
Abstract
5′-Ectonucleotidase (NT5E) catalyzes the conversion of adenosine monophosphate to adenosine and free phosphate. The role of this ectonucleotidase and its production of adenosine are linked with immune function, angiogenesis, and cancer. NT5E activity is typically assayed either by chromatographic quantification of substrates and products using high-performance liquid chromatography (HPLC) or by quantification of free phosphate using malachite green. These methods are not suitable for robust screening assays of NT5E activity. HPLC is not readily suitable for the rapid and efficient assay of multiple samples and malachite green is highly sensitive to the phosphate-containing buffers common in various media and sample buffers. Here the development and validation of a novel high-throughput ectonucleotidase screening assay are described, which makes use of a luciferase-based assay reagent, the Promega CellTiter-Glo kit, to measure the catabolism of AMP by NT5E. This multiwell plate-based assay facilitates the screening of potential ectonucleotidase antagonists and is unaffected by the presence of contaminating phosphate molecules present in screening samples.
Introduction
Adenosine monophosphate (AMP) is converted to adenosine (Ado) and free phosphate by the cell membrane–anchored ectonucleotidase NT5E. The role of NT5E (also known as CD73) and its production of adenosine are linked with immune function,1,2 angiogenesis, and tumorigenesis. 3 NT5E enzyme activity is typically assayed by either chromatographic quantification of substrates and products (AMP and/or Ado) using high-performance liquid chromatography (HPLC) before and after incubation with the enzyme or by quantification of free phosphate using malachite green.
These methods are not suitable for robust screening assays of 5′-ectonucleotidase activity. HPLC is not readily suitable for the rapid and efficient assay of multiple samples, and the malachite green assay is highly sensitive to the phosphate-containing buffers common in medium and other sample preparations used in screening. Specifically, the assay measures NT5E metabolism of AMP indirectly through relief of AMP-mediated inhibition of adenosine triphosphate (ATP) detection in a luciferase-based system. Although an indirect assay of NT5E activity, the ease of use and ready availability of instrumentation (standard luminescence plate readers) used for the assay described suggest an alternative to direct assay methods, which are less efficient or require specialized instrumentation. The present report describes a robust, reproducible multiwell plate-based assay for antagonists of NT5E that is sensitive, efficient, and impervious to the presence of contaminating phosphate molecules in screening samples.
Materials and Methods
Buffers and reagents
Tris magnesium (TM) buffer was prepared in water containing 25 mM Tris (Sigma Chemicals, St. Louis, MO) and 5 mM MgCl2 (Sigma Chemicals) as suggested by the manufacturers of recombinant human NT5E (R&D Systems, Minneapolis, MN). 5′-Ectonucleotidase assay buffer was prepared by adding ATP (Invitrogen, Carlsbad, CA) and adenosine monophosphate (AMP; Sigma Chemicals) to TM buffer for a final concentration of 100 µM and 300 µM, respectively. The CellTiter-Glo luciferase reagent from the CellTiter-Glo Luminescent Cell Viability Assay was used for measuring ATP and purchased from Promega Corporation (Madison, WI). α,β-Methylene adenosine-5′-diphosphate (APCP, Sigma Chemicals) was dissolved in TM buffer.
Assay conditions
Enzyme assays were carried out in 96-well multiwell plates in a final volume of assay buffer. Assay mixtures were incubated at 37 °C for the indicated incubation periods. The optimized assay buffer conditions included 300 ng/mL NT5E in TM buffer.
Antibody isolation using phage display technology
Phage display selections were performed to isolate scFv antibodies able to bind to MDA-MB-231 breast carcinoma cells. A combination of two large naive human scFv phage display libraries was used for antibody isolation as described previously. 4 The phage libraries and an aliquot of MDA-MB-231 cells were individually blocked, and blocked phage were added to the cells and incubated for 1 h at room temperature on a rotator before centrifugation at 1200 rpm for 1 min to pellet cells. The supernatant was removed and replaced with phosphate- buffered saline (PBS) to wash unbound phage from the cells. The centrifugation and washing steps were repeated a further 6 times before the addition of 500 µL elution buffer (triethanolamine 100 mM) and incubation at room temperature for 3 min, prior to adding 500 µL Tris pH 8.0. Infection of eluted phage into Escherichia coli cells and propagation were performed as described previously. 4 The scFv clones were converted directly into a human IgG format without affinity optimization. The target binding protein of the antibody used in these studies was determined to be 5′-ectonucleotidase using immunoprecipitation of MDA-MB-231 lysates followed by mass spectroscopy (Steven Rust, unpublished data).
Enzyme cloning and production
Human NT5E (NM_002526), minus the GPI linkage region, was obtained from placenta cDNA (BioChain, Newark, CA) using standard RT PCR and cloned into a mammalian expression vector (amino acids 1–552 cloned; GPI linkage region from amino acids 556–572 was not included). A 6× His tag was added at the C-terminus to aid with protein purification. The vector was used for transient expression of HEK293F cells (Invitrogen) using standard lipid transfection methods. Protein was purified from the supernatant.
Results and Discussion
The assay described makes use of a luciferase-based assay reagent, the Promega CellTiter-Glo (CTG) kit, to measure the conversion of AMP to Ado by 5′-ectonucleotidase. Briefly, the CTG kit measures ATP levels through the luciferase-linked emission of light accompanying catabolism of ATP to AMP and free diphosphate. The luciferase reaction of the CTG kit is inhibited by AMP and therefore yields little or no light in the presence of AMP. Addition of soluble recombinant NT5E to a mixture of ATP and AMP in the appropriate buffer results in the conversion of AMP and relief of AMP-mediated inhibition of the CTG reaction. The resultant increase in ATP detection (detected as light in the CTG assay) is directly proportional to NT5E enzyme activity.
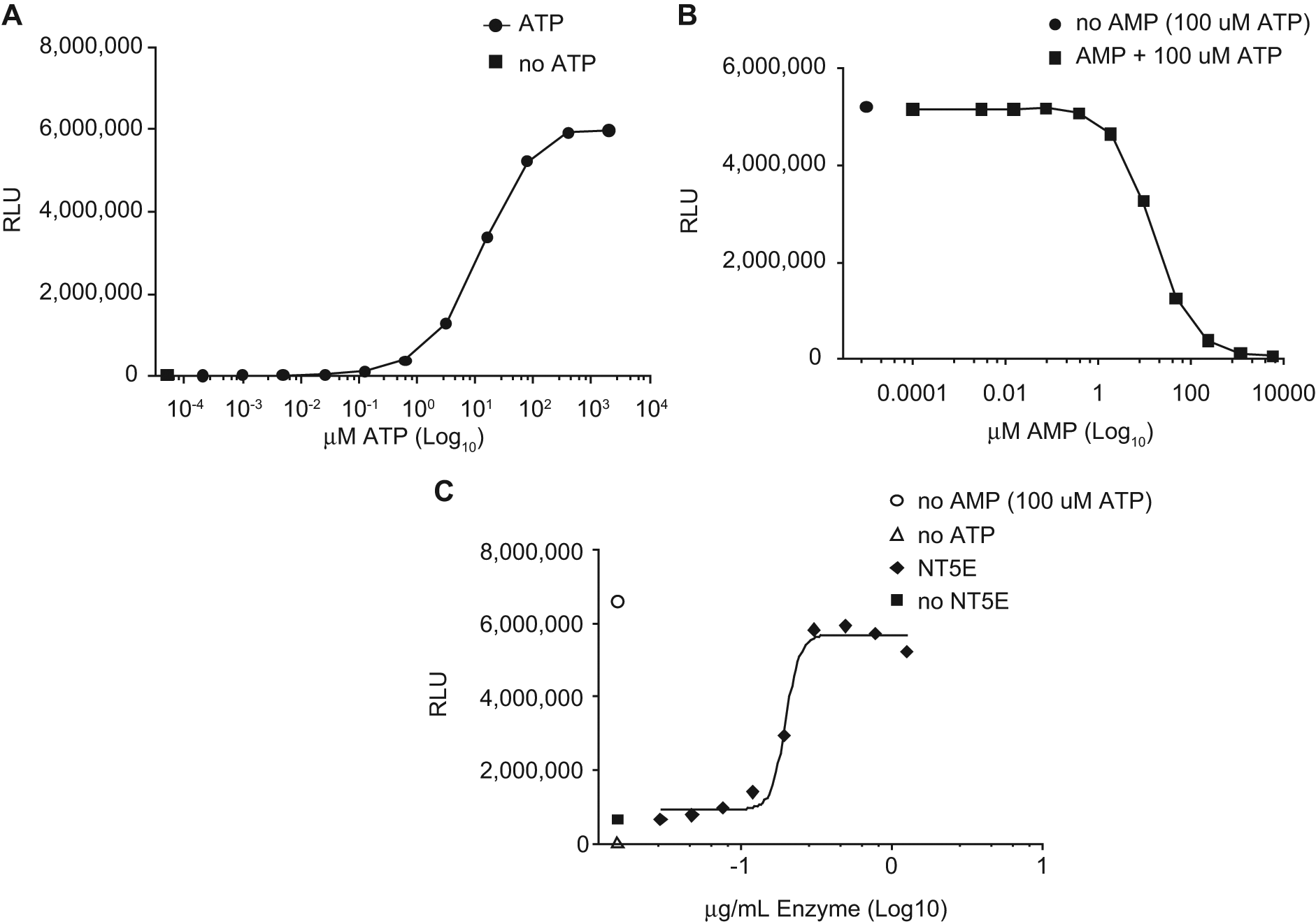
Serial dilutions of ATP were prepared in TM buffer and assayed in the CTG assay to determine what levels of ATP would show sensitivity to AMP inhibition ( Fig. 1A ). A near-maximal signal was observed at 100 µM ATP. The lowest levels of ATP yielded a signal comparable with TM buffer alone with no ATP. Similar results were observed in PBS (data not shown).
(
Serial dilutions of AMP were prepared in a solution of 100 µM ATP in TM buffer and assayed in the CTG assay to determine what levels of AMP will inhibit ATP quantification ( Fig. 1B ). Near-maximal inhibition was observed at approximately 300 µM AMP. The lowest and highest levels of AMP yielded a signal comparable to ATP or TM buffer alone, respectively. Similar results were observed in PBS (data not shown). The optimal molar ratio of AMP to ATP was determined to be approximately three to one under the assay conditions described. This relative excess of AMP appears to suppress luciferase activity without overwhelming the capacity of NT5E to metabolize AMP. It is currently unknown whether this ratio of AMP to ATP is physiologically relevant in healthy or diseased tissues, although ATP in solid tumors has been reported at similar levels. 5
Serial dilutions of recombinant human NT5E were prepared in assay buffer and incubated at 37 °C for 30 min to measure the ability of NT5E to relieve AMP-mediated inhibition of ATP detection by CTG ( Fig. 1C ). Half-maximal NT5E activity was observed at approximately 300 ng/mL. This level corresponds to ~5 nM rhNT5E (molecular weight [MW] = 63 kD). The highest and lowest levels of rhNT5E yielded ATP quantification levels comparable to AMP-free and enzyme-free preparations, respectively.
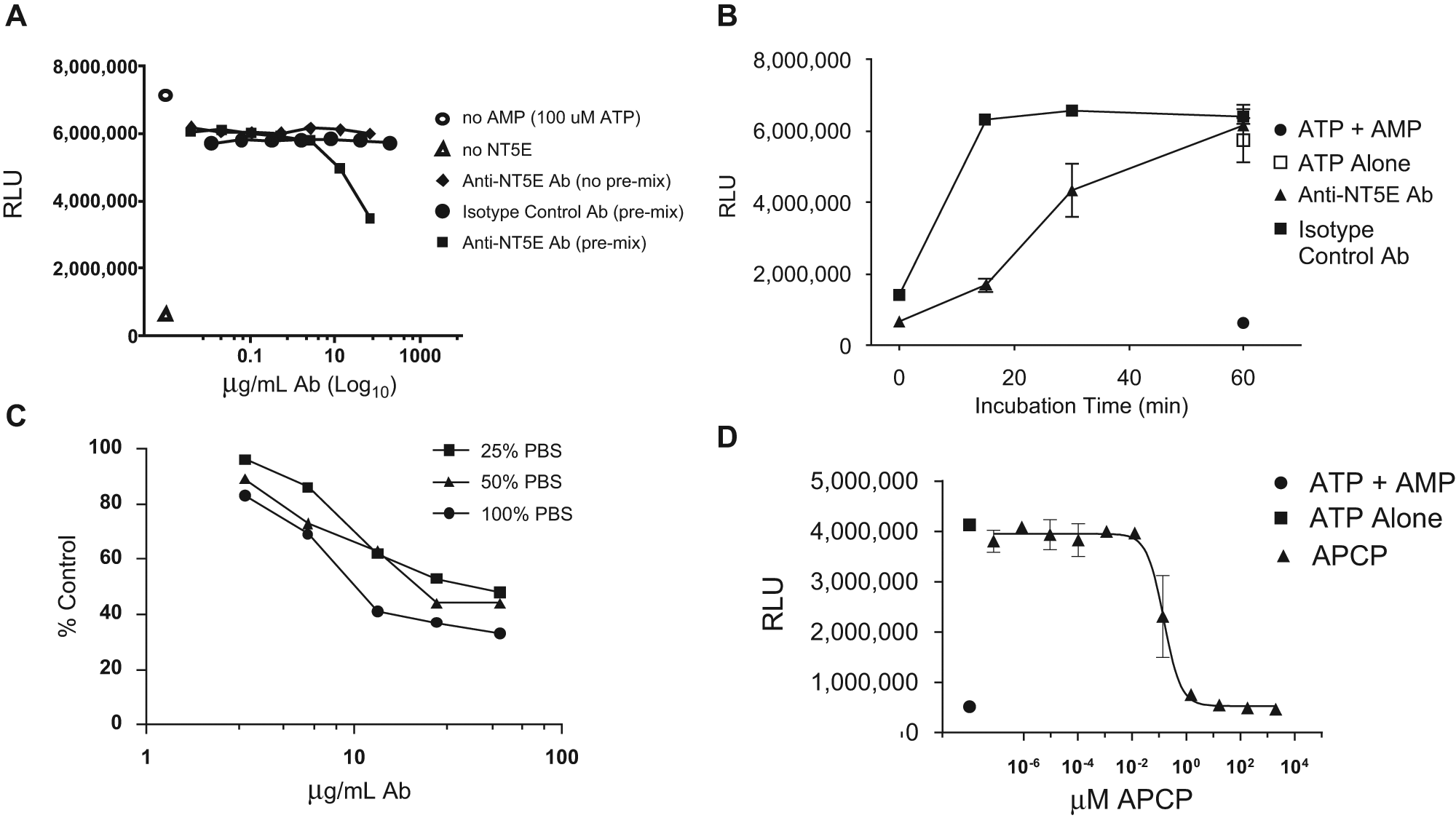
Serial dilutions of an inhibitory anti-NT5E antibody were prepared starting from 100 µg/mL in a solution of assay buffer containing 300 ng/mL rhNT5E. Dilutions were performed either with or without AMP to determine the impact of premixing antibody and enzyme prior to substrate (AMP) addition. An irrelevant isotype control antibody was similarly diluted to examine the specificity of antibody-mediated NT5E inhibition. Antibody/enzyme preparations were incubated for 30 min at 37 °C, and enzyme activity was quantified using CTG. NT5E activity was reduced to approximately half of that observed with AMP-free controls by the anti-NT5E antibody but not by the isotype control antibody. AMP reduced the CTG signal to levels similar to those observed with ATP-free and enzyme-free controls. The order of antibody addition was important for inhibition of NT5E as seen by the absence of enzyme inhibition when substrate (ATP) was added concomitantly with inhibitory antibody. This feature is not uncommon with inhibitory antibodies and is consistent with a potential role for antibody inhibition of substrate binding to the enzyme molecule. Complete enzyme inhibition by an antibody at the concentrations described may not result in 100% inhibition because the antibody is in dynamic equilibrium with the enzyme and, as such, is in competition with substrate AMP for binding to the enzyme. Even if an antibody is known to compete with substrate for binding to the active site of the ectonucleotidase, the antibody (at a MW of 150 kD, 150 µg/mL is only 1 µM) is not in sufficient excess over 200 µM AMP to expect full inhibition of enzyme activity.
To determine the optimal kinetics for assay antibody-mediated inhibition of NT5E activity, a fixed amount of antibody was incubated for various time periods at 37 °C in assay buffer containing 300 ng/mL rhNT5E. After 60 min of incubation, enzyme activity was similar to that observed with enzyme-free controls containing ATP alone. Enzyme incubated with isotype control antibody showed activity comparable with AMP-free controls at the first time point measured. The near-linear time-dependent nature of NT5E activity inhibition is consistent with simple enzyme assay kinetics within the range of substrate and assay conditions described. The modest (~50%) inhibition observed in the 30-min incubation used for order of addition experiments ( Fig. 2A ) is consistent with inhibition observed in the time course experiment shown ( Fig. 2B ). The enzyme inhibition observed after 15 min is suitable for high-throughput assays using automated liquid handling and signal readout instrumentation. A dependence on the order-of-addition for antibody-mediated enzyme inhibition similar to that shown in Figure 2A , following 30 min of incubation at 37 °C, was also observed following the 15-min incubation described in Figure 2B .
(
The performance of the ectonucleotidase inhibition assay in the presence of phosphate-containing buffers was examined. Figure 2C shows a dose-dependent inhibition of enzyme by the anti-CD73 antibody under the optimized assay conditions described, except using TM buffer prepared by substituting PBS (pH 7.5) for water at final concentrations of 25%, 50%, and 100%. The slight PBS-dependent impact on enzyme inhibition, even after normalizing data for antibody-free controls at each PBS concentration, was not significant. The overall degree of enzyme inhibition was comparable with that observed in TM buffer prepared in water. PBS-mediated effects may arise from salts present in PBS and as expected in samples used for screening.
The assay was tested using a nonhydrolyzable competitive inhibitor of NT5E, 6 APCP. As shown in Figure 2D , half-maximal inhibition was achieved at approximately 200 nM APCP. Enzyme activity was reduced to that of enzyme-free controls at approximately 20 µM APCP.
Assay performance was tested in the presence of crude samples representative of typical high-throughput phage antibody library discovery campaigns ( Fig. 3 ). These experiments were conducted to test the potential impact of contaminants such as phosphate, phage debris, or other contaminating sample components on assay parameters. Data for individual replicates are shown in Figure 3 , and quantitative summaries of assay characteristics are shown in Table 1 . The absolute assay signal window was over 495 000 relative light units (RLU) for an approximate twofold window between the anti-NT5E and isotype control antibodies. The assay showed intrasample coefficients of variability of 3% for the isotype control antibody and 4% for inhibitory anti-NT5E antibody. The thresholds for avoiding false-negative and false-positive hits were 92% and 19% of total enzyme inhibition, respectively, based on the observed signal window. A Z′ value of 0.650 indicated an excellent profile of signal window and variability as defined by Zhang et al. 7 Similar data were observed, including a larger window, for samples assayed in phosphate buffer (data not shown).
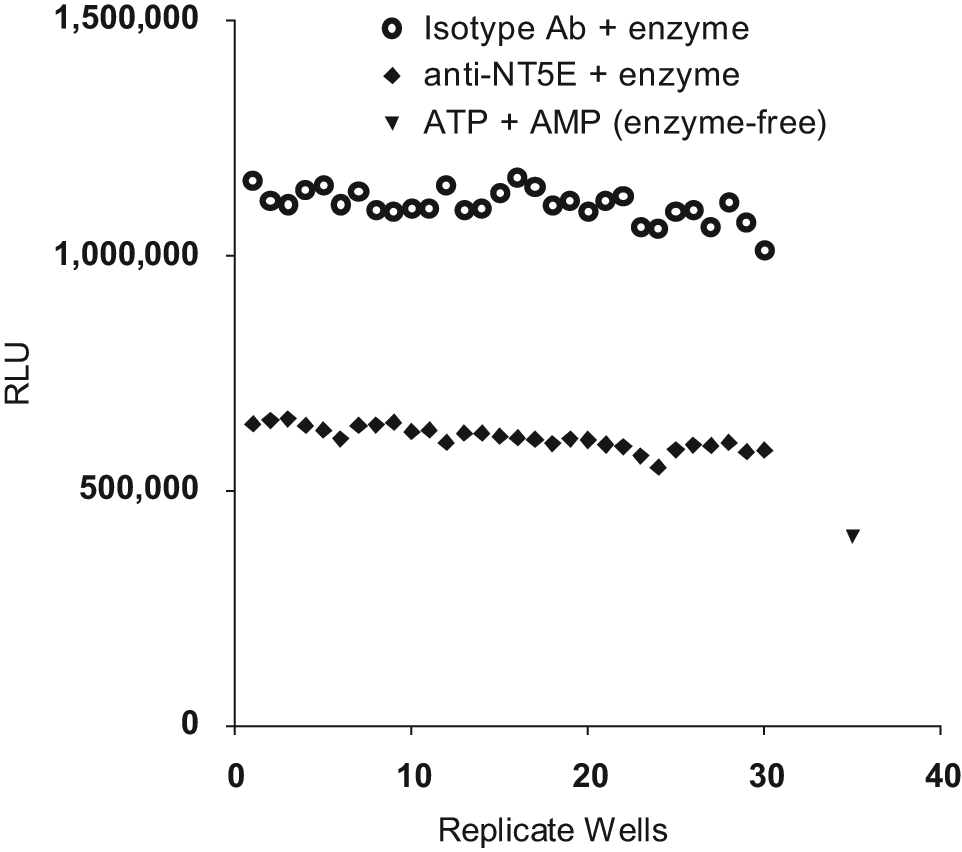
Impact of crude bacterial culture preparations upon assay performance. Thirty identical replicate wells containing an inhibitory anti-NT5E antibody (diamonds) or an isotype control antibody (circles) were incubated with a pooled collection of crude phage antibody library supernatants in assay buffer containing rhNT5E for 30 min. As a positive control for the inhibition of adenosine triphosphate (ATP) detection by adenosine monophosphate (AMP), a set of six wells was incubated with ATP and AMP in the absence of enzyme. The average value of these control wells is shown (triangle). ATP levels were then measured by luminescence (relative light units [RLU]) using CellTiter-Glo (CTG) reagent.
Characteristics of a NT5E Enzyme Activity Inhibition Assay
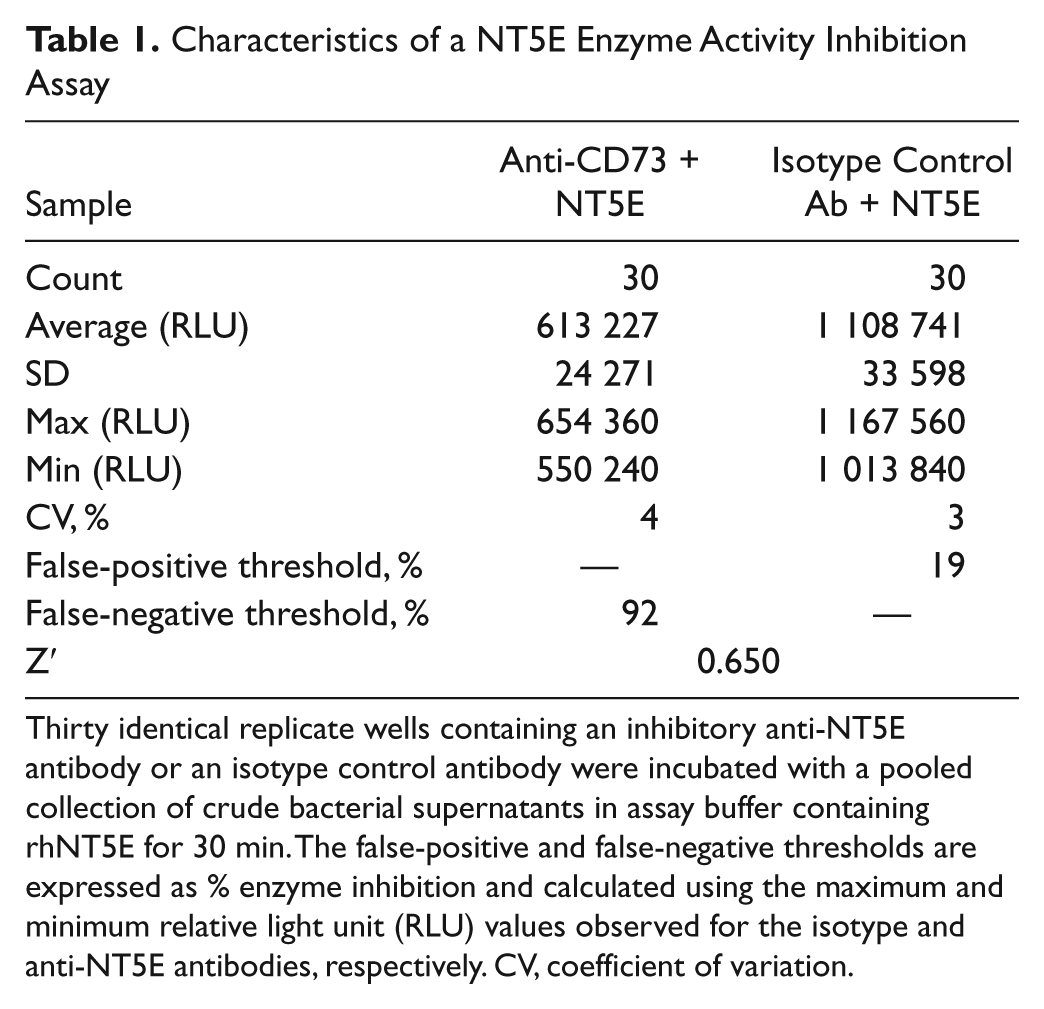
Thirty identical replicate wells containing an inhibitory anti-NT5E antibody or an isotype control antibody were incubated with a pooled collection of crude bacterial supernatants in assay buffer containing rhNT5E for 30 min. The false-positive and false-negative thresholds are expressed as % enzyme inhibition and calculated using the maximum and minimum relative light unit (RLU) values observed for the isotype and anti-NT5E antibodies, respectively. CV, coefficient of variation.
Specific enzyme inhibition by anti-NT5E antibody and not by an isotype control antibody within the context of a phage supernatant matrix is comparable with that seen after a similar incubation period using purified antibody. This suggests that the assay is sufficiently sensitive and specific as to detect other candidate inhibitory antibodies within crude preparations. It should be noted that the overall absolute ATP signal was reduced by crude supernatant preparations. The observation of similar results from recombinant mouse NT5E (data not shown) suggests that the assay can be used with other 5′-ectonucleotidase family members.
Finally, the assay showed characteristics suitable for high-throughput screening, including a reasonable signal window, relatively low coefficients of variation, and an excellent Z′ factor. A false-positive threshold was demonstrated when enzyme inhibition from all tested isotype control replicates (negative controls) reached a maximum of 19%. Similarly, a false-negative threshold was shown to be 92%—the minimum inhibition seen from all tested anti-NT5E (positive controls) antibodies.
It should be noted that these experiments were performed with the fully bivalent IgG antibody format and not the original monovalent library scFv format. Nevertheless, it is believed that the assay may have utility for various N5TE inhibitors, including small molecules or the bivalent scFv-Fc or full IgG expression systems used with increasing frequency for library screens. Furthermore, the vast molar excess of antibody compared with enzyme is expected to overcome simple mass-action effects.
Footnotes
Acknowledgements
The authors thank Dr. Peter Pavlik for critical reading of the manuscript and Qihui Huang for technical assistance.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.