
Abstract
Cap color is an important commercial trait for oyster mushrooms. Various pigment constituents determine a diverse color. However, the pigments of oyster mushrooms are still ambiguous. The pink oyster mushroom (Pleurotus salmoneostramineus or Pleurotus djamor) chromoprotein is one of the few proteins belonging to this fungus that has a record of its sequence of amino acid residues. However, even though there are studies about this chromoprotein isolation, purification, and crystallization, the current information focused on its 3-dimensional model and the cofactor and prosthetic group (3H-indol-3-one) binding sites is unreliable and fragmented. Therefore, in this study, using free online servers such as Prot pi, GalaxyWEB, MIB, and CB-Dock2, a structural analysis and the prediction of its physicochemical and biological properties were conducted, to understand the possible function of this chromoprotein. The obtained results showed that this molecule is a protein with a molecular weight of 23 712.5 Da, an isoelectric point of 7.505, with oligomerization capacity in a dimer and glycation in the Ser6 residue. In addition, the participation of the residues Leu5, Leu8, Lys211, Ala214, and Gln215 in the binding of the prosthetic group to the protein was highlighted; as well as Ser6 and Pro7 are important residues for the interaction of the Mg2+ ion and eumelanin. Likewise, morphological changes based on different culture conditions (light/dark) showed that this protein is constitutive expressed and independent of blue light. The findings in this study demonstrate that pink chromoprotein is a melanosomal protein, and it possibly has a critical role in melanogenesis and the melanin polymerization. However, more experimental studies are needed to predict a possible mechanism of action and type of enzymatic activity.
Introduction
Oyster mushrooms are grown commercially all over the world, particularly in many developing countries, due to their ease of cultivation and high biological efficiency. One of the most important commercial characteristics of mushrooms is cap color. Fresh oyster mushrooms come in a variety of cap colors, ranging from gray, brown, light brown, yellow, red, and cream to pure white. The cap color of mushrooms is determined by a variety of factors, some of which may also be involved in discoloration after mechanical damage. 1
Fungi commonly produce aromatic polyketide pigments such as melanins, quinones, flavins, ankaflavin, anthraquinone, and naphthoquinone. 2 Mushroom dye chromophores contain a wide range of intriguing organic compounds. Their pigmentation varies with age, and some undergo distinct color changes when bruised; thus, the colors of mushrooms are an important feature used in their identification. Furthermore, mushroom pigments may protect the organism from UV damage and bacterial attack, as well as serve as insect attractants. 1
Pleurotus djamor var. salmoneostramineus is a pink-colored fruiting body-forming basidiomycete. This basidiomycetous fungus is simple to grow in the laboratory and produces a distinctive pink pigment when forming the fruit body primordia.3,4 Regarding the chromoprotein of P. salmoneostramineus (PsPCP), its color is due to the presence of a molecule of 3H-indol-one; 5 it has also been suggested that it has a cofactor due to its possible binding to metal ions such as Zn2+, Fe2+, and Cu2+. 3 It is important to highlight that when this protein is found in an aqueous medium and is irradiated by light, there is oxygen production, so it is likely to participate in some photosynthetic process. 5 Moreover, Zhang et al 6 recently extracted and identified pigments from oyster mushroom species with black, yellow, and pink caps. Using a panel of spectroscopic and physical/imaging techniques, the extracted pigments revealed 3 color kinds correlating to the cap color, which were all identified as melanin. Despite this, high-performance liquid chromatography (HPLC) and elemental analysis revealed that the melanin in oyster mushrooms was a blend of eumelanin and phaeomelanin. According to electron microscopy research, the melanin units are most likely found in the cell wall, as observed in other fungus. The color variation in oyster mushroom caps was caused by differences in the amounts and relative proportions of eumelanin and phaeomelanin.
As the number of protein structures rapidly increases, methods are required to efficiently exploit structural information for biological and pharmaceutical purposes. In the database of the National Library of Medicine (NIH), in the genome of PsPCP are registered 10 073 potential open reading frames that possibly express a protein. However, only 15 amino acid sequences of different proteins are identified, such as glyceraldehyde-3-phosphate dehydrogenase, metalloprotease, chromoprotein, partial topoisomerase II, subunit 1 of cytochrome C oxidase partial, and various laccases. 7
Protein function prediction methods are techniques used by bioinformatics researchers to assign biological or biochemical roles to proteins. These proteins are typically understudied or poorly predicted based on genomic sequence data. However, because homologous proteins frequently have distinct functions, predicting protein function from sequence and structure is a difficult problem. There are numerous methods for predicting protein functions based on sequence-based features, protein-protein interaction networks, protein structure, or literature.
The gene sequence of PsPCP was determined by Fukuta et al, 3 and it is possible to access it (ACCESSION: LC269947) beside the amino acid sequence it encodes, although more than 2 decades ago this chromoprotein was crystallized, 8 there is only one unreported record of its possible 3-dimensional structure in UNIPROT (ACCESSION: A0A2Z5U5S2_9AGAR) generated by AlphaFold with a low confidence level of around 50 to 70 predicted local distance difference test (pLDDT) so it is not a reliable model, 9 in addition to the fact that the binding sites of the cofactor and the prosthetic group in the apoprotein are not described.
The aim of the present work has been to better understand the structure and possible function of this chromoprotein and perform an analysis of its structure and predict its physicochemical and biological properties, using a series of free online servers headed mainly by AlphaFold, GalaxyWEB, and CB-Dock2. In addition, studies were conducted on the morphology of the fungus in light and dark conditions to determine the possibility that this protein works as a photoreceptor in vivo.
Materials and Methods
Prediction of the physicochemical properties of PsPCP
The amino acid sequence of the PsPCP was obtained from the DNA Data Bank of Japan (ID: LC269947), which was entered into the Prot pi program (https://www.protpi.ch/), to predict the physicochemical parameters of the protein, among which the isoelectric point, molecular mass, and absorption coefficient stand out. In turn, the DeepRex-WS server was used to study solvent exposure of the residues that make up this protein. 10 The prediction of secondary structure (beta sheets, alpha-helices, and spirals) and protein glycosylation sites were determined by PsiPred (http://globin.bio.warwick.ac.uk/psipred/) and DictyOGlyc 1.1 (http://cbs.dtu.dk/services/DictyOGlyc/), respectively.11,12 Finally, we employ the servers DeepLoc (cbs.dtu.dk/services/DeepLoc/) and CELLO v.2.5 (http://cello.life.nctu.edu.tw/) for the subcellular localization prediction of the PsPCP.13-15
Structural analysis of the PsPCP
To perform this analysis, a 3-dimensional model of the protein was generated using the UniProt (http://www.uniprot.org/) and AlphaFold (https://alphafold.ebi.ac.uk/) servers.9,16 Once obtained, the model underwent a refinement process using the GalaxyRefine function of the GalaxyWEB program (https://galaxy.seoklab.org/index.html). 17 The stereochemical quality of the model was evaluated using the UCLA-DOE LAB SAVE v6.0 servers ERRAT, Verify3D, PROVE, WHATCHECK, and PROCHECK.18-20 The best model with the best quality was used to explore the possibility of oligomerization of this structure with the GalaxyHomomer function of GalaxyWEB. Also, a characterization of the internal cavity of this protein was performed with the CICLOP server (https://ciclop.raylab.iiitd.edu.in/). 21
In addition, a prediction of binding of the main metal ions such as Cu2+, Fe2+, Zn2+, Mg2+, Ni2+, and Mn2+ was performed with the MIB program (http://bioinfo.cmu.edu.tw/MIB/). 22 Finally, a molecular coupling of the ligand 3H-indol-3-one (PubChem ID: 319297676) was performed in cavities 1 and 2 of the structure, using CB-Dock2 (https://cadd.labshare.cn/cb-dock2/php/index.php), 23 the preparation of the ligand conformer was performed through the online platform Cheminformatics (https://chemoinfo.ipmc.cnrs.fr/).
Morphological studies of PsPCP
Various lignocellulosic materials have been used; 24 in this work, the base medium prepared was sawdust and wheat bran in a weight ratio of 1:3, respectively, which was pasteurized in hot water at approximately 65°C and maintaining a humidity of 65%. 25 The inoculum of the strain P. djamor var. salmoneostramineus N-S419 (Hongos Ndexi, Texcoco, Estado de México) was grown in the middle of PDA (Potato Dextrose Agar). The substrate (cereal straw mixed with lignin-cellulosic and protein additives) was inoculated on the base medium and incubated at 24°C under conditions of relative humidity of 90% until the formation of fruiting bodies (approximately 1 month). 3
Results
The chromoprotein of PsPCP has been purified and crystallized; these crystals were reported with a resolution of 1.8°A in an X-ray synchroton. 8 However, the 3-dimensional structure is not found in databases such as Protein Data Bank (PDB), and in UniProt, there is a 3-dimensional model generated by AlphaFold, but it is incomplete as it lacks the prosthetic group that gives it the characteristic pink color and its cofactor. 5 Due to the lack of information in this context, in the present work, it was decided to use various bioinformatics tools to determine the physicochemical properties, the secondary structure, and the refinement of the 3-dimensional model, the molecular coupling, and the oligometric state of the PsPCP (Figure 1).

Flowchart for the in silico characterization of PsPCP.
The prediction of the physicochemical properties of PsPCP showed that it is formed by 226 amino acid residues presenting a high amount of alanine (42 residues, 18.58%), serine (26 residues, 11.5%), glutamine (17 residues, 7.52%), leucine (17 residues, 7.52%), lysine (21 residues, 9.29%), aspartic acid (8 residues, 3.54%), glutamic acid (14 residues, 6.19%), valine (11 residues, 4.87%), threonine (11 residues, 4.87%). In addition, it has an isoelectric point of 7.505, a net charge (z) at pH 7.4 of 0.116, a molecular weight of 23 712.5 Da, and an absorption coefficient at 280 nm of 21 430 M−1 cm−1. Finally, the DeepRex-WS server predicted that the chromoprotein has 68.58% of residues exposed to the solvent and 31.42% of the residues are hidden.
Evaluation of secondary structure content with the PsiPred program showed that PsPCPs contain 11 alpha-helix segments (pink) and 8 turns (light gray) of different lengths (Figure 2).

Prediction of the secondary structure elements of the PsPCP with the PsiPred server.
In the AlphaFold Protein Structure database, a 3-dimensional model of the chromoprotein was found with UniProt ID: A0A2Z5U5S2 (Figure 3). This model presents the following residue confidence score (pLDDT) values: 60% of the protein is in yellow, as it has a low score (70 > pLDDT > 50), and in orange, the remaining 35% has a very low score (pLDDT < 50). It is important to mention that the low prediction values are due to the fact that there is no mold with a high degree of similarity in the PDB and the tempered ones found in Swiss Model, eg, only have an identity percentage between 13.89% for a mitochondrial ribosomal protein (6ydw.16.A) and 6.45% for the methyl acceptor chemotaxis protein (3g6b.1B). Therefore, it was decided to refine the existing model (Figure 3).

Three-dimensional structure of the PsPCP monomer taken from the AlphaFold database.
The structure tuning was performed with the GalaxyRefine function, and each of them was evaluated with UCLA-DOE LAB SAVE v6.0 (ERRAT, Verify3D, PROVE, WHATCHECK, and PROCHECK). All the proposed models presented a better-quality factor value than the AF-A0A2Z5U5S2 model. In addition, models 1 and 5 were the best proposals for each of the evaluated parameters (Table 1). Therefore, model 1 was selected and used to analyze the degree of oligomerization and the binding site of 3H-indol-3-one in the protein structure.
Tuning parameters of the monomeric 3-dimensional model of the PsPCP.

Abbreviation: PsPCP, Pleurotus salmoneostramineus.
It is important to mention that due to the high percentage of hydrophobic amino acids, alpha-helices, and the cyclic nature of the prosthetic group, this protein has a high probability of forming oligomers. 26 Therefore, it was evaluated if the monomer is capable of oligomerizing, using the GalaxyHomomer function. The server generated 5 models, and in this case, the first 4 were dimers and the fifth model turned out to be an octamer (Figure 4).

Three-dimensional models of the possible oligomerization of PsPCP. Each monomer is shown in distinct color.
The characteristics of the 5 oligomeric models are represented in Table 2, which are listed according to the interface area and the modeling score. The data showed that model 1 presented the best evaluation; therefore, it was used to identify the site of interaction with the prosthetic group.
Results of PsPCP oligomerization prediction.

Abbreviation: PsPCP, Pleurotus salmoneostramineus.
For the subsequent analysis on the binding of the 3H-indol-3-one as prosthetic group to the protein, the CB-Dock2 free server was used, which performs a blind molecular modeling and detects the different cavities of the protein that are potential ligand binding sites that may or may not be part of the active site.
Figure 5 shows the 2 main cavities of the PsPCP dimer. The first cavity (C1) has a volume of 1541 Å 3 , and the coordinates of the center were X = −22, Y = −14, Z = 13, and the size of the cavity has the coordinates X = 22, Y = 19, Z = 21. In the case of the second cavity (C2), its volume was 1402 Å 3 and its center coordinates were X = −15, Y = −5, Z = 5, and the size of the cavity presented the coordinates X = 17, Y = 19, Z = 24.

The amino acid residues shown in red belong to cavity 2 and those in blue to cavity 2 into PsPCP.
The ligand obtained the same docking score in both cavities (−5.3 kcal/mol), forming hydrophobic-type interactions with equivalent amino acids in both subunits (Figure 6). Moreover, in both cases, interactions were formed with 2 residues of the other cavity (Pro10 and Ala218). It should be noted that within the residues that participated in the ligand binding, Ser6 is found, which suggests that it could participate in the biological function of PsPCP. 27 Therefore, these results suggest that PsPCP must has 2 3H-indol-3-one molecules in its structure located at the dimer interface.

Localization of the binding sites of the prosthetic group by molecular docking of the PsPCP.
However, according to the previous published works,3,5 this chromoprotein can also bind cofactors, and among them, Zn2+ is suggested as the most viable candidate. Therefore, the MIB server was used to predict if any metal ions can be bound to the PsPCP. The 6 metal ions generally found in metalloproteins were analyzed. 28 The metals that presented the best score were Zn2+, Mn2+, and Mg2+ (Table 3). It is important to mention that the Mg2+ ion binds to the residues Ser6 and Pro7, which are part of the cavities where 3H-indol-3-one binds (Figure 8). Whereas Mn2+ interacts with Asp196 and Glu199. Finally, Zn2+ interacts with Tre213 and Glu217.
Metal ion binding sites on the PsPCP by MIB.
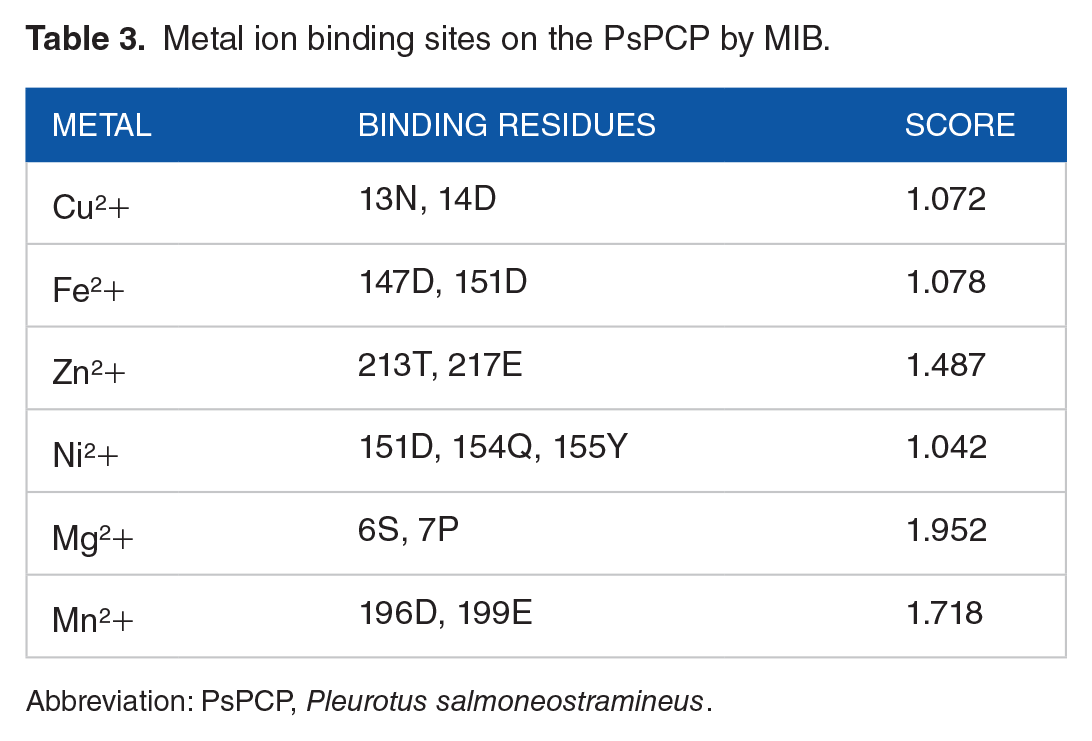
Abbreviation: PsPCP, Pleurotus salmoneostramineus.
Post-translational modifications are the mechanisms that modulate or regulate the function of proteins, especially those with catalytic activity. Therefore, using the DictyOGlyc-1.1 server, the possible glycation sites in the PsPCP sequence were analyzed. The data showed that there are several residues that could be glycated; however, only the Ser6 residue exceeds the threshold of relevance, and therefore, it is the only one that can be modified by glycation according to the criteria of this program (Figure 7). It is important to mention that Ser6 is part of the binding sites of 3H-indol-3-one and also this residue participates in the binding to the cofactor which reinforces that it can be a critical amino acid for the function of the protein and its chemical modification by glycation can have a potential effect on the structure.

Prediction of potential PsPCP glycation sites.
In addition, the possibility of both cavities communicating through a tunnel or channel was analyzed, and to evaluate this possibility, the CICLOP server was used (Figure 8). This program detects a tunnel that can be physiologically relevant as it maintains a diameter between 1 and 6 Å which allows to maintain a microenvironment to avoid exposure of waste to solvent or oxygen, which can potentially generate the protein itself in the presence of water. 5 In addition, it has 2 cavities with a volume of about 400 Å 3 in the central part of the protein. To our surprise, the protein has a tunnel along the dimer; however, it does not seem to include the 3H-indole-3-one binding sites. It is important to mention that the N-terminal domain of each subunit functions as an arm that traps the prosthetic group (Figure 8). In addition, Mg2+ ions bind to the same N-terminal domain and interact with the same prosthetic group binding site cavity. Therefore, there is a proximity between the 3H-indole-3-one and the Mg2+ ion that could be key in catalysis; however, it was not possible with the results of this in silico study to predict a possible mechanism of action and what type of enzyme function this chromoprotein may develop or whether it acts as a photoreceptor. In general, the chromoproteins are characterized by an electronic adsorption band in the UV-near, visible, and near-infrared range. These bands may arise from the π → π* transitions of the prosthetic group (3H-indole-3-one) or from the charge transfer transitions of specifically bonded transition metal ions (Mg2+ ion).

Identification of the internal tunnel of the PsPCP dimer and the central alignment for 3H-indole-3-one binding sites and Mg2+ ions.
To know the subcellular location of the PsPCP, it was decided to use the DeepLoc-1.0 server to predict whether this protein could be a photoreceptor and what type of light it uses. The results suggest that our protein is soluble and that its most probable location is the cytoplasm with a score of 0.591, whereas the second possibility was the nucleus with a value of 0.339 and the third place was the mitochondria with a score of 0.042. These results were verified by CELLO2GO and CELLO v2.5 server who revealed that PsPCP is predicted to be localized in the nucleus of the cell or is an extracellular protein (Figure 9). Therefore, this protein could be a blue light photoreceptor.

Prediction of PsPCP subcellular localization using DeepLoc-1.0 and CELLO2GO.
Finally, to evaluate the possibility that PsPCP functioned as a blue light photoreceptor, it was decided to cultivate PsPCP in the presence of blue light or darkness. The results suggest that the chromoprotein is produced with the same intensity in the presence or absence of blue light; however, its morphology is affected by darkness (Figure 10). Therefore, these results suggest that PsPCP may not have a primary photoreceptor function and may be a potential electron-transferring enzyme.
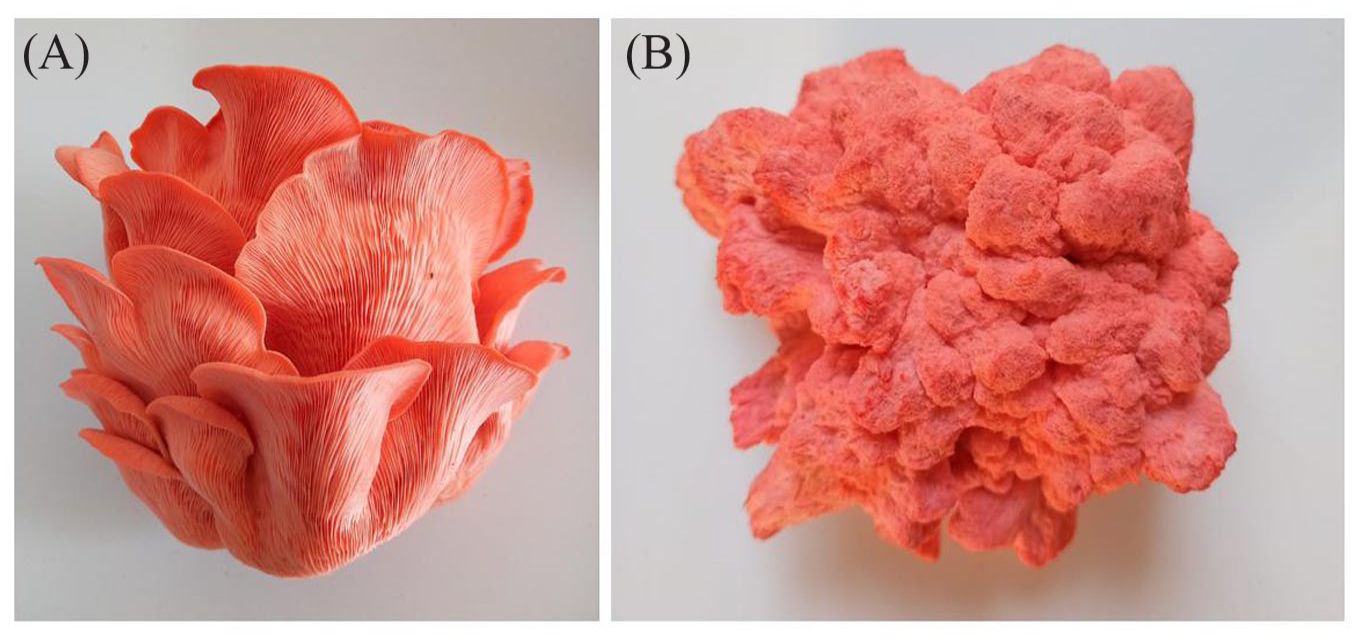
Morphological changes in the fruiting bodies of P. salmoneostramineus cultivated in the presence of blue light (A) and darkness (B).
According to recent research, fungal melanin can be synthesized in internal vesicles like mammalian melanosomes and transported to the cell wall. 29 The mammalian melanin pigments are synthesized in melanosomes, a specialized lysosome-related organelle in which the pigment is deposited on fibrils made up of proteolytic fragments derived from the pigment-cell-specific protein PMEL17. The PMEL17 is an integral membrane protein that is transported from the endoplasmic reticulum to the Golgi apparatus, where it is glycosylated before being trafficked to the melanosome. 30 Then, we hypothesized that PsPCP could function similarly to PMEL17 and decided to see if melanins could interact with the pink chromoprotein. As a result, we used the Zhang data to create the 3-dimensional structure of the eumelanin and pheomelanin fragment. Figure 11A shows that eumelanin and pheomelanin bind with high affinity (−8.7 and −7.7 kcal/mol, respectively) in active site of pink chromoprotein remarkably close to the prosthetic group and the cofactor. Eumelanin interacts with the residues Leu3B, Thr4B, Leu5B, Ser6B, Pro7B, Leu8B, Ala206B, Lys207B, Val210B, Lys211B, Tyr81A, Tyr85A, Glu195A, Gly198A, Glu199A, Ile201A, Ala202A, Asn203A, Trp205A, and Ala206A of chromoprotein (Figure 11B). The interaction of eumelanin is with the residues Ser6 and Pro7, the cofactor binding site, where eumelanin binds by weak hydrogen bonds. Whereas pheomelanin interacts with the residues Leu5A, Ser6A, Pro7A, Leu8A, Lys207A, Val210A, Lys211, Ala214, Gln215A, Tyr85B, Ile192B, Ala193B, Ser194B, Glu195B, Gly198B, Glu199B, Ala202B, Asn203, and Ala206 of protein (Figure 11C). Therefore, this result agrees with pheomelanin is tightly bound to proteins, indicating that in vivo it occurs as a chromoprotein.31-33

(A) Potential binding sites of pheomelanin (yellow), eumelanin (blue) 3H-indole-3-one (pink) binding sites, and Mg2+ ions (green) into pink chromoprotein of P. salmoneostramineus. (B) Aminoacid involved in the interaction with eumelanine. (C) Aminoacid involved in the interaction with pheomelanine.
Discussion
Although chromoproteins have been extensively studied in many marine organisms such as jellyfish and corals,34-36 little is known about these proteins in fungi. Oyster mushroom is an edible white-rot fungus and is classified into Pleurotus species comprising about 40 species. 37 Some kinds of oyster mushroom contain colored fruit bodies, and one of which is yellow (Pleurotus cornucopiae), 38 pink (PsPCP), 8 or white (Pleurotus florida). 39 Shibata et al 8 reported that the pink color of PsPCP was due to a chromoprotein, which plays a photosynthetic function. Later, Takekuma et al 5 were the first to isolate and report PsPCP, which has a maximum absorption spectrum at 496 nm. This protein contains a pigment molecule, 3H-indol-3-one, which occupies a portion of the active site and is thought to include 3 different metals: Zn, Fe, and Cu. In addition, Shibata et al 8 earlier completed crystallization and preliminary crystallographic research on PsPCP; however, the protein’s 3-dimensional structure was not reported. Finally, Shirasaka et al 4 provided thorough information on PsPCP purity, stability, and molecular mass. PsPCP has a molecular mass of approximately 24.5 kDa and is stable against heating at 28°C for 30 min and at pH values ranging from 4 to 10. The above information agrees with our bioinformatics results, which indicate that PsPCP has an isoelectric point of 7.505, a net charge at pH 7.4 of 0.116, and a molecular weight of 23 712 Da.
However, the MIB server confirmed that different metal ions can bind to PsPCP. However, the magnesium ion presented a higher affinity than iron, zinc, or copper (Table 3). This result is very interesting because a recent study performed with P. djamor showed that supplementation with magnesium salts in the culture medium increased the antioxidant capacity of the fruiting bodies, polyphenols, amino acids, including
It is worth noting that none of the prior studies on PsPCP documented its oligomerization state. Using GalaxyHomomer, we found that PsPCP forms dimers and prefers the creation of cavities between subunits where the Mg2+, 3H-indol-3-one, and possible substrate (eumelanin and pheomelanin) bond to protein. Furthermore, this region of the protein contains a serine residue (Ser6) that can be glycosylated. These findings are consistent with those of Korunova et al 42 who reported that Rhizostoma pulmo has an extracellular chromoprotein named rpulFKz1 contains an N-glycosylation site and produces oligomers (mainly dimers). Considering the similarity of biochemical properties between PsPCP and rpulFKz1, one can conclude that PsPCP is a chromoprotein relatively rare.
The amino acid residues involved in the interaction of the prosthetic group 3H-indol-3-one were identified using molecular modeling using CB-Dock2 server. The active site of PsPCP is formed by the residues Lue5, Ser6, Pro7, Lue8, Lys211, Ala214, and Gln215 in each subunit (Figure 6). In addition, some of these residues of amino acids also participate in the binding of eumelanin and pheomelanin (Figure 11). It is important to mention that the non-polar aliphatic amino acids are important in the formation of a non-aqueous environment, aiding the polar residues in substrate binding and enzymatic catalysis.43,44 However, crystallographic studies are required to corroborate the bioinformatic information and to be able to identify the amino acid residues that are critical for the binding of the 3H-indole-3-one.
We tried to predict the type of photoreceptor based on its possible subcellular location. Our results demonstrate that PsPCP could be a cytoplasmic and nuclear blue light photoreceptor. Accordingly, recombinant PsPCP was expressed as the soluble form in Escherichia coli, which suggest that PsPCP is a potential cytoplasmic protein. 3 Light controls important morphological and physiological responses in fungi. These organisms use chromoproteins to perceive blue, green, red, far-red, and near-UV light. These proteins present different subcellular localization. For example, The opsin photoreceptor for green light is a transmembrane protein, 45 whereas that the red light receptor phytochrome is found in the nucleus, mitochondria, and cytoplasm and is linked to other signaling proteins, 46 whereas the blue light photoreceptors reside in the nucleus, directly regulating the transcription of light-dependent genes. 47
To confirm whether PsPCP is a photoreceptor, we cultivated the fungus in the presence and absence of blue light and observed that the red hue did not change. This result does not agree with the real-time polymerase chain reaction (PCR) analysis performed by Fukuta et al 3 who observed a strong expression of PsPCP in the primordium formation stage of the life cycle of the fungus; however, its expression decreased with the maturation of the fruit body. It is interesting to note that the permanence of pink in carpophores is a still inconclusive part of fungus biology, which has generated opposing opinions in the published works, whereas Cedano et al 48 suggested that the loss of pink color in cultivated fruitings of a Mexican strain of Pleurotus ostreatorroseus was due to a highlight intensity in the growing areas. However, other authors did not find a correlation between the pink color variations of the basidiomes and the intensity of light used during their development. 49 This is probably due to the fact that the permanence of the color depends, both on the genetic characteristics of the parents, as well as on the cultural and environmental conditions in which the spores develop, similar to what has been reported for other species of the genus.50,51
Taken together, the data presented here suggest a complex and potentially novel role of PsPCP in melanin synthesis. Melanins are produced through the oxidation and polymerization of phenolic/indolic precursors; notably, these polymers are characterized by a strong negative charge, high molecular weight, and hydrophobic nature. Melanin contributes to the ability of fungi to survive in harsh environments. In addition, it plays a role in fungal pathogenesis. Melanin is an amorphous polymer that is produced by one of two synthetic pathways. Microbial melanins are heterologous, with 4 main types classified according to their chemical structures and biosynthetic pathways: eumelanin, pheomelanin, pyomelanin, and allomelanin.
52
Fungi may synthesize melanin from endogenous substrate via a 1,8-dihydroxynaphthalene (DHN) intermediate. Alternatively, some fungi produce melanin from
In the
Conclusions
Using free computer servers, it was possible to identify the potential binding sites of 3H-indole-3-one in the PsPCP apoprotein. The prosthetic group remains bound to the protein by hydrophobic interactions with Leu5, Leu8, Lys211, Ala214, and Gln215. In addition, the functional protein is possibly a dimer that can be glycated in Ser6 and binds to the Mg2+ ion in the same cavity through the residues of Ser6 and Pro7. Moreover, the chromoprotein is constitutive expression and is not dependent on blue light, so a putative function as a photoreceptor is ruled out. In addition, molecular docking studies showed that Leu5, Ser6, Pro7, and Leu8 are key residues because they contribute to the good binding affinity between prosthetic group, cofactor, melanin, and PsPCP. Therefore, chromoprotein is a melanosomal protein, and it perhaps has a critical role in melanogenesis and the melanin polymerization. However, additional experimental studies are required to evaluate the enzymatic activity of chromoprotein and to corroborate these in silico results.
Footnotes
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration Of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
All authors contributed significantly to the study: E.S.C. and M.A.V.S. design and led the study. M.A.V.S., E.K.V.G., I.A.C.G., and A.G.C. participated in the conduct and data analysis, and E.S.C., C.A.D., A.T.V., E.K.V.G., I.A.C.G., and M.A.V.S., participated in the final manuscript’s conduct, review, and correction. All authors read and approved the final manuscript.