
Abstract
The future global food security depends on the availability of water for agriculture. Yet, the ongoing rise in nonagricultural uses for water, such as urban and industrial uses, and growing environmental quality concerns have increased pressure of irrigation water demand and posed danger to food security. Nevertheless, its severity and duration are predicted to rise shortly. Drought pressure causes stunted growth, severe damage to photosynthesis activity, loss in crop yield, reduced seed germination, and reduced nutrient intake by plants. To overcome the effects of a devastating drought on plants, it is essential to think about the causes, mechanisms of action, and long-term agronomy management and genetics. As a result, there is an urgent need for long-term medication to deal with the harmful effects of drought pressure. The review focuses on the adverse impact of drought on the plant, physiological, and biochemical aspects, and management measures to control the severity of drought conditions. This article reviews the role of genome editing (GE) technologies such as CRISPR 9 (CRISPR-Cas9) related spaces and short palindromic relapse between proteins in reducing the effects of phytohormones, osmolytes, external compounds, proteins, microbes (plant growth-promoting microorganism [PGPM]), approach omics, and drought on plants that support plant growth. This research is to examine the potential of using the microbiome associated with plants for drought resistance and sustainable agriculture. Researchers also advocate using a mix of biotechnology, agronomic, and advanced GE technologies to create drought-tolerant plant varieties.
Keywords
Introduction
Population growth, drastic climate change, water supply, availability of workable land, and biodiversity losses have caused major changes to the global food security.1,2 Abiotic stress factors such as drought, salinity, floods, heat, cold, heavy metal pollution, toxic compounds, and biotic stress affect plant growth and yield.3,4 Drought pressures have become the most critical and critical constraints, and negatively affect agricultural output and ultimately endangering food security. 5 Drought affects agricultural output while exacerbating neglect, soil depletion, and environmental damage. Consequently, water scarcity has been identified as a significant worldwide ecological problem. 6 This abiotic stress leads to substantial physiological, metabolic, and molecular changes in plants, thus decreasing agricultural yields. 7 With current technologies, researchers have made tremendous progress in recent years to unravel the genetic, biochemical, and signaling mechanisms associated with plant stress responses.8-10 The complex process that distinguishes resistance and vulnerability within crop species is not well known. 11 Drought stress affects a variety of morphological biochemical parameters, including a decrease in the leaf area, a total reduction in chlorophyll content, leaf wilting, lengthening of the roots, and the formation of reactive oxygen species (ROS).4,12,13 Under stress, several osmotic pressure decomposes amass in plant cells, such as proline, glycine betaine (GB), soluble sugars, and spermines, to sustain cell penetration ability. 14 Catalase (CAT), superoxide dismutase (SOD), glutathione reductase (GR), and ascorbate peroxidase (APX) are examples of oxidation-protective enzymes, known to be altered in drought-stressed plants. 15 Drought stress, on the contrary, alters the expression of a large number of genes, including those involved in the transmission of stress signals; hundreds of functional proteins are also transcribed and controlled, which play a crucial role in drought-tolerant gene expression patterns. 16 As a result, to maintain or raise global food production, sustainable agricultural biotechnology must be developed to ameliorate abiotic stress. Various techniques, including genetic engineering and traditional plant breeding methods, have been used worldwide to improve crop drought tolerance.17,18
CRISPR/Cas9 is a gene-editing technology that enables precise changes to DNA in living organisms. It targets specific DNA sequences using RNA and Cas9 protein. This technology can correct genetic mutations, treat diseases, develop new therapies, and understand biological processes. It holds immense potential in medicine, agriculture, and biotechnology. CRISPR/Cas9 is a precise and effective technique that enables researchers to specifically alter an organism’s DNA. To make alterations, such as gene deletion, gene insertion, or gene editing, the Cas9 enzyme is guided by a guide RNA molecule to a specific site in the genome. 19
The genes involved in plant-microbe interactions can be altered using GE techniques like CRISPR/Cas9 to lessen the impact of plant growth-promoting microorganism (PGPM) on plants. For instance, by focusing on particular genes that recognize and react to particular phytohormones or osmolytes, researchers may be able to change how sensitive or responsive the plant is to these substances. 20
Genome editing can also be used to change genes that are involved in the absorption or metabolism of exogenous substances, such as pesticides or herbicides, lessening the effect these substances have on plants. It may be able to improve tolerance or resistance by carefully modifying the genes responsible for the plant’s reaction to these chemicals. 21
In addition, plants with improved resistance to hazardous bacteria or diseases can be created using GE technology. Researchers may be able to improve the plant’s capacity to identify and react to microbial invaders by focusing on specific genes involved in plant defense systems, which might lessen the impacts of PGPM. 22 It is important to note that the application of GE technologies in agriculture is still an active area of research and development. While these technologies hold immense promise, their use is subject to regulatory frameworks and ethical considerations. Ongoing studies are being conducted to further explore the potential benefits and implications of GE in reducing the effects of PGPM on plants. 23 It is significant to emphasize that research and development on the use of GE technologies in agriculture is currently ongoing. 24 Although these technologies have great potential, their usage is constrained by legal and moral standards. Research is ongoing to examine the possible advantages and consequences of GE in minimizing the impacts of PGPM on plants. 25 An introductory approach is to use drought-tolerant biological inoculations or plant growth-promoting rhizobacteria (PGPR) as an option for agricultural production in water-scarce areas. Various research suggested by researchers validates the idea that PGPR can help plants tolerate nonbiological stressors by altering their genetic responses. 26 Plant growth-promoting rhizobacteria promise to change the physiological response to water shortage and enhance their drought resistance. 27 Multidisciplinary solutions are needed to take full advantage of the potential of microorganisms in agricultural production systems. Data-driven poly-omic science improves our knowledge of microbe structure, dynamics, and composition and their physical function in a variety of designs and ecosystems, like root-to-root, where microbial interactions at the community level guide plant reactions to multiple pressures.28,29 However, given the complexity of plant-microbial interaction and its dependence on environmental variables, optimizing the relationship of plant microbes to increase drought tolerance in agricultural facilities is still a challenge. Developing drought-resistant plant genotypes, seed processing, genetic changes, plant-growing microorganisms, plant fertilizers, and appropriate dissolved substances are management approaches to promote drought tolerance in various agricultural facilities. Therefore, it is essential to have a thorough understanding of the possible improvement that can be attributed to plant-associated microbiomes on the production of agricultural plants as a mitigating technique for drought stress circumstances.
Drought Stress Effect on Plant Development
Drought is a period without rainfall, resulting in a considerable decrease in soil and relative humidity and a rise in atmospheric temperature.30,31 A mismatch between water input and soil steam flow can result in drought conditions. Plant health is directly and indirectly affected by drought stress. The penetration and ion balance of plant cells are affected by reducing the water availability of the roots and the loss of liquid during evapotranspiration, which leads to changes in the plant condition. 32 It also affects crop growth in various ways, like slowing down seed germination, which negatively impacts on crop yield. Drought is the primary principal abiotic stress that influences morphological, biochemical, physiological, and molecular reactions by affecting the activity of proteins in drought stress; drought stress can damage the morphology, biochemistry, and physiology of wheat plants. 33 Drought substantially influences leaf size, plant height, and crop development owing to silk division, reduced cell growth, and elongation of photosynthesis because the decline in leaf growth and leaves’ reduces the early stages of photosynthesis, thereby reducing the loss of food production. According to published studies, all essential activities, under drought stress, several processes, such as pure photosynthesis, the generation of photosynthetic pigments, electron transfer rates, the quantum efficiency of photosystem II, also known as PSII, is a crucial part of plants, algae, and some bacteria’s photosynthetic machinery. It consists of proteins, pigments, and cofactors that absorb light energy and convert it into chemical energy. The main pigment is chlorophyll a, which maximizes photosynthesis efficiency. The energy is then excited in a reaction center, facilitating the synthesis of ATP and the reduction of NADP+ to NADPH. Photosystem II also plays a role in water splitting through photolysis, releasing oxygen and contributing to the oxygen-rich atmosphere. Understanding this system is vital for bioenergy research, plant biology, and environmental studies (II optical systems (PSII), protein synthesis, lipids, and energy metabolism, are suppressed.34,35 Drought prevents plant growth by disrupting the plant’s water balance, membrane permeability, mineral nutrition, and enzyme activity, which substantially influence plant growth. 36
Drought pressure indirectly increases the production of ROS, which causes oxidative damage to the macromolecules. 37 Reactive oxygen species production leads to lipid oxidation, which causes damage to the membrane, enzyme inactivation, and structural and functional degradation of proteins. Protein oxidation and oxidative stress may result in the loss of enzyme activity and the creation of protease anticonvergent clumps. Therefore, plants often use various techniques to reduce ROS-mediated damage, such as the production of low molecular weight enzymes and zinc-free antioxidants. Other enzymes in the ascorbate-glutathione cycle, such as monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and GR, function to prevent dangerous oxygen derivatives from forming. 38
Various Techniques to the Amelioration of Drought Stress on Plants
Drought-resistant approaches have been demonstrated in a variety of studies, including (1) plant breeding and marker-assisted selection 39 ; (2) exogenously administered substances and osmolytes, such as polyols, glycine, betaine, proline, or other amino acids (sorbitol, pinitol, myoinositol, and mannitol), among others improve osmotic adjustment 40 ; (3) hormones produced by plants are (salicylic acid [SA], auxins, gibberellic acids [GAs], cytokinins [CKs], brassinosteroids [BRs], jasmonic acid [JA], and abscisic acid [ABA]) 41 ; and (4) antioxidant function (glutathione, polyamines, ascorbate, and enzymes) 42 . In the following subsections, we shall further dwell on some of the strategies to ameliorate drought stress.
Impact of phytohormones
Phytohormones play a crucial role in controlling plant responses to different types of drought. Plant hormones such as SA, JA, auxin, indole-3-acetic acid (IAA), ethylene (ET), CKs, BRs, and GAs are also needed for plants responsiveness to abiotic stresses. 43 The balance of auxin and CK, which regulates root growth and determines RSA, is a potent regulator of the plant organogenesis. 44 The endogenous ratio of auxin to CK in plants may be influenced by auxin and CK synthesis by rhizospheric bacteria and microbial metabolites that interact with these hormonal pathways. Low IAA levels may promote parent root elongation, while high IAA levels stimulate lateral root formation, root hair synthesis, and parent root growth. 45 Likewise, rice endophyte Phomopsis liquidambari B3 inoculation dramatically increased auxin, CK, and ET levels in rice at varied nitrogen levels. 46 Azospirillum brasilense also contains nitrite reductase activity and creates nitric oxide during root colonization, which mediate auxin signaling pathway to control the growth of LR. 47 Plant growth has been boosted by inoculating them with CK-producing rhizospheric bacteria. 48 Abscisic acid and GA, or substances that govern the concentration of these phytohormones in plants, are produced by rhizospheric bacteria, according to many studies.49,50 Rice root elongation is influenced by GAs, notably GA3, which affects local auxin production and the polar auxin transport. 48 Indole butyric acid (IBA) therapy has been shown to help mung bean seedlings overcome root development inhibition. 51 Surprisingly, rhizobacteria reduce drought-induced yield loss in plants by increasing auxin levels.52,53 Drought resistance is conferred by overexpression of sweet potato auxin response factor (ARF)5 in Arabidopsis through altering carotenoid biosynthesis, which is a precursor of IAA. 54 The co-expression of GA REQUIRING-5 (GA5) and DREB1A in Arabidopsis significantly increased drought tolerance without compromising growth or yield. 55
Drought triggers the production of the phytohormone ABA. Plant roots create ABA, which is then transferred to the leaves and used to regulate the stomatal aperture, SA and JA, in addition to ABA, are essential in the drought stress response. In addition, during drought, genes producing JA and SA are downregulated in the sweet sorghum. 56 Salicylic acid is a nonenzymatic antioxidant that helps remove ROS from the body. Salicylic acid-related gushing signals influence microorganisms’ whole-body resistance in the root and microbial selection mediated by host plants. 56 Sunflower (Helianthus annus) phytoremediation and drought tolerance improve when PGPR and SA are used in combined form.57,58 Methyl jasmonate (MeJA) function in enhancing drought stress tolerance has been explored in 5 different wheat growers, and it was discovered that MeJA efficiently mitigates the negative effects of drought stress.59,60 In stressed plants, glutathione is engaged in a variety of physiological tasks, including the elimination of toxic ROS and metal detoxification. Drought-induced oxidative stress can be reduced by combining seed and foliar treatments with MeJA (20 M) + SA (2 mM), which controlled ABA and osmolytes levels as well as antioxidant enzyme activity. 61
Impact of nanoparticles in the mitigation of drought stress
Due to the large surface area, varied pore size, and amorphous nature, nanoparticles (NPs) are very reactive. The contents of NPs are discharged from the organelles of plants. 62 Antioxidant enzymes, which include CAT, SOD, peroxidase, APX, and others, are regulated by them. Drought, heat, cold, salt stress, and heavy metal toxicity may all be reduced by silicon-based nanoparticles (SNPs). 63 Stress alleviation strategies include (1) plant antioxidant defense mechanisms are activated; (2) coprecipitation of Si with dangerous metal ions; (3) immobilization of hazardous metal ions in the growth medium; and (4) absorption and compartmentation of metal ions inside plants. Linh et al 64 discovered that metal-based (copper, iron, cobalt, and zinc oxide) NPs may improve soybean drought tolerance by increasing relative water content, biomass reduction rate, and drought tolerance index. Seed germination (%) and total germinate rate are improved by zinc oxide NPs have suggested the efficacy of ZnO NPs in using seed reservoirs for seedling development and drought stress tolerance. 65 Researchers employed chitosan NPs to change biochemistry and gene expression in Catharanthus roseus (L) to increase drought stress resistance.66-68
Exogenously administered chemicals in the alleviation of drought stress
To address global issues in agricultural activities, research must focus on the enhancement and application of exogenous hormones, and the establishment of crop management systems to promote the efficiency of water usage.93,94 Reactive oxygen species is a vital deliverable that can lead to oxidation signal conduction, systemic acquired resistance (SAR), and systemic acquired adaptation (SAA) corresponding to exculfactors. Under pressure, oxidation signal conduction causes a series of protective responses and yield management. Plant ROS (superoxide and hydrogen peroxide) oxidizing enzymes interact and are found in various plant tissues. Glutathione has a combined effect on methylglyoxal detoxification and antioxidative enzyme production in water-stressed mung bean plants. 95 Through similar research, glutathione-mediated drought stress resistance was connected to enhanced ionic homeostasis and decreased oxidative stress in rice (Oryza sativa L). 96 Silicon is a valuable ingredient that helps mitigate the negative impacts of drought. Fortified fertilizers have lately sparked attention due to their various functions, including boosting plant growth, photosynthetic machinery, and preserving cellular ion homeostasis under stress. 97 Various silicon exporters have indeed been identified in the monocot and dicot plants. 98 Diamond et al 99 found that integrating Si and Arbuscular mycorrhizal fungus (AMF) increased strawberry plant biomass by increasing water content, water consumption performance, antioxidant enzyme activity, photosynthetic rate, and notably zinc nutritional intake. Exogenous silicon treatment reduces oxidative damage caused by abiotic stressors in Brassica napus L 100 and wheat plants 97 by enhancing antioxidant enzyme activity. Selenium supplementation can cause an accumulation of solutes in grown plants in water-stressed conditions, lowering oxidative stress. In addition, selenium therapy can promote plant growth and antioxidant formation as they age and maintain a healthy water balance, enabling them to withstand drought stress. 101 Osmolytes come in a wide variety of sizes and shapes. Because of their cellular defense mechanisms against dehydration loss, osmoprotectants are also called osmolytes.
The involvement of osmolytes and phytohormones, and the interactions with some other chemicals in reducing drought stress in various plants.
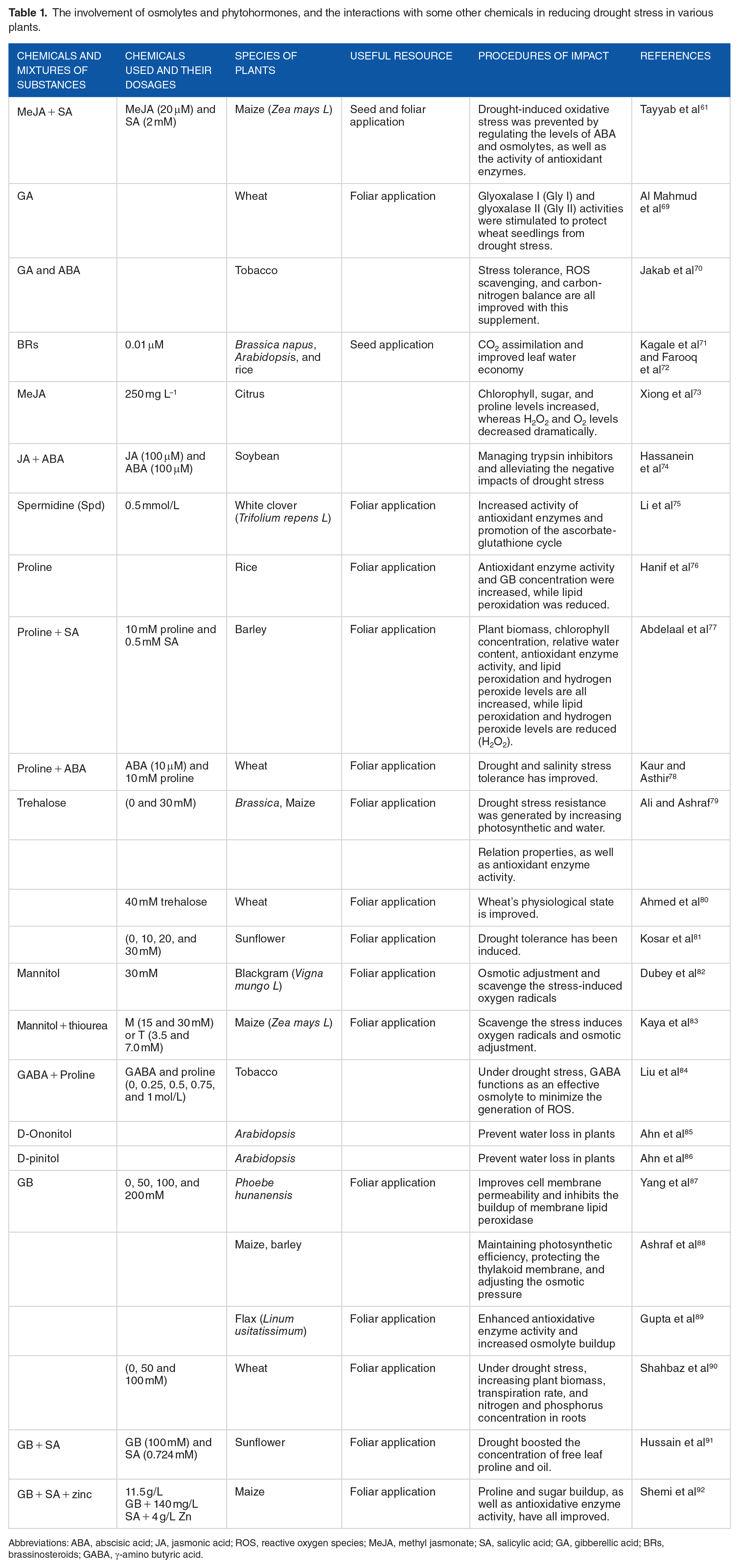
Abbreviations: ABA, abscisic acid; JA, jasmonic acid; ROS, reactive oxygen species; MeJA, methyl jasmonate; SA, salicylic acid; GA, gibberellic acid; BRs, brassinosteroids; GABA, γ-amino butyric acid.
Microbiome-induced resistance to drought stress
The plant microbiome is made up of bacteria found in the rhizosphere, endosphere, and phyllosphere, as well as other plant-related components, which include nectar and pollen.13,102 Plants produce a range of chemical compounds in their root exudate, which improves plant absorption, stimulates the development, and increases yield.103,104
Drought-tolerant microbiome plant recruitment may have developed through generations of recurring drought responses, resulting in positive and stable plant-microbe relationships that benefit both the microbe and the host plant.105,106 Drought-stressed PGPMs can be studied to discover which microbial features are advantageous to plants. Bacteria that promote plant growth (PG) have been discovered to increase nutrient bioavailability in soils for root absorption,18,107 especially in areas where there has been a protracted period of drought. 108 Plant growth-promoting rhizobacteria have been intensively examined in various crops, including soybeans, for their ability to mitigate the negative impacts of drought stress,4,109,110 wheat,111,112 chickpea,12,113 maize,114,115 rice, 116 and mung bean.117,118
Drought resistance is provided through several PGP abilities, the well-studied of which is the enzyme 1-aminocyclopropane-1-carboxylate deaminase.
In Vigna mungo L and Pisum sativum, ACCd-producing bacteria were shown to be beneficial in decreasing the negative impacts of drought stress,119,120 maize,4,121 soybean, 4 and Capsicum annuum. 122 Plant endophytes that dwell in the spaces between cells have been discovered to help plants cope with drought stress by generating a range of mechanisms that enable them to grow even when stressed.123,124 Even though drought has a considerable impact on plant-associated microbial communities, drought-tolerant microbiota has a significant beneficial impact on the phenological development of stressed plants.125,126 As a consequence, endophytes are considered among the most effective and widely accessible bioinput to increase plant drought tolerance. 4
Plants that are drought-stressed undergo various physiological, metabolic, and molecular changes, all of which are mediated or regulated by microbiota.
To deal with abiotic stress, endophytic microorganisms can serve as a source of several “protective” substances such as antioxidants, polysaccharides, and proline. 127
Stress tolerance seems to be conferred by microbial endophytes through the activation and modulation of stress-responsive genes, the production of metabolites that mitigate stress, and the production of ROS waste pickers. 128
Etesami and Maheshwari 125 found that enzymatic activity of SOD, CAT, and peroxidase was all higher in plants infected with beneficial microorganisms (ie, Bacillus cereus, B subtilis, and Serratia sp). Several enzymes have been reported to be responsible for the reduction in adverse effects of drought on plants, thus indicating that the species of bacteria might be used as tools to assist plants in dealing with drought stress and growing more effectively.129,130
Endophytic bacteria that produce melatonin, on the contrary, result in prolonged impacts on plant endogenous melatonin quantity when they invade parts of the plant. Endophytes have been shown to affect tissue osmoregulation, stomatal conductance regulation, and cell-wall elasticity maintenance, all of which help plants withstand drought. 4
Using proteomic and metabolomic approaches, P indica colonization of moisture-stressed barley roots increased the formation of proteins that protect photorespiration, energy management, primary metabolism, transporters, and autophagy, and during drought. 131 P indica increased the accumulation of proteins that protect photorespiration, energy management, primary metabolism, transporters, and autophagy. 131 The modifications in the host’s amino acid metabolism might explain the favorable impact of P indica invasion on autophagy in drought-stressed plants. 131
A long-term answer to the challenge of food security under abiotic stress settings is to integrate microbiota with agricultural practices. Notwithstanding, using these microbes in the field necessitates addressing a variety of issues, such as agronomic systems, microbe establishment, microbial inoculant invasion of soil and plant, and biosynthesis of bioactive metabolites primary and secondary metabolites involved in plant growth regulation, all of which have been done under a variety of soil types, climatic condition, and genotype conditions.29,132,133
Late embryogenesis abundant proteins
Late embryogenesis abundant (LEA) proteins have traditionally been detected in seeds, but they may also be found in the plant’s other reproductive tissues. Abscisic acid or ecological stress cues such as drought, low temperature, and salt, among others, activate the gene. They may act as water-holding molecules in ion sequestration and help in the stability of membranes and proteins, according to certain hypotheses. 134 Late embryogenesis abundant proteins have been examined in diverse plants by several researchers, 135 barley,135,136 rice, 137 and wheat, 138 to determine their role in drought stress tolerance.
Heat shock proteins
Plants in the field are subjected to a variety of biotic and abiotic stressors. 139 Plants react to these challenges by maintaining transcriptional and translational control over several metabolic processes. 140 It requires a team of molecular chaperones, like heat shock proteins (HSPs), to unravel the genetically diverse biological stress pathways. 141 Although HSPs are commonly overexpressed in plants reacting to abiotic stress, their functional involvement in stress tolerance has received little attention. Arabidopsis thaliana is an example of a plant, 142 soybean,143,144 and rice.145,146 The function of these HSPs has been investigated. Two nuclear/cytosolic HSPs identified in soybeans, GmDjp1 and DNA, have been connected to the abiotic 144 and biotic 143 stresses Heat shock protein90s, also known as glucose-regulated protein 94 (GRP94), are ER proteins that are essential for drought stress tolerance in drought-tolerant plants. Overexpression of SoyBiPD (soybean BiPs), another ER-resident HSP70s known as BiPs, enhanced drought stress resistance in tobacco (luminal-binding proteins).
Profiling impact of the unculturable microbiome in drought stress tolerance
Droughts are predicted to be more prolonged, frequent, and severe in the future than they have been in the past. It requires the development of innovative and fast-accessible technologies for enhancing and restoring drought resistance in crops. Plants and other land animals should cope with the drought created by diurnal and seasonal water changes.147,148 Plants are built to fight, endure, and avoid drying circumstances by changing their root architecture and development, altering their physiology, and closing their stomata on their aboveground segments. 149 Owing a better knowledge of the complex mechanism and feedback between plants and their microbial partners before and after drought would undoubtedly pave the way for better drought resistance in agricultural production by using the rhizosphere microbiome. Several studies have revealed that during drought, the microbiome of the plant root alters in favor of a few chosen microbial taxa, displacing the bulk of other microbial lineages.150,151
The increase in the number of genes involved in the breakdown of complex plant polysaccharides, indicating the development of oligotrophic bacteria, is mainly responsible for this change in microbial communities. 105 One reason for Gram-positive bacteria’s development in dry environments is that they are metabolically more active and hardier than Gram-negative bacteria. 105 Furthermore, research has shown that this abundance is linked to the length and severity of the drought and that it is thought to vanish whether water is reintroduced to the root system promptly. 108
Recent advances in omic biology, such as metagenomics, proteomics, and metabolomics, have allowed scientists to group genomes, proteomes, and metabolomes from different environments to understand better soil and plant-microbial dynamics, structure, and function communities, including secondary metabolite biosynthesis and genes.99,152 This research would benefit genes, glucose and secondary metabolites, transport, and metabolic pathways linked to bacterial enrichment under drought. In addition, the information obtained will help in the discovery of genetic features associated with these bacterial populations. The usefulness of multiomic approaches in understanding microbial features under drought has been proven in many recent studies 153 Researchers investigated the dynamics of iron metabolism in a drought-induced rhizosphere microbiome using metagenome-guided comparative genomics.9,154 They discovered a link between drought, iron metabolism, and the plant root microbiome. 154 Michaletti et al 155 looked at some of the critical metabolites that may be used to construct models that show the association between yield-related parameters and different metabolic pathways while analyzing the proteome and metabolomics of spring wheat leaf tissues. Researchers studied soybean’s metabolomic and transcriptome profiles treated with melatonin throughout the grain filling stage under drought stress conditions via modifying secondary metabolite formation pathways. 156
Utilization of genome editing technologies and tools
The utilization of GE technologies and tools has revolutionized the field of genetics and has had a profound impact on various areas of research and applications. 157 Genome editing refers to the precise modification of an organism’s DNA, allowing scientists to make targeted changes to specific genes or genetic sequences. 158 One of the most widely used GE technologies is CRISPR-Cas9. CRISPR-Cas9 is a versatile and efficient tool that uses a guide RNA molecule to direct the Cas9 enzyme to a specific DNA sequence, where it can make precise cuts or introduce desired changes. This technology has greatly simplified the process of GE and has made it more accessible to researchers across different disciplines. The utilization of GE technologies has opened up new possibilities in agriculture. Researchers are using these tools to develop crops with improved traits, such as increased yield, enhanced nutritional content, and resistance to pests, diseases, and environmental stresses. 159 Genome editing has the potential to accelerate the breeding process and reduce the time required to develop new crop varieties. 160 Genome editing technologies are also being used in basic research to study gene function and understand the underlying mechanisms of various biological processes. 161 By selectively modifying genes in model organisms, scientists can gain insights into the roles of specific genes and their contributions to development, disease, and other biological phenomena. 162 However, it is important to note that the utilization of GE technologies also raises ethical and societal considerations. The potential for off-target effects and unintended consequences of GE must be carefully evaluated and addressed. 163 There is an ongoing need for responsible and transparent governance to ensure that these technologies are used ethically and for the benefit of society. 164 It is challenging to decipher the processes behind the harm caused by drought stress for the development of drought-tolerant plant. As a result, introducing the desired drought stress-resistant traits into many crops will be simpler by using transgenic or GE technologies. CRISPR has emerged as a cutting-edge method for GE in plants, allowing them to generate desirable traits and boost crop output under abiotic stress situations (Figure 1).

Utilization of genome modification tools and technologies.
CRISPR/Cas9 has been found in a range of crops lately, including bread wheat,165,166 potato, 167 and model plants including Arabidopsis thaliana 168 and rice. 169 Increased expression of a variety of transcription factors (TFs) and genes involved in drought stress signaling aids plants in accumulating various metabolites, signaling molecules, and osmolytes, increasing drought stress tolerance. Plant dryness is caused by sensitive (S) gene expression, which causes hormonal imbalance, increases the formation of ROS, and decreases antioxidant activity. The expression of TF genes may also be through GE. The AREB1 TF gene’s promoter activity was enhanced in Arabidopsis using a modified Cas9 (dCas9) linked with the histone acetylation transferase domain. Through the positive regulation of drought-tolerant genes, the genome-edited lines demonstrated increased transcription of the AREB1 gene and acquired drought tolerance. 170 Plant genetic engineering has been transformed because of CRISPR technology’s efficiency, resilience, and ease of engineering. The current status of this technology can revolutionize agriculture for improved crop production, increase crop resistance to biotic and abiotic challenges, and increase climate change resilience. Drought, heat, and salt are all abiotic stress elements that considerably impact crop output across the globe. Various complicated signaling networks regulate plant responses to stress stimuli. CRISPR technology’s enhanced tools, such as base and prime editing, have considerably increased GE possibilities in agricultural plants without the requirement for DSBs. 171 The CRISPR-Cas technology has opened up new possibilities in wheat research for improving grain quality. The TaGASR7 gene, linked to grain length, is part of the Snakin/GASA gene family. 172 A CRISPR/Cas9 system that targeted TaGASR7 via the shoot apical meristem produced 11 mutant plants with desired alleles, and 3 plants passed the mutation down to the next cycle. 173 The TaGW2 gene, which encodes RING-type E3 ubiquitin ligase and is believed to be a negative regulator of wheat grain size and 1000-grain weight, was targeted using the CRISPR/Cas9 method. All 3 copies of the TaGW2 gene were mutated in the T1 knockout plant.174,175 Compared with wild-type plants, mutants exhibited dramatically enhanced attributes such as 1000-grain weight, grain area, grain breadth, and grain length. Wheat grains with a lower immunogenic response were obtained using CRISPR/Cas9 technology. 172 The ARGOS family genes, specifically the Auxin-Regulated Gene Involved under Organ Size 8 (ARGOS8), block the ET signaling pathway in drought circumstances resulting in increased yield and drought stress tolerance. In maize, on the contrary, endogenous expression of the ARGOS8 mRNA is very variable and low, according to the research. 176 Understanding gene function and the mutation necessary to bring about the desired change would be a key hurdle for successfully using CRISPR for crop improvement. It is worth noting that the targets and features generated using CRISPR are from functional research on model plants. CRISPR success must, however, be expanded to field crops to solve the issues of boosting food supply in the face of climate change. Similarly, in each plant species, genetic transformation processes are confined to a few genotypes referred to as “models” These plants often exhibit poor agronomic performance, like low yield, and are sensitive to several biotic and abiotic stressors. Introducing the CRISPR platform to field crops would save years of breeding time, but the barrier will remain a significant barrier to fully achieving the CRISPR promise. 177 Overall, these GE methods will open up new avenues for nucleotide-specific alterations. They may be actively researched in the future to enhance drought tolerance in crops, making them a powerful weapon for maintaining global food security.
Conclusions
Conclusively, the plant-associated microbiome has demonstrated potential qualities in reducing the stress caused by drought on agricultural plants. Improved drought resistance in plants may be a result of its capacity to increase water intake and retention, create stress protectants, and strengthen defense mechanisms against oxidative stress. It is crucial to remember that the microbiome’s efficacy might vary based on a number of circumstances. To fully use the potential of the plant-associated microbiome for sustainable agriculture and drought control, further study and knowledge of the individual microorganisms and their interactions with various plant species and environmental circumstances are required. Many strategies are needed to address the adverse effects of drought stress on agricultural plants, including PGPR and endophytes, seed lining with exogenous chemicals, growth hormones, biostimulants, and genetic modification. Using an omic approach that includes genomics, metabolomics, proteomics, and transcriptomics can help us better understand critical drought-tolerant genes and decipher complex signaling steps and gene structures associated with drought tolerance in many plants. Drought-tolerant products can be developed with the help of revolutionary methods like CRISPR/Cas9 GE tools, minimizing the global risk of food insecurity. Furthermore, learning more about the mechanisms through which soil microorganisms impact plant drought tolerance and recovery, as well as their interest and use in field situations, has a lot to say about increasing drought resistance in agricultural production systems. Novel treatments that incorporate physiological, molecular, and genetic methodologies would be advantageous to combat stress-related diseases.
Footnotes
Acknowledgements
VFA would like to thank the North-West University for the postgraduate doctoral bursary.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: OOB recognized the National Research Foundation of South Africa for the grants (UID: 123634; 132595) that support work in her research group.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
VFA and OOB had the idea for the review article and suggested the review topic. VFA performed the literature search and wrote the first draft. OOB made substantial and technical contributions to the structure of the various manuscript drafts. Both authors read and approved the final manuscript.