
Abstract
In this research, we have established a high-throughput screening (HTS) platform based on the influenza A virus (IAV) vRNA promoter. Using this HTS platform, we selected 35 medicinal plants out of 83 examples of traditional Chinese medicine and found that 7 examples had not been reported. After examining many previous reports, we found that Vaccinium angustifolium Ait., Vitis vinifera L, and Cinnamomum cassia Presl had a common active compound, procyanidin, and then determined the anti-IAV effect of procyanidin and explored its mechanism of action. With a plaque inhibition assay and a time-of-addition experiment, we found that procyanidin could inhibit the IAV replication at several stages of the life cycle. In the Western blot and EGFP-LC3 localization assays, we found that procyanidin could inhibit the accumulation of LC3II and the dot-like aggregation of EGFP-LC3. In the RT-PCR and Western blot assays, we found procyanidin could inhibit the expression of Atg7, Atg5, and Atg12. Finally, by the bimolecular fluorescence complementation–fluorescence resonance energy transfer and co-immunoprecipitation assays, we found that procyanidin could inhibit the formation of the Atg5-Atg12/Atg16 heterotrimer and the dissociation of the beclin1/bcl2 heterodimer. In conclusion, we have established an HTS platform and identified procyanidin as a novel and promising anti-IAV agent.
Introduction
Influenza A virus (IAV) is a severe worldwide threat to human health, which can give rise to thousands of deaths, enormous economic losses, and social disturbances. Influenza viruses are RNA viruses belonging to the Orthomyxoviridae family (including influenza A, B, and C); the type A (i.e., IAV) has a negative single-stranded RNA with eight gene segments. By the antigenicities of hemagglutinin (HA) and neuraminidase (NA), it can be further classified into many subtypes; up to now, 16 serotypes of HA (H1–H16) and 9 of NA (N1–N9) have been found in the avian and mammalian hosts. Through the antigenic shift, a new virus strain can be generated by the exchange of HA and NA genes from different species, leading to a new pandemic. Although IAV consists of many subtypes, they have a common feature: The genome of IAV consists of eight RNA segments, with each segment flanked by the untranslated regions (UTRs) at the 5′ (13 nt) and 3′ (12 nt) termini, which are highly conserved among all IAV subtypes and among the eight segments themselves. These conserved sequences can anneal to form a bulged duplex structure that is essential for the transcription and replication of IAV, called the vRNA promoter. 1 Using this common feature, it is possible to establish a high-throughput screening (HTS) platform for all kinds of IAV subtypes.
On the basis of this common feature, we established a platform for high-throughput drug screening based on an IAV vRNA promoter luciferase reporter plasmid. Using this HTS platform, we analyzed 83 examples of traditional Chinese medicine and found that 7 medicinal plants had excellent inhibition on this platform that has not been reported. We picked out the blueberry (Vaccinium angustifolium Ait.), grape seed (Vitis vinifera L), and cinnamon bark (Cinnamomum cassia Presl) because, after examining many previous reports, we found that these plants had a common active compound, procyanidin. Procyanidin is a subclass of flavonoids that possesses antioxidant, anti-inflammatory, antibacterial, and antiarthritic activity. 2 It has been reported that procyanidin can inhibit the infections of hepatitis C virus (HCV), 3 severe acute respiratory syndrome coronavirus (SARS-CoV), 4 human immunodeficiency virus type 1 (HIV-1), 5 herpes simplex virus type 1 (HSV-1), 6 and Epstein-Barr virus (EBV), 7 but to our knowledge, there are no any reports about its ability to inhibit the infection of IAV. Therefore, we decided to determine the anti-IAV effect of procyanidin and explored its mechanisms of action.
Autophagy has been reported to be involved in the replication of IAV, and inhibiting autophagy can inhibit IAV replication. 8 IAV infection impedes the autophagy signal pathway by (1) producing many reactive oxygen species (ROS) via NOX2 NADPH oxidases, 9 which damage Atg4 and lead to the accumulation of LC3II (a light chain of the microtubule-associated protein), which is a mammalian Atg8 homologue 10 ; (2) binding to beclin1 by the IAV M2 protein, thus impeding the fusion of autophagosomes and lysosome 11 ; and (3) upregulating the expression of Atg7, Atg5, and Atg12. All of these lead to the accumulation of autophagosomes and prevent the IAV component from degradation. In addition, autophagy also plays an important role in the induction of the cytokine storm, which has been thought to be a main cause of death in IAV-infected patients, 12 so we decided to focus on the inhibition of procyanidin on IAV-induced autophagy.
Materials and Methods
Plant Extraction and Compound
Eighty-three examples of medicinal plants were collected from the Yulin medicine market (Guangxi Province, China). Each specimen was deposited in the Herbarium of Medical College, Shantou University (Shantou, China). The extracts from these medicinal plants were obtained according to the Chinese Pharmacopoeia (Chinese Pharmacopoeia Committee Publishing House of People’s Health, 2000). Procyanidin (purity >98%) was purchased from the National Institutes for Food and Drug Control (Beijing, China).
Viruses, Cells, and Cytotoxicity Assay
IAV subtypes A/PuertoRico/8/34 (H1N1), A/ShanTou/169/06 (H1N1), A/ShanTou/1233/06 (H1N1), A/ShanTou/602/06 (H3N2), A/ShanTou/364/05 (H3N2), A/Quail/HongKong/G1/97 (H9N2), A/Chicken/Guangdong/A1/03 (H9N2), and A/Chicken/GD/1/05 (H5N1) were used in this study. Virus stocks were prepared in the 10-day-old embryonated eggs or in a Madin-Darby canine kidney (MDCK) cell culture. The virus titer was determined by a plaque assay. 13 The cytotoxicity of all medicinal plant extracts and procyanidin was determined by an MTT method on A549 and MDCK cells, as previously reported. 14 The concentration of procyanidin required to lower cell viability by 50% (IC50) was calculated using Origin 8.0 software (OriginLab Corporation, Northampton, MA). Maximal concentrations with no cytotoxicity were used as the optimal concentrations in our tests.
Construction of Plasmids
To create an IAV vRNA promoter luciferase reporter plasmid, human pol-I promoter (GenBank: NT_167214.1) was cloned into a pGL3-basic vector and named the pPol-1 vector. A pGL3-Basic plasmid was used as the template for the amplification of the luciferase (luc) gene. The PCR products were cloned into the pPol-1 vector and named the pPolI-vluc plasmid, which contained a negative strand luciferase (–Luc) gene flanked by the 5′ and 3′ UTRs of the IAV A/PuertoRico/8/34 (H1N1) NP segment.
To construct the bimolecular fluorescence complementation–fluorescence resonance energy transfer (BiFC-FRET) plasmids, two segments of a red fluorescent protein (GenBank: HQ423140.1), corresponding to amino acids 1 to 159 and 160 to 262, were inserted into a pcDNA 3.0 plasmid and named pMC and pMN, respectively. Beclin 1 (NM_003766.3) and Atg5 (NM_004849.2) were inserted into the pMC plasmid and named pMC-beclin and pMC-Atg5, respectively. Bcl2 (NM_000633.2) and Atg12 (NM_ 004707.3) were inserted into the pMN plasmid and named pMN-bcl2 and pMN-Atg12, respectively. Atg16L1 (NM_030803.6) was inserted into a pEGFP-C1 plasmid and named pEGFP-Atg16.
To construct a pEGFP-LC3 plasmid, human LC3B gene (NM_022818.4) was inserted into a pEGFP-C1 plasmid. All constructs were verified by DNA sequencing.
HTS Assay
A549 cells were seeded into a 96-well microplate and transfected with the pPolI-vluc plasmid using the Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA). A pRL-CMV plasmid was used as an internal control to normalize the transfection efficiency. After a 6-h incubation at 37 °C, the cells were washed with phosphate-buffered saline (PBS), and the virus was introduced. After infection, the cells were grown in the virus growth medium (minimum essential medium [MEM]; 0.5 µg/mL tosylsulfonyl phenylalanyl chloromethyl ketone [TPCK]–trypsin [Sigma, St. Louis, MO], 0.125% [wt/vol] bovine serum albumin) containing the corresponding drugs (the plant extracts and procyanidin). The incubation time and multiplicity of infection (MOI) are shown in the following corresponding text. In the blank group, the cells were not treated with the virus and drug. The Luc activity was determined with the Luciferase Reporter Assay Kit (BD Biosciences Clontech, Franklin Lakes, NJ) using the GloMax-Multi+ Detection System Base Instrument with shaking (Promega, Madison, WI). The data from three independent experiments, each performed in triplicate, were collected and analyzed. All experiments with IAV were performed in a biosafety level 3 containment, including enhancements required by the U.S. Department of Agriculture and the Select Agents program. Moreover, the Z′ factor, which is a statistical parameter to quantify the suitability of a particular assay for use in a full-scale, high-throughput screen, was calculated according to a previous report. 15
Plaque Inhibition Assay
MDCK cells were seeded into six-well plates for 24 h. Before the experiment, the viruses were pretreated with a series of mediums containing procyanidin (3.13, 6.25, 12.5, and 25 µg/mL) for 2 h and then washed with PBS three times. The pretreated viruses were added and adsorbed for 1 h (MOI = 0.001) and washed with PBS three times, and the cells were cultivated in a series of mediums containing procyanidin (3.13, 6.25, 12.5, and 25 µg/mL) for 48 h. After frost-thawing one time, the supernatants were collected and the titers were determined using a plaque assay. 13 The concentration of procyanidin required to inhibit virus titer by 50% (50% effective concentrations [EC50]) was calculated using Origin 8.0 software. The antiviral index (AI) is represented as the ratio of IC50 to EC50.
Time-of-Addition Experiments
To detect which stage(s) the IAV life cycle was inhibited by procyanidin, we performed a time-of-addition experiment, as reported previously,16–18 which contained four tests: (1) Before infection, the virus was incubated with a medium containing procyanidin (25 µg/mL) for 3 h; (2) before infection, MDCK cells were incubated with a medium containing procyanidin (25 µg/mL) for 3 h; (3) procyanidin was added during viral adsorption for 1 h and removed by washing three times with PBS; and (4) procyanidin (25 µg/mL) was added at the beginning of incubation and removed by washing three times with PBS at the different time points after virus challenge. The incubation time was 24 h (MOI = 0.001). Other performances were same with the aforementioned plaque inhibition assay.
Dot-Like Aggregation of EGFP-LC3II onto Autophagosomes
After transfection with the pEGFP-LC3 plasmid for 6 h, A549 cells were infected with IAV (MOI = 2.0) and treated with drugs. At 8 h postinfection (PI), the cells were visualized under an invert fluorescence microscope (10 × 40), and the percentages of cells containing EGFP-LC3 dots to cells expressing enhanced green fluorescent protein (EGFP) were calculated in 10 fields chosen at random from three independent experiments.
Reverse-Transcription PCR (RT-PCR)
Extraction of RNA and RT-PCR reactions was performed according to the protocol of the TRIzol Reagent Kit and RT-PCR Kit (Invitrogen). The PCR product was electrophoresed in a 1% agarose gel and visualized on a UV-transilluminator.
Western Blotting
The anti-LC3B antibody, anti-beclin1 antibody, anti-Atg5 antibody, anti-Atg7 antibody, anti-Atg12 antibody, and anti-β-actin antibody were purchased from Cell Signaling Technology (Danvers, MA) and performed as previously reported. 19
BiFC-FRET Analysis
To assess the influence of procyanidin on the dissociation of the beclin-bcl2 heterodimer by the BiFC assay, A549 cells were seeded into 96-well plates for 24 h. After being cotransfected with the pMN-beclin and pMC-bcl2 plasmids for 6 h, the cells were infected with IAV at an MOI of 2 for 8 h. After 1 h at 4 °C, the fluorescence intensities were determined at 610 ± 10 nm after excitation at 587 ± 20 nm using a Tecan infinite M1000 premium Quad4 monochromator microplate reader (Tecan, Männedorf, Switzerland). The BiFC signals were shown as ΔFI610nm after deduction of the background fluorescence intensity.
Because the influence of procyanidin on the formation of the Atg5-Atg12 heterodimer will influence the formation of the Atg5-Atg12/Atg16 heterotrimer, we only quantified the fluorescence resonance energy transfer (FRET) signal between the Atg5-Atg12 heterodimer and Atg16. A549 cells were seeded onto a 96-well microplate for 24 h, and after being cotransfected with the pMN-Atg5, pMC-Atg12, and pEGFP-Atg16 plasmids for 6 h, the cells were infected with IAV at an MOI of 2, washed three times, and incubated for 8 h. After 1 h at 4 °C, the fluorescence intensities were determined by a Tecan infinite M1000 premium Quad4 monochromator microplate reader using the following settings: excitation at 488 nm with a 20-nm bandwidth, emission at 610 nm and 509 nm with a 20-nm bandwidth, a 100-µs lag time, and a 200-µs integration time. All FRET values were deducted against the background value, and the FRET efficiency (FRETe), expressed as the ratio of acceptor- and donor-emission intensities, was calculated according to the instructions of the Magellan Standard Soft (Tecan infinite M1000) and as previously reported 20 :
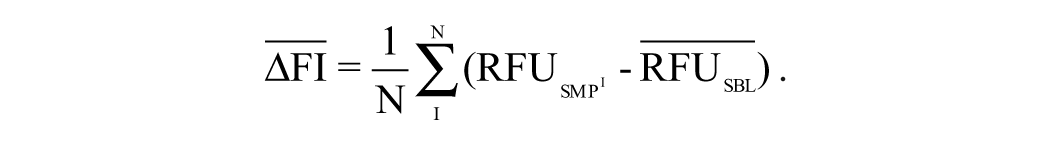
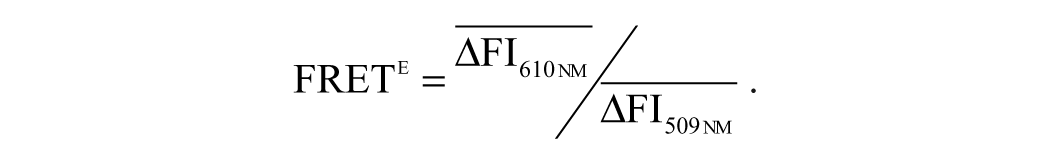
In these equations, RFUsmp is the fluorescence intensity of the sample group, and RFUsbl is the fluorescence intensity of the blank group.
To visualize the experimental results, A549 cells were plated onto glass coverslips in 12-well plates coated with poly-L-lysine for 24 h and then cotransfected with the aforementioned plasmids. After 6 h, the cells were infected with IAV at an MOI of 2, washed three times, and incubated for 8 h. After 1 h at 4 °C, the cells were fixed using 3% formaldehyde for 20 min and visualized using an upright fluorescence microscope (Nikon Eclipse 90i; Nikon, Tokyo, Japan). All images were acquired under the same setting. In this experiment, we could observe the results of the BiFC assay, but the FRET signal could not be observed because the human eye cannot distinguish the light of 610 nm and 509 nm at the same time.
Co-Immunoprecipitation
To detect the influence of the drugs on the interaction between the Atg5-Atg12 heterodimer and Atg16, as well as the interaction between beclin1 and bcl2 in physiological conditions, A549 cells were seeded into a 6-well plate for 24 h, then infected with IAV (MOI = 0.001) and treated with the drugs, and the cell lysate was collected at 24 h PI. The interactions were determined following the instrument of the Co-Immunoprecipitation Kit (#23600; Thermo Scientific, Waltham, MA). Normal rabbit IgG was used as a control.
Statistical Analysis
All data are expressed as mean ± SD. Statistical significance was determined using SPSS 13.0 software (SPSS, Inc., an IBM Company, Chicago, IL).
Results
Establishment of the IAV vRNA Promoter Luciferase Reporter HTS Platform and the Initial Screening by This Platform
The design of the IAV vRNA promoter luciferase reporter HTS platform is shown in Figure 1A . As shown in the figure, a negative strand of luciferase gene (–Luc) is flanked by the UTRs at the 5′ (13 nt) and 3′ (12 nt) termini, called the vRNA promoter, and then further flanked by the human pol-I promoter (rRNA promoter) and pol-I terminator. After transcription of this pPolI-vluc plasmid, a negative strand of luciferase RNA (–Luc RNA) flanked by the UTRs at the 5′ (13 nt) and 3′ (12 nt) termini would produce in cell nucleus transcribed by human rRNA promoter. After infection with IAV, the PA, PB1, PB2, and NP proteins of IAV would enter the cell nucleus and form vRNPs, which would transcribe the negative strand luciferase RNA (–Luc RNA) into positive strand luciferase RNA (+Luc RNA); +Luc RNA could be translated into luciferase. Determination of the activity of luciferase could detect the influence of testing drugs on IAV replication.
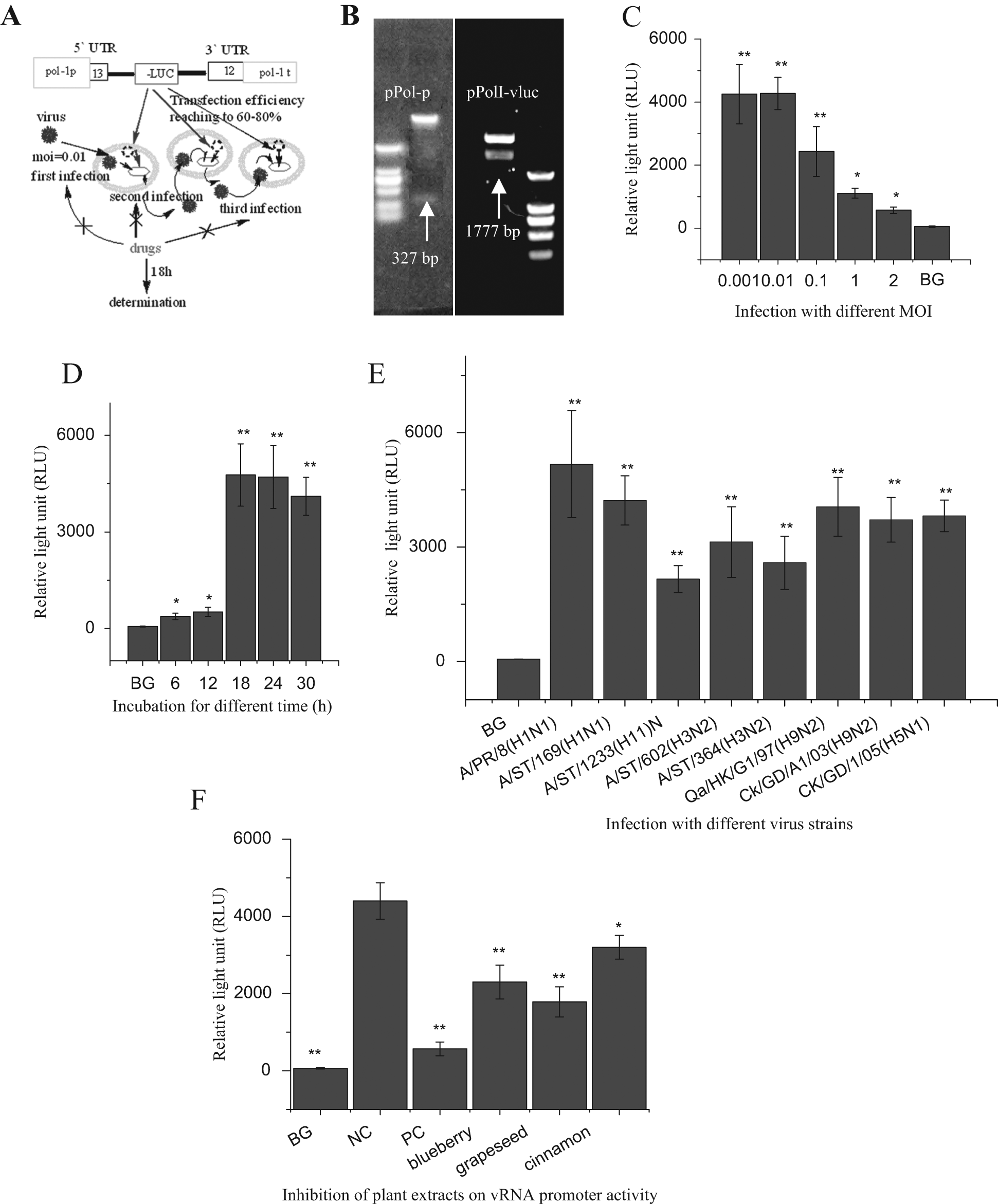
Construction and characterization of our influenza A virus (IAV) vRNA promoter luciferase reporter high-throughput screening (HTS) platform and the initial screening of natural products. (
Figure 1B shows the results of the double enzyme digestion of the pPol-1 vector and pPolI-vluc plasmid. In the establishment of our HTS platform, we first constructed a pPol-1 vector that contained a human pol-I promoter, and then we inserted a negative strand luciferase (–Luc) gene flanked by the 5′ and 3′ UTRs of the IAV A/PuertoRico/8/34 (H1N1) NP segment to construct a pPolI-vluc plasmid. The human pol-I promoter and the negative strand luciferase (–Luc) gene were 327 and 1777 bp, respectively. The results of the double enzyme digestion and DNA sequencing showed that these two constructs were correct.
To determine the optimal infectious dose, we transfected A549 cells with the pPolI-vluc plasmid and infected them with IAV A/ShanTou/169/06 (H1N1) at MOIs of 1, 0.1, 0.01, and 0.001. The Luc activity was assayed at 24 h PI, and the MOI of 0.01 was selected as the optimal infectious dose ( Fig. 1C ). To determine the optimal incubation time, we determined the luciferase activity at 6, 12, 18, 24, 30 h PI at an MOI of 0.01, and the optimal incubation time was selected as 18 h ( Fig. 1D ). To determine the universality of this platform, eight strains—A/PuertoRico/8/34 (H1N1), A/ShanTou/169/06 (H1N1), A/ShanTou/1233/06 (H1N1), A/ShanTou/602/06 (H3N2), A/ShanTou/364/05 (H3N2), A/Quail/HongKong/G1/97 (H9N2), A/Chicken/Guangdong/A1/03 (H9N2), and A/Chicken/GD/1/05 (H5N1)—were introduced, and all subtypes significantly upregulated the vRNA promoter activity ( Fig. 1E ). In these tests, the cells in the blank groups (BGs) were transfected with the pPolI-vluc plasmid but not infected with IAV. From these data, we can see that this reporter system could provide a platform for high-throughput drug screening.
Using this HTS platform, we investigated 83 examples of medicinal plants and found that 7 crude extracts had excellent activity that had not been reported. In
Figure 1F
, we show the results of the blueberry extract (V. angustifolium Ait.), grape seed extract (V. vinifera L.), and cinnamon bark (C. cassia Presl) extract. The data for the other 80 examples of medicinal plants are presented in
The Z′ factor is a statistical parameter to quantify the suitability of an HTS method. 15 The Z′ factor of this HTS platform was 0.57. According to the criterion stated in a previous report, 15 this HTS platform was statistically valid. In addition, to control the transfection efficiency, a pRL-CMV plasmid was used, and no significant difference was seen among all groups. Transfection efficiency can reach 60% to 80% according to the directions of the Lipofectamine 2000 reagent (Invitrogen).
Cytotoxicity and Inhibition of Procyanidin on the IAV Replication
The cytotoxicity of all extracts and procyanidin was determined on A549 cells and MDCK cells using an MTT method. Here we show the effect of procyanidin on MDCK cells. As shown in Figure 2A , procyanidin could significantly inhibit the activity of MDCK cells at concentrations of 50 to 125 µg/mL; at 25 µg/mL, procyanidin had no cytotoxicity. Y = −0.284 Ln(X) + 1.5598, R2 = 0.992, IC50 = 79.31 µg/mL. Here we selected 25 µg/mL as the optimal concentration.
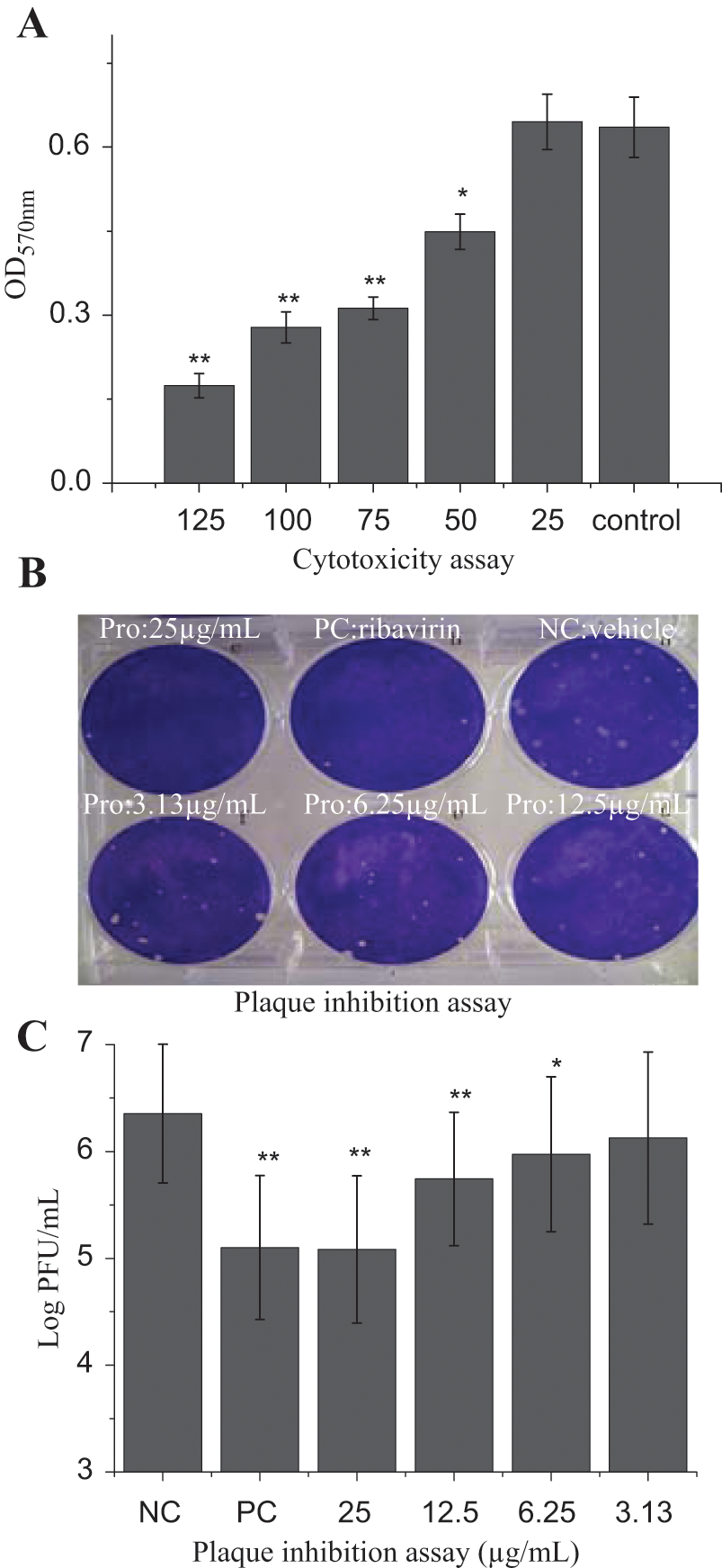
The cytotoxicity and plaque inhibition assay. (
Here we also directly assess the anti-IAV activity of procyanidin using a plaque inhibition assay on MDCK cells. As shown in Figure 2B , C , procyanidin significantly inhibited the viral replication at a range of 6.25 to 25 µg/mL in a concentration-dependent manner. Y = 0.049X + 6.3155, R2 = 0.9941, EC50 = 5.37 µg/mL, AI = IC50 / EC50 = 14.77.
Procyanidin Inhibits IAV Replication at Several Stages of the Virus Life Cycle
After determining the anti-IAV activity of procyanidin, we explored its mechanism of action. First we investigated which step(s) of the virus life cycle was affected by procyanidin using a time-of-addition experiment. As indicated in Figure 3A , procyanidin could significantly inhibit the replication of IAV, which meant that procyanidin could directly inactivate IAV. As indicated in Figure 3B , C , both of these experiments showed no significant difference, which meant that procyanidin had no influence on cells per se and on the adsorption of IAV. As shown in Figure 3D , procyanidin could significantly inhibit the IAV replication at 1 to 4 h PI. These results showed that procyanidin could inhibit the replication of IAV by several mechanisms of action.
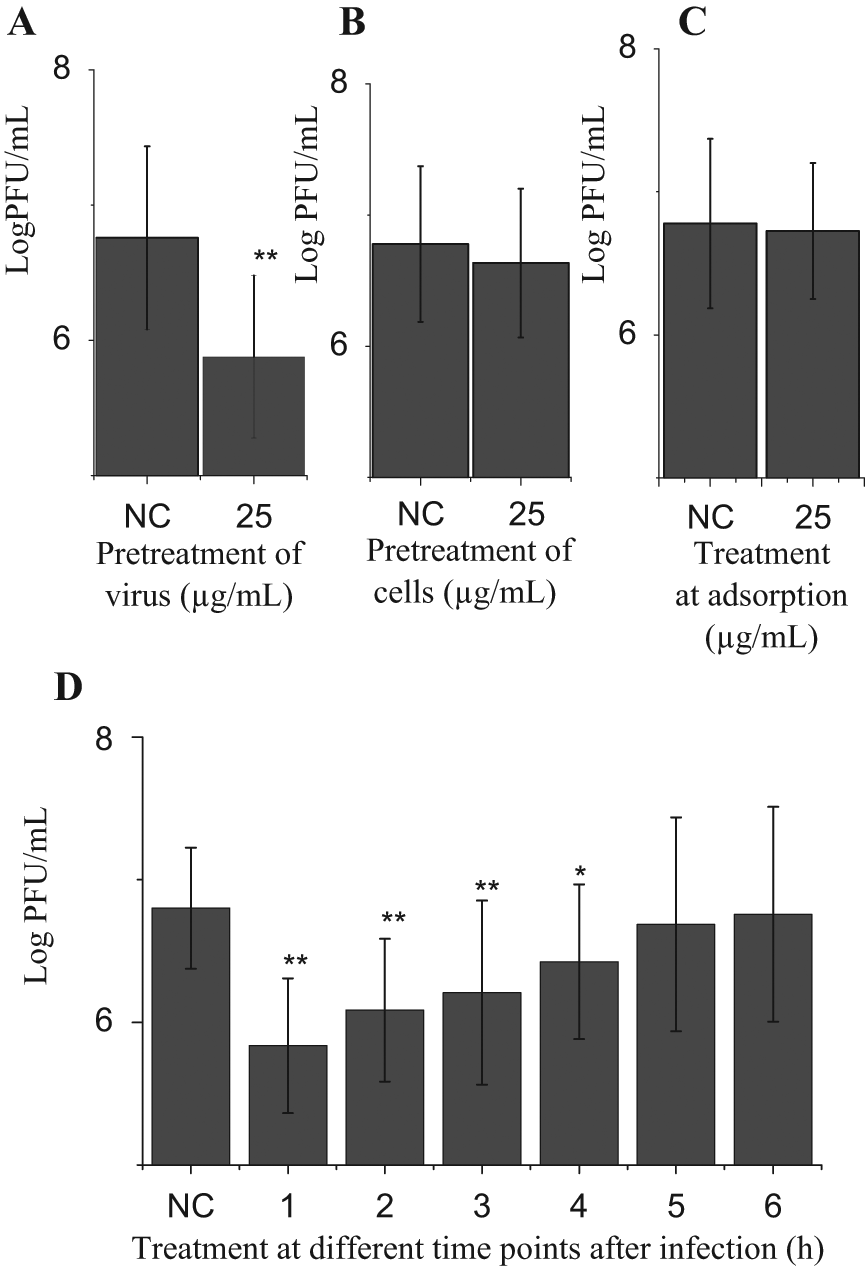
Time-of-addition experiments. (
Procyanidin Inhibits the Accumulation of Autophagosomes Induced by IAV
It has been reported that autophagy is a key process involved in IAV replication and may be responsible for the cytokine storm induced by highly pathogenic avian influenza H5N1 and H9N2 viruses. 12 The cytokine storm contributes to the high death rate in the flu pandemic. 22 In this section, we investigate the influence of procyanidin on IAV-induced autophagy.
Because the ratio of LC3II to β-actin was considered an accurate indicator for determining autophagy, 8 we investigated the levels of LC3II by Western blot. As shown in Figure 4A , after virus infection (negative control [NC]), the ratio of LC3II to β-actin significantly (p < 0.01) increased as compared with the blank group (BG); the ratio of LC3II to β-actin in the procyanidin (25 µg/mL) treatment group (Pro) significantly (p < 0.01) decreased as compared with that of the NC group.
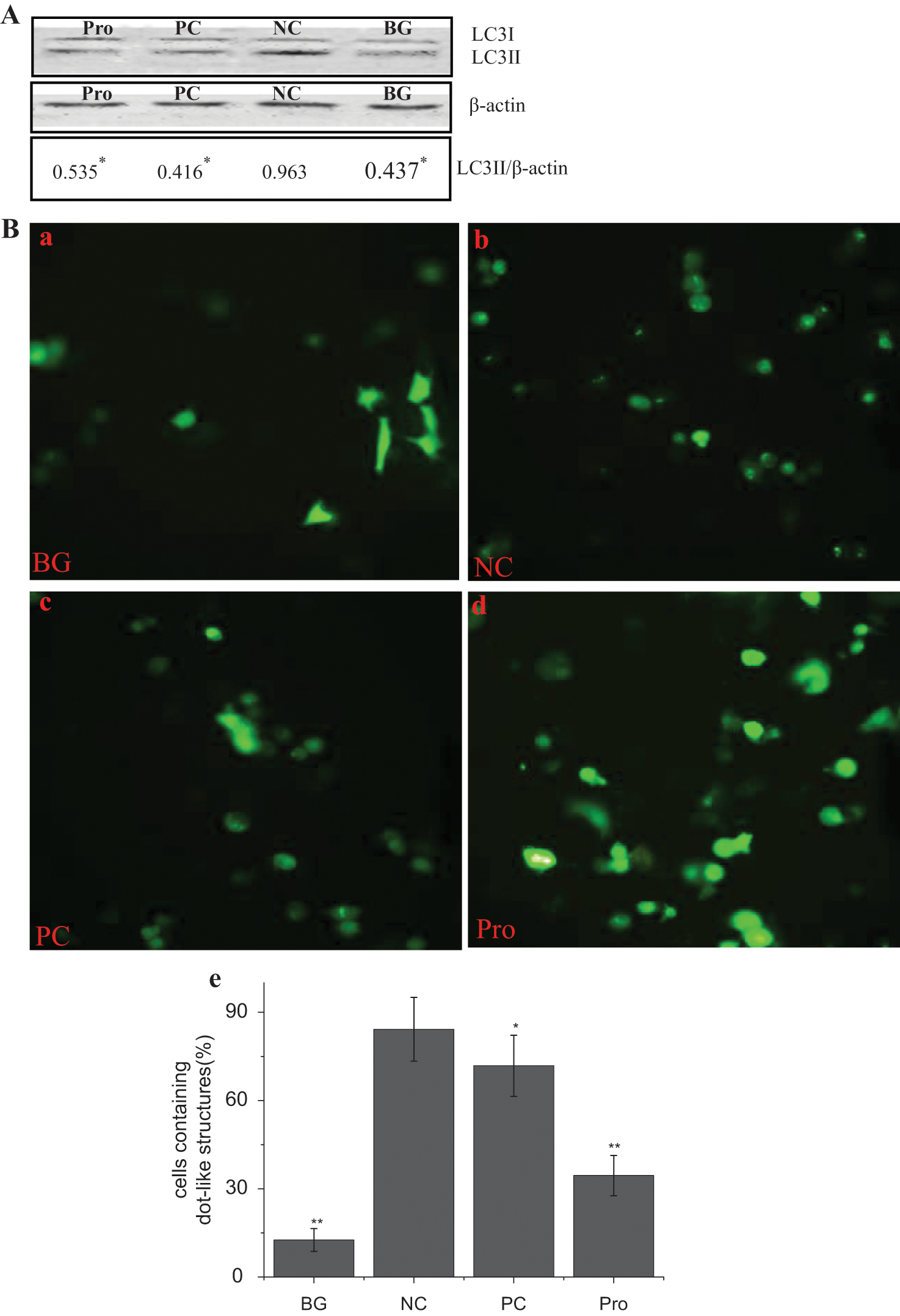
Procyanidin inhibits the conversion of LC3BI to II and the accumulation of autophagosomes induced by the influenza A virus (IAV). (
LC3II could accumulate on the autophagosome membrane,
23
and EGFP-LC3II fusion protein could form some dot-like structures on autophagosomes and were considered a specific marker of autophagosomes. We constructed a pEGFP-LC3 plasmid. As demonstrated in
Figure 4B
, the percentage of cells containing EGFP-LC3II dots to cells expressing EGFP after virus infection in the NC group (
Fig. 4B
,
Procyanidin Inhibits the Expression of Atg7, Atg5, and Atg12
It has been reported that the expression of beclin1, Atg16, and the dimer Atg5/Atg12 is upregulated after IAV infection 1 h. 24 Here we assayed the expression of Atg7, Atg5, Atg12, and beclin1 by RT-PCR ( Fig. 5A , B ) and by Western blot ( Fig. 5C , D ). As indicated in Figure 5A , B , the mRNA levels of Atg7, Atg5, Atg12, and beclin1 were significantly increased after IAV infection (NC group), as compared with the blank group (BG). Ribavirin (positive control [PC]) and procyanidin (Pro) could significantly inhibit the transcription of these genes.
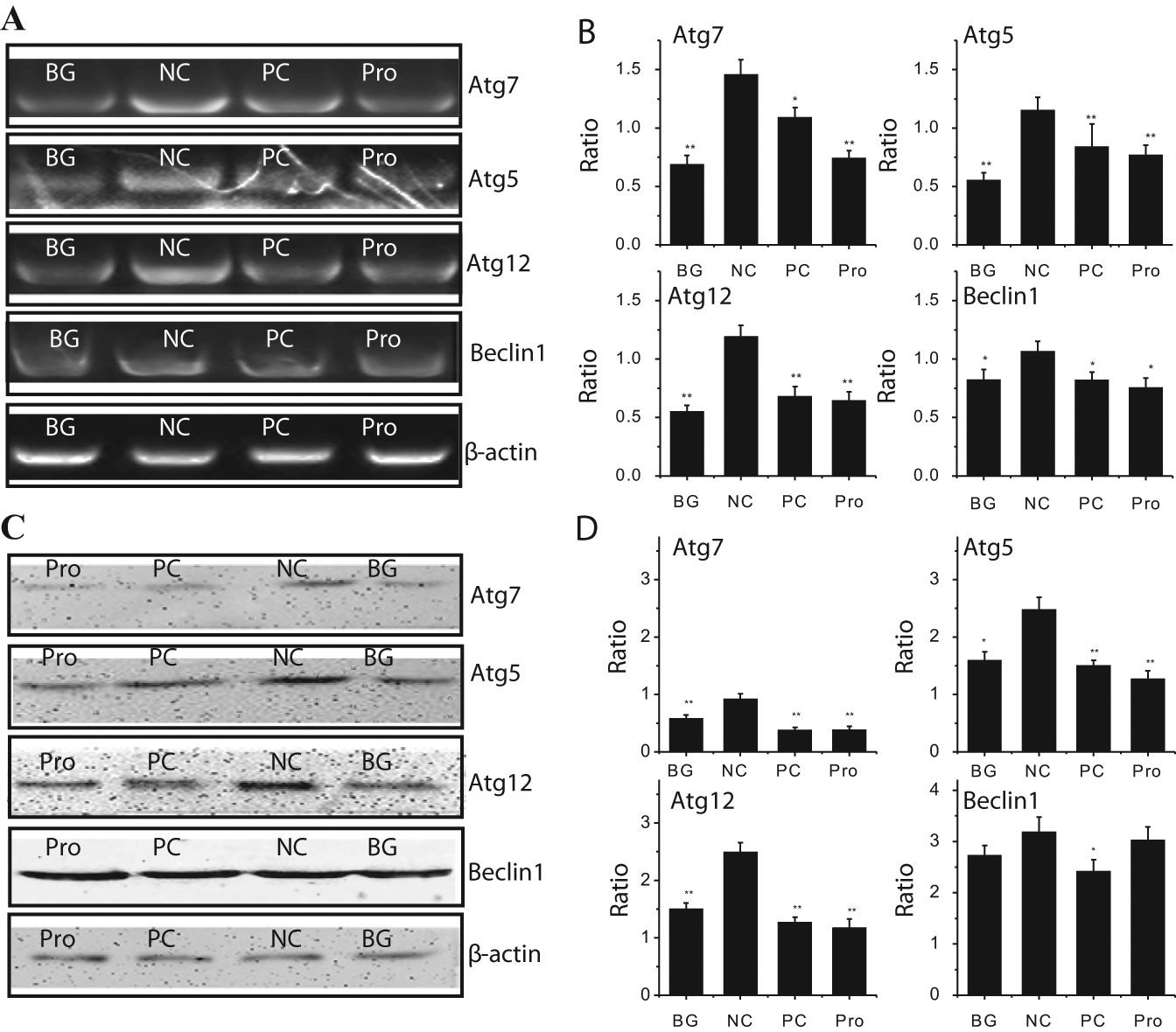
The inhibition of procyanidin on the expression of the autophagy genes. (
As indicated in Figure 5C , D , the protein levels of Atg7, Atg5, and Atg12 were also significantly increased after IAV infection (NC group) as compared with the blank group (BG group). Ribavirin (PC) and procyanidin (Pro) also could significantly decrease the levels of these proteins compared with the NC group. The levels of Atg7, Atg5, and Atg12 in ribavirin-treated and procyanidin-treated groups were even lower than that in the BG group, and between the ribavirin-treated and procyanidin-treated groups, the differences of these proteins were not significant. Moreover, the influence of ribavirin (PC) on beclin1 was significant, but the influence of procyanidin (Pro) was not significant.
Procyanidin Inhibits the Formation of the Atg5-Atg12/Atg16 Heterotrimer and the Dissociation of the Beclin1/bcl2 Heterodimer
The formation of the Atg5-Atg12/Atg16 heterotrimer and the dissociation of the beclin1/bcl2 heterodimer are two important events occurring in the autophagy signal pathway.
25
IAV is involved in these two events,
26
as shown in
The BiFC-FRET assay is a newly emerged technique and can be used to provide evidence of the ternary complex formation in living cells.
27
The BiFC technique is based on the principle of protein fragment complementation, in which two nonfluorescent fragments derived from a fluorescent protein are fused to a pair of interacting proteins. When the two proteins interact, the two nonfluorescent fragments are brought into proximity, and an intact fluorescent protein will be reconstituted. The FRET technique is based on the transfer of excitation energy between two fluorophores that are in close spatial proximity; this energy transfer results in a change in the fluorescence intensity of the two fluorophores. In our study, we used this BiFC-FRET technique to investigate the influence of procyanidin on the formation of the Atg5-Atg12/Atg16 heterotrimer and the dissociation of the beclin1/bcl2 heterodimer. The design of the BiFC-FRET assay is shown in
In our study, we first examined the influence of procyanidin on the interaction among Atg5, Atg12, and Atg16. As shown in Figure 6 , in the BG group ( Fig. 6A , Q ), the A549 cells still had the BiFC and FRET signals, which might be due to the autophagy of the cell per se. In the NC group ( Fig. 6D , Q ), the BiFC and FRET signals dramatically increased as compared with the BG group. Ribavirin and procyanidin could significantly downregulate the BiFC and FRET signals ( Fig. 6G , J , Q ) as compared with the NC group. This indicates that procyanidin can inhibit the formation of the Atg5-Atg12/Atg16 heterotrimer.
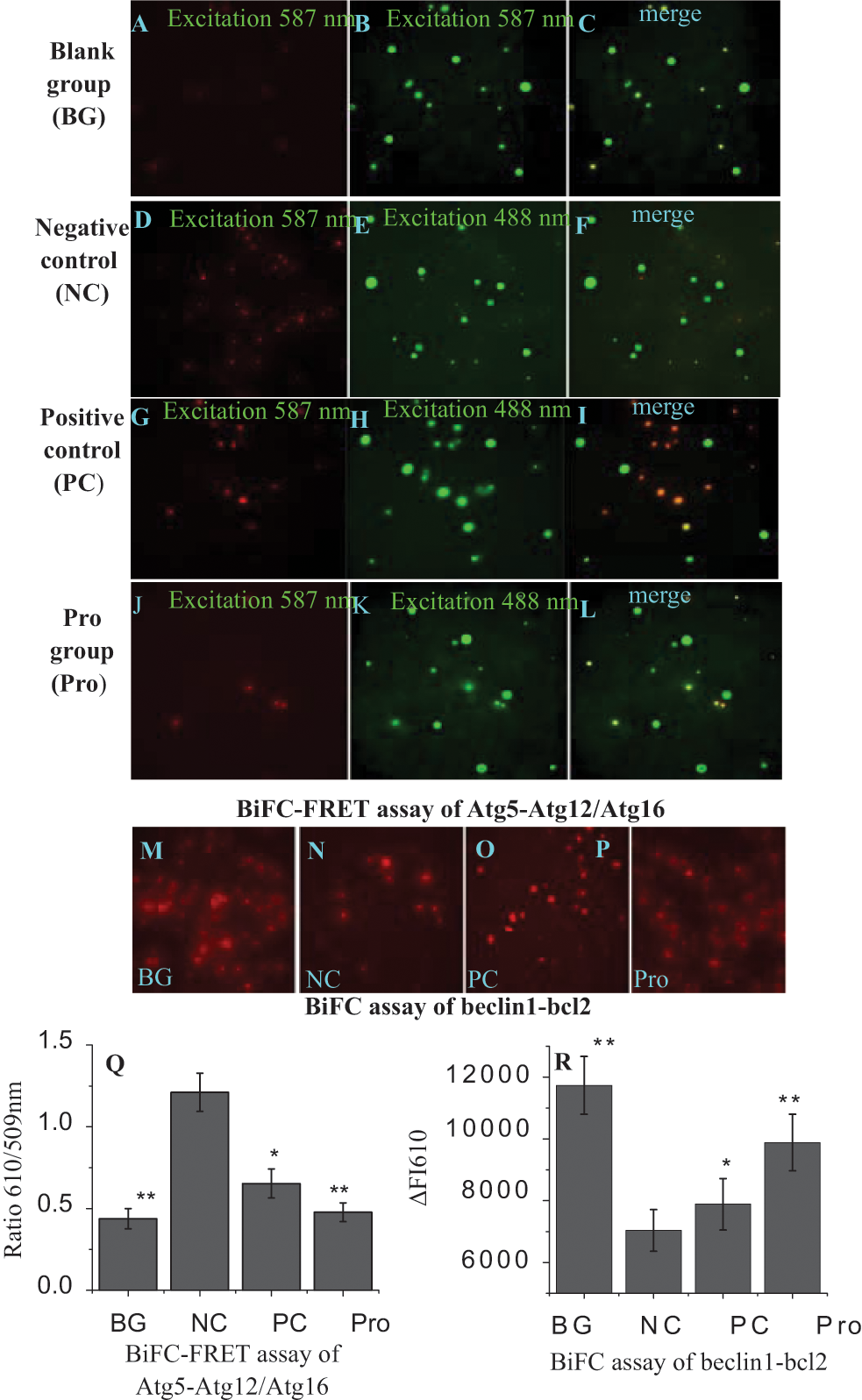
Procyanidin inhibits the formation of the Atg5-Atg12/Atg16 heterotrimer and the dissociation of the beclin1/bcl2 heterodimer. (
Then we examined the influence of procyanidin on the dissociation of the beclin1-bcl2 heterodimer. After infection with virus, the BiFC signal in the NC group ( Fig. 6N , R ) significantly (p < 0.05) decreased as compared with that in the BG group ( Fig. 6M , R ), whereas the BiFC signals in ribavirin-treated and procyanidin-treated groups significantly (p < 0.01) increased ( Fig. 6O , P , R ) as compared with the NC group. This indicates that ribavirin and procyanidin could promote the stability of the beclin1-bcl2 heterodimer.
To further confirm the influence of procyanidin on the interaction between the Atg5-Atg12 heterodimer and Atg16 in physiological conditions, we performed the immunoprecipitation experiments using anti-Atg5 antibody and then performed a Western blot assay using the anti-Atg16 antibody. We found that Atg5 could co-immunoprecipitate with Atg16; after infection with IAV, the conjugation of Atg5 and Atg16 was obviously increased, and ribavirin and procyanidin could inhibit these processes ( Fig. 7A ). Using the same method, we found that beclin1 could co-immunoprecipitate with bcl2; after infection with IAV, the conjugation of beclin1 and bcl2 was obviously decreased, but ribavirin and procyanidin could increase the level of the beclin1-bcl2 heterodimer.
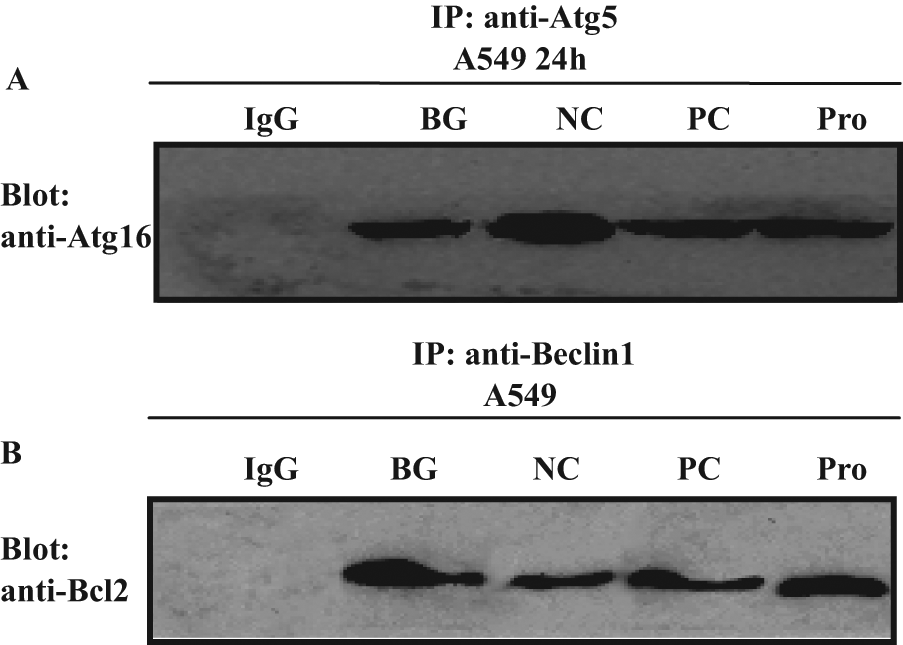
Co-immunoprecipitation assay of the Atg5-Atg12/Atg16 heterotrimer and the beclin1-bcl2 heterodimer. (
These experiments indicate that procyanidin could inhibit the formation of the Atg5-Atg12/Atg16 heterotrimer and promote the stability of the beclin1-bcl2 heterodimer.
Discussion
When confronted with a sudden outbreak of an IAV pandemic, no matter what subtype it is, an HTS platform that can screen the drugs effective against all types of IAV will be very useful. Our HTS platform has this character. In our HTS platform, we use the common promoter of all IAV subtypes, which is highly conserved among all types of IAV and among the eight segments themselves, so in theory, this HTS platform should have excellent universality, and in fact, we have found that eight strains of IAV, including four subtypes (H1N1, H3N2, H9N2, and H5N1), can transcribe this common vRNA promoter ( Fig. 1E ). Therefore, we deduce that the drugs screened by this HTS platform may have the ability to inhibit all subtypes of IAV, which will be very useful for the sudden outbreak of pandemics.
Moreover, in this HTS platform, the transfection efficiency can reach 60% to 80% (i.e., 60%−80% cells in a well of a 96-well plate can be transfected), and the MOI we used is 0.01—that is, in the first round of infection, only 1% of cells are infected, and a large number of cells are not infected but have been transfected with the pPolI-vLuc plasmid, so several rounds of infection will take place ( Fig. 1A ). The drugs that inhibit the replication of IAV at any stage of the IAV life cycle (e.g., entry, uncoating, biosynthesis, and assembly) will decrease the yield of progeny viruses and thus decrease the infectious dose of the virus at the next round of infection, so the total yield of luciferase will decrease. Therefore, this platform also provides a method for screening anti-IAV drugs, no matter which stage(s) of the virus life cycle they target.
Using this HTS platform, we screened 83 examples of Chinese herbal medicine and obtained 35 examples that could significantly inhibit IAV vRNA promoter transcription activity. After examining a lot of literature, we found at least 7 examples that had not been reported up to now (i.e., Peucedanum praeruptorum Dunn., Lantana camara L., Litsea cubeba [Lour.] Pers., Ginkgo biloba L., V. angustifolium Ait., V. vinifera L, and C. cassia Presl); many of the others had been recorded or reported in Chinese ancient or modernistic literature but many not in scientific journals. There is a common problem in the research of traditional Chinese medicine: The active ingredient and mechanism of action are not clear. As for the above 7 examples, we found that V. angustifolium Ait., V. vinifera L, and C. cassia Presl are rich in procyanidin, and many studies show that procyanidin is one of the major active ingredients of these herbal medicines, so we decided to research its effect and mechanism on anti-IAV infection.
Procyanidin is a very strong antioxidant. IAV infection will lead to the accumulation of autophagosomes by producing a lot of ROS, 9 so we speculate that the antioxidant of procyanidin may be related to its anti-IAV activity by inhibiting the accumulation of autophagosomes induced by IAV. In our study, the plaque inhibition assay showed that procyanidin could inhibit the replication of IAV. The Western blot and dot-like aggregation of EGFP-LC3II assays showed it could inhibit the accumulation of LC3II, which showed that procyanidin could reduce the autophagy induced by IAV. The RT-PCR and Western blot assays showed that procyanidin could inhibit the expression of the autophagic genes Atg7, Atg5, and Atg12. The BiFC-FRET and co-immunoprecipitation assays showed that procyanidin could inhibit the formation of the Atg5-Atg12/Atg16 heterotrimer and the dissociation of the beclin1/bcl2 heterodimer. So we think that one of the mechanisms of procyanidin on anti-IAV infection is related to its ability to inhibit the autophagic gene expression and the autophagic signal pathway. But it is only a part of the mechanism of its anti-IAV activity because our time-of-addition experiments have shown that procyanidin can inactivate IAV directly. Moreover, procyanidin also can inhibit the NF-kappaB signal pathway, 28 which is an important pathway needed by IAV replication.
Phytotherapy has a long history to cope with resistance against viruses and plays a fundamental role in traditional medicines in China, Korea, Japan, and India. 29 In 2009, the swine H1N1 IAV pandemic originated from Mexico and the United States, which prompted the Chinese government to release three editions of a document titled ‘‘Recommended Schemes for Pandemic Influenza A Diagnoses and Treatments.’’ 30 The third edition recommends the use of not only oseltamivir and zanamivir but also four anti-flu traditional Chinese medicine prescriptions. In fighting the pandemic, traditional Chinese medicines have played a significant role. But the mechanism of action of many traditional Chinese medicines is poorly understood, so our task and purpose are not only to select these drugs but also to rapidly categorize these drugs according to their targets of action and obtain the most optimal prescriptions.
Footnotes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China (30972766, 31170852, 81001322, 81001340, and 81172795) and Guangdong Natural Science Foundation (9451503102003499).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.