.N.N.-M., B.E., B.K.A.T., A.O., and E.S.D.; investigation, N.T.K.D.D., F.C.N.K., F.N.N.N.-M., B.K.A.T., B.E., and A.O.; resources, N.T.K.D.D., F.C.N.K., F.N.N.N.-M., D.E.K.K., A.D.D., P.B.T.-Q., B.E., and E.S.D.; data curation, F.C.N.K., F.N.N.N.-M., B.E., B.K.A.T., D.E.K.K., A.O., P.H.; writing—original draft preparation, N.T.K.D.D., F.C.N.K., B.E., F.N.N.N.-M., P.H., A.D.D., and E.S.D.; writing—review and editing, N.T.K.D.D., F.C.N.K., F.N.N.N.-M., B.E., B.K.A.T., D.E.K.K., A.O., P.H., A.D.D., P.B.T.-Q., and E.S.D.; visualisation, N.T.K.D.D., F.C.N.K., F.N.N.N.-M., D.E.K.K., A.D.D., P.B.T.-Q., B.E., and E.S.D.; supervision, N.T.K.D.D., B.E., and E.S.D.; project administration, N.T.K.D.D., A.D.D., P.B.T.-Q., and E.S.D.; funding acquisition, B.E. All authors have read and agreed to the published version of the manuscript.”