
Abstract
Objective: To examine the effects of the methylenetetrahydrofolate reductase (MTHFR) and methionine synthase reductase (MTRR) gene polymorphisms and their interactions with environmental factors on serum lipid levels. Methods: We investigated totally 340 patients with essential hypertension, from Dongzhi community, Anhui, China. High-throughput TaqMan allelic discrimination assay was used for the genotyping of MTHFR C677T (Ala222Val), MTHFR A1298C (Glu429Ala), MTRR A66G (Ile22Met), and MTRR His595Tyr. Results: Compared with the MTRR 66AA genotype carriers, the GG genotype carriers had lower serum total cholesterol (TC) levels (adjusted β ± standard error [SE]: −0.5 ± 0.2 mmol/L; P = .003) and low-density lipoprotein cholesterol (LDL-C) levels (adjusted β ± SE: −0.4 ± 0.2 mmol/L; P = .005). Their false discovery rate (FDR)-adjusted P values were 0.056 and 0.056, respectively. We further found that there was a statistically significant interaction between 677TT genotype and sex in their associations with LDL levels (P interaction = .020), and significant interaction between 677TT genotype and smoking on LDL levels (P interaction = .036). A similar pattern of interaction was found between 66GG and drinking on levels of TC (P interaction = .034) and LDL (P interaction = .020). However, there were no significant interactions observed after FDR adjustment. Conclusion: Both MTHFR and MTRR gene polymorphisms could be important genetic determinants of serum lipid levels in Chinese patients with hypertension. These findings need to be replicated in a larger sample.
Introduction
Increased homocysteine (Hcy) level is generally considered to be an independent risk factor for the development of cardiovascular disease (CVD). The links between hyperhomocysteinemia (HHcy) and elevated cardiovascular event risks have been established, 1 but the exact role of Hcy in CVD is unclear. Especially, whether Hcy affects lipid function remains underexplored.
In recent years, multiple lines of evidence showed that serum Hcy is positively correlated with total cholesterol (TC).
2
–5
In rats with experimentally induced HHcy, the amounts of plasma TC levels significantly increased, likely due to the high expression and activity of hepatic 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase, a rate-limiting enzyme in TC biosynthesis.
2
–4
Diet-induced HHcy caused elevated TC and triacylglycerol (triglyceride [TG]) in mouse liver
5
and correspondingly homocysteinemia (Hcy) reduced high-density lipoprotein cholesterol (HDL-C).
6
Apolipoprotein (apo)-A1 is the main apolipoprotein of HDL-C.
7
S-adenosyl-
Methylenetetrahydrofolate reductase (MTHFR) is a key enzyme in the Hcy metabolic pathway and reduces 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate, which provides the methyl group that converts Hcy to methionine. The gene is located on chromosome 1p36.3 with 2.2 kb length of complementary DNA (cDNA). C677T mutation in MTHFR gene is closely related to CVD risk. 10 The variant creates a thermolabile enzyme with 50% reduction in activity at 37°C and is associated with a 25% increase in Hcy concentrations. 11 Yilmaz et al 12 have shown by screening MTHFR C677T polymorphism in renal transplant patients that the C allele has a protective effect on blood lipid levels, while the T allele has a harmful effect. Compared with the CC genotype, participants with the T allele have significantly higher TC and low-density lipoprotein cholesterol (LDL-C) levels. Zhang et al showed that the MTHFR 677T carriers in Yao and Han people of Guangxi, China, have significantly higher TC and LDL-C levels. 13 As for the MTHFR A1298C polymorphism, there is no clear effect on Hcy level in plasma and no any report of its association with serum lipid level.
The roles of polymorphisms in other genes in the Hcy metabolic pathway have not yet been fully evaluated for associations with the risk of hyperlipidemia. Methionine synthase reductase (MTRR), in particular, plays a crucial role in restoring the active state of methionine synthase (MS) activity by reductive methylation of cob (II) alamin, using S-adenosylmethionine as methyl donor. It is another important enzyme in the metabolism of Hcy. The MTRR gene mutation may lead to differences in its enzyme expression and function, affecting the levels of plasma Hcy. Studies have shown that the 66AA genotype contributes to a moderate increase in Hcy levels, 14,15 with an approximate 4% increase in CVD risk, compared to 66GG homozygotes. 15 The MTRR helps to maintain the levels of intracellular methionine, folate, and nontoxic Hcy. The MTRR gene A66G causes elevated Hcy and is an important candidate gene for HHcy. 16 Ohnami et al 17 found that His595Tyr as a common missense single-nucleotide polymorphism (SNP) of the MTRR gene is a novel pancreatic cancer susceptibility factor. However, the correlation between the MTRR gene and blood lipids remains unclear.
Epidemiological studies 18 –20 showed that cigarette smoking and alcohol drinking were both risk factors for HHcy and associated with elevated Hcy concentrations. Long-term heavy smoking and alcohol drinking interfere with vitamin B6, vitamin B12, and folic acid metabolism, which will lead to plasma Hcy accumulation. Gender is also a key determinant of Hcy level. 18 We hypothesize that there might be interactions between Hcy metabolic-related genetic variants and gender, smoking, and/or alcohol in their associations with lipid levels.
In the present study, we aim to examine the association of the MTHFR C677 T, MTHFR A1298C, MTRR A66G, and MTRR His595Tyr, 4 functional gene polymorphisms, with blood lipids and whether interactions between these variants exist with gender and 2 common environmental factors, drinking and smoking, in Chinese patients with hypertension.
Patients and Methods
Study Population
A total of 340 patients with mild to moderate hypertension were enrolled from Dongzhi community, in Anhui Province, China, from April 2003 to January 2004. The average age of the participants was 50.3 ± 6.3 years. There were 147 men (43.2%) and 193 women (56.8%). In brief, hypertensive participants who met the following criteria were enrolled: (1) systolic blood pressure (SBP), 140 to 200 mm Hg or diastolic BP (DBP), 90 to 115 mm Hg; (2) aged 35 to 65 years; and (3) the participants were not taking medications known to affect serum lipid levels (lipid-lowering drugs such as statins or fibrates, beta-blockers, diuretics, or hormones) within 4 weeks before the study. Participants with a history of any of the following conditions were excluded: secondary hypertension, pregnancy, hypercalcemia, chronic CVD, chronic cerebrovascular disease, chronic liver or renal diseases, or body mass index (BMI) >33 kg/m2. The study was approved by the ethics committee of Anhui Medical University, Hefei, China. The purpose and procedures of the study were carefully explained to all the participants and a written informed consent was obtained.
Laboratory Determinations
Venous blood samples were drawn and collected in ethylenediaminetetraacetic acid tubes after a 14-hour fasting at 8:00
Genotyping of MTHFR and MTRR Polymorphisms
TaqMan allelic discrimination technique was used for detecting MTHFR C677T (Ala222Val), MTHFR A1298C (Glu429Ala), MTRR A66G (Ile22Met), and MTRR His595Tyr genotypes in our central laboratory. Universal reaction conditions for each genotyping are as follows: 4 ng dried DNA, 0.08 μL 40× assay locus-specific probe, and 2.0 μL TaqMan universal polymerase chain reaction (PCR) master mix made to a final volume of 4 μL with 1.92 μL of sterile water. The PCR cycle conditions consisted of initial denaturation at 95°C for 10 minutes, followed by 50 cycles of 92°C for 15 seconds and 60°C for 1 minute. All sample sets genotyped for each SNP in our present study had overall call rates of ≥95%, after excluding samples that consistently failed. Concordance of 100% was repeated for all samples’ quality control.
Statistical Analysis
Epidata 3.1 Chinese version was used for database design, data entry, and data check. Mean ± standard deviation were calculated for continuous variables. One-way analysis of variance and t test were used to compare the mean differences for continuous variables. The chi-square test was used for categorical variables. Allele frequency was determined via direct counting, and the standard goodness-of-fit test was used to test the Hardy-Weinberg equilibrium. Both TC and LDL-C followed normal distribution, while TG was log-normally distributed, not for HDL-C. A multivariate linear regression model was used to evaluate the effect of MTHFR and MTRR polymorphisms on the baseline TC, TG, HDL-C, and LDL-C levels before and after adjusting for possible confounders, including age, gender, and BMI. The interactions of the MTHFR and MTRR genetic variants and modifying factors (such as sex, smoking, and drinking) on serum lipid levels were tested by adding a product term in the linear models. Smokers were defined as those who had smoked more than 100 days or more than 100 cigarettes in his or her life. Drinkers were defined as those who had ever drunk any alcoholic beverage during his or her life. A q value measures the false discovery rate (FDR) by accepting the given test and every test with a larger P value, if they improve the FDR coded by SAS program. Differences were considered to be significant at P < .05. All the statistical analyses were carried out using the R packages.
Results
General Characteristics
Totally, 340 patients with hypertension, from Dongzhi community in Anhui Province, China, with both available genotypes and phenotypes were analyzed. The genotype distributions of 4 functional SNP loci including MTHFR (C677T and A1298C) and MTRR (A66G and His595Tyr) polymorphisms did not deviate from Hardy-Weinberg equilibrium (both P values >.1; Table 1). So in the following analysis, we tested all these 4 loci. Linkage disequilibrium (LD) plots showed that the MTHFR C677T and A1298C SNPs were not in complete LD (D′ = 1, r2 = .258); or MTRR A66G and His595Tyr were not yet in complete LD (D′ = 1, r2 = .091). The distributions of participants’ age, height, weight, BMI, baseline SBP and DBP, TC, TG, HDL, and LDL are shown in Table 2. Among participants with hypertension, 43.2% were male, 28.2% were current cigarette smokers, and 35.3% were current alcohol drinkers.
Hardy-Weinberg Equilibrium Tests for MTHFR and MTRR Polymorphisms.

Abbreviations: MTHFR, methylenetetrahydrofolate reductase; MTRR, methionine synthase reductase.
Basic Characteristics in Hypertensive Participants.

Abbreviations: BMI, body mass index; BP, blood pressure; TC, total cholesterol; LDL-C, low-density lipoprotein cholesterol; MTHFR, methylenetetrahydrofolate reductase; MTRR, methionine synthase reductase; TG, triglyceride; HDL-C, high-density lipoprotein cholesterol; SD, standard deviation.
Genotypes and Serum Lipid Levels in Total Population
As shown in Table 3, among the total population, the levels of TC and LDL-C were significantly different among the 3 genotypes in the MTRR A66G gene. Multiple linear regression showed that compared with the AA genotype carriers, the GG genotype carriers had lower serum TC levels (adjusted β ± standard error [SE]: −0.5 ± 0.2 mmol/L; P = .003) and LDL-C levels (adjusted β ± SE: −0.4 ± 0.2 mmol/L; P = .005). False discovery rate adjusted P values were .056 and .056, respectively. The levels of LDL were different among the 3 genotypes in MTHFR C677T gene. Multiple linear regression showed that the TT genotype carriers had higher serum LDL-C levels (adjusted β ± SE: 0.2 ± 0.1 mmol/L; P = .046) than the CC genotype carriers. The FDR-adjusted P value was .280. There were no significant differences in the levels of TG and HDL-C among the 3 genotypes in either MTRR A66G or MTHFR C677T (P > .05 for all).
Multiple Linear Regression of MTHFR and MTRR Polymorphisms and Serum Lipid Levels in Total Population.
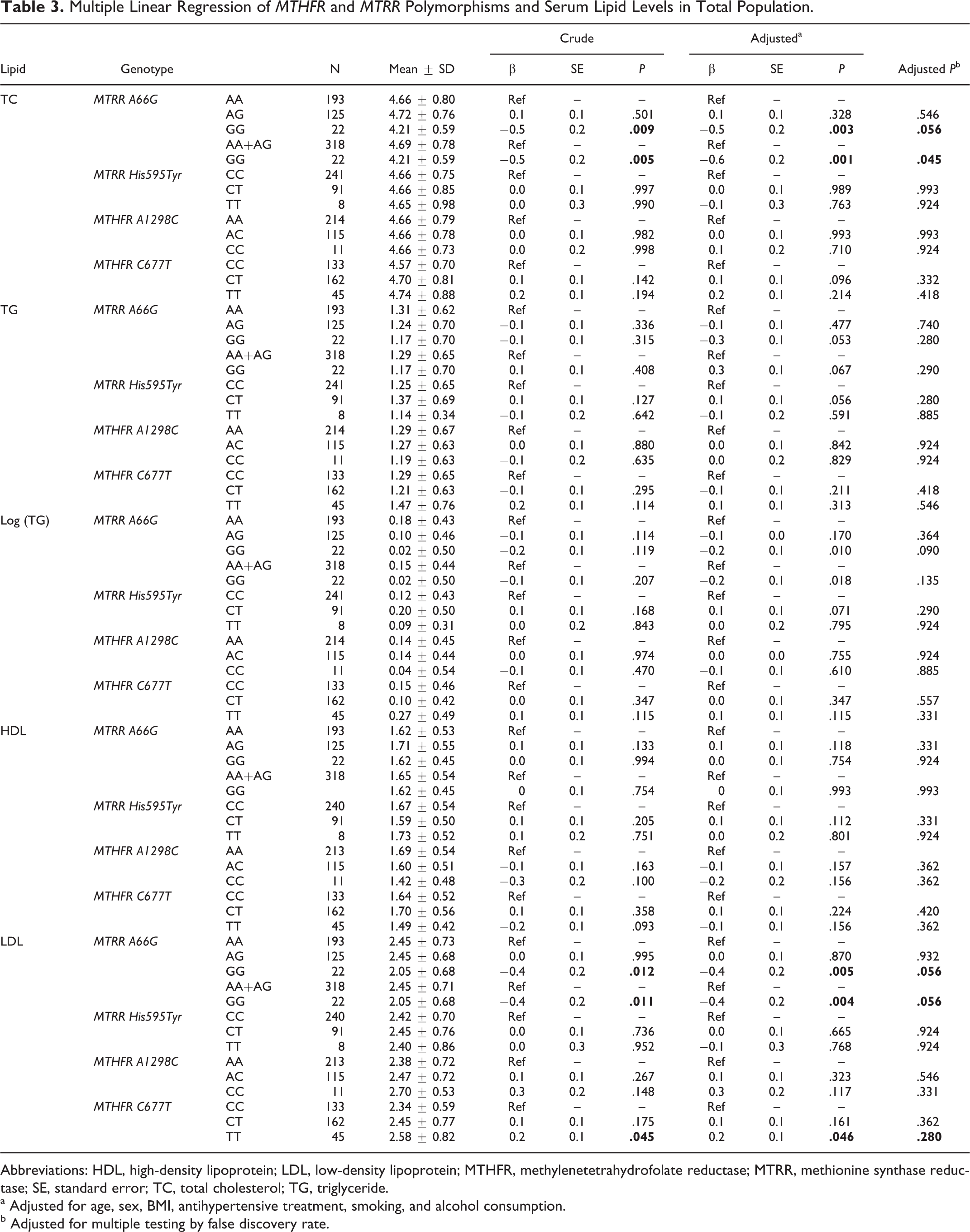
Abbreviations: HDL, high-density lipoprotein; LDL, low-density lipoprotein; MTHFR, methylenetetrahydrofolate reductase; MTRR, methionine synthase reductase; SE, standard error; TC, total cholesterol; TG, triglyceride.
a Adjusted for age, sex, BMI, antihypertensive treatment, smoking, and alcohol consumption.
b Adjusted for multiple testing by false discovery rate.
Gender-Specific Associations of Genotypes and Serum Lipid Levels
As shown in Table 4, in the female population, the levels of TC and LDL-C had significant differences among the 3 genotypes in the MTRR A66G gene. Multiple linear regression showed that compared with the AA genotype carriers, the GG genotype carriers had lower serum TC (adjusted β ± SE: −0.7 ± 0.2 mmol/L; P = .001) and LDL-C levels (adjusted β ± SE: −0.7 ± 0.2 mmol/L; P = .001). The FDR-adjusted P values were .006 and .006, respectively. The levels of TC and LDL-C were also different among the 3 genotypes in MTHFR C677T gene. Multiple linear regression showed that the TT genotype carriers had higher serum TC (adjusted β ± SE: 0.4 ± 0.2 mmol/L; P = .027) and LDL-C levels (adjusted β ± SE: 0.5 ± 0.2 mmol/L; P = .002) than the CC genotype carriers. The FDR-adjusted P values were .100 and .010, respectively. There were no significant differences in the levels of TG and HDL-C among the 3 genotypes in either MTRR A66G or MTHFR C677T (P > .05 for all).
Multiple Linear Regression of MTHFR and MTRR Polymorphisms and Serum Lipid Levels Stratified by Gender.
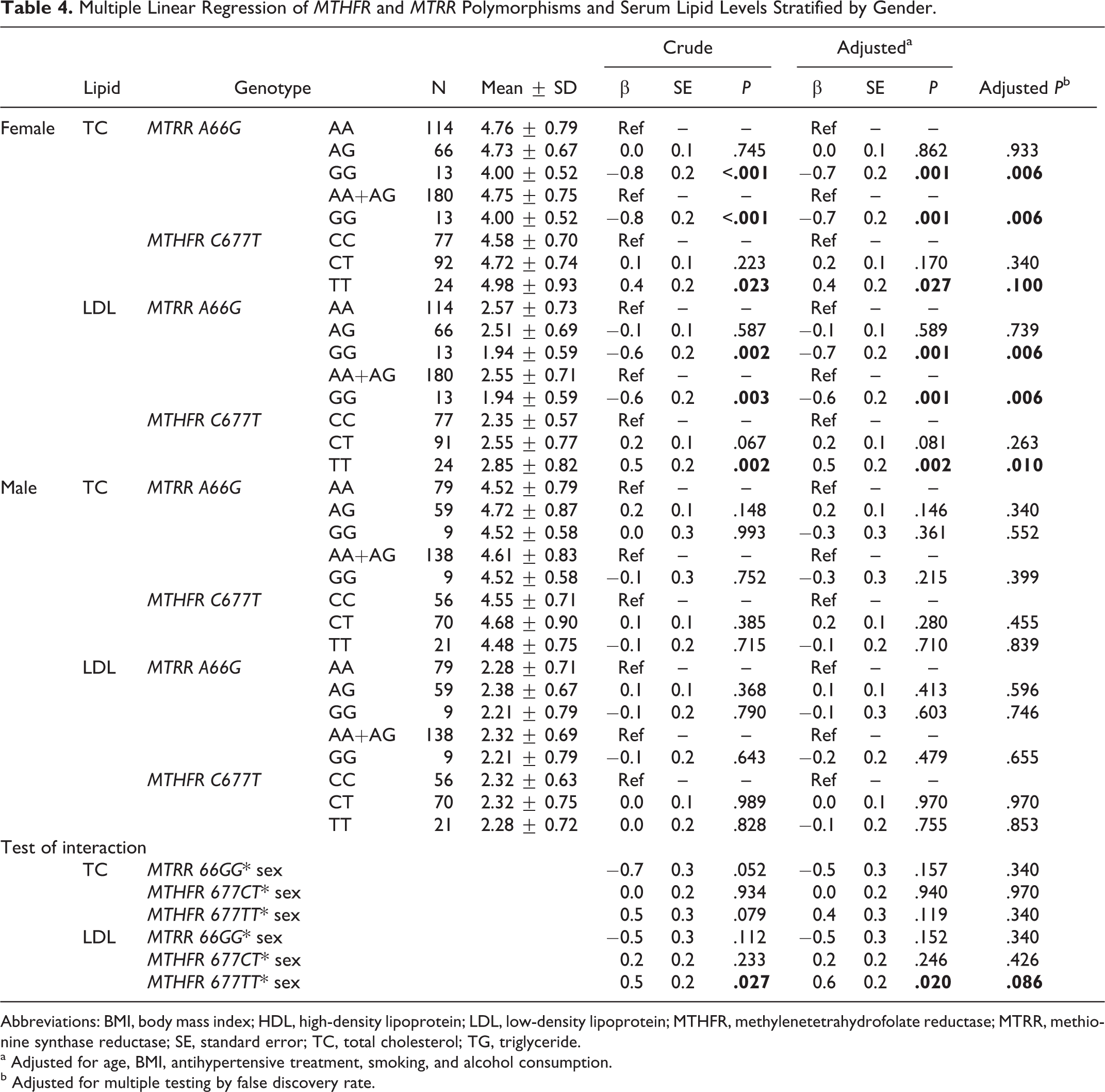
Abbreviations: BMI, body mass index; HDL, high-density lipoprotein; LDL, low-density lipoprotein; MTHFR, methylenetetrahydrofolate reductase; MTRR, methionine synthase reductase; SE, standard error; TC, total cholesterol; TG, triglyceride.
a Adjusted for age, BMI, antihypertensive treatment, smoking, and alcohol consumption.
b Adjusted for multiple testing by false discovery rate.
Smoking-Specific Associations of Genotypes and Serum Lipid Levels
In nonsmokers, as shown in Table 5, multiple linear regression showed that compared with the AA genotype carriers, the MTRR 66GG genotype carriers had lower serum TC levels (adjusted β ± SE: −0.6 ± 0.2 mmol/L; P = .004) and LDL-C levels (adjusted β ± SE: −0.6 ± 0.2 mmol/L; P = .002). The FDR-adjusted P values were .026 and .026, respectively. The MTHFR 677TT genotype carriers had higher serum TC levels (adjusted β ± SE: 0.3 ± 0.2 mmol/L; P = .041) and LDL-C levels (adjusted β ± SE: 0.4±0.1 mmol/L; P = .005) than the CC genotype carriers. The FDR-adjusted P values were .152 and .026, respectively. However, in smokers, no significant differences in lipid levels among the 3 genotypes were observed.
Multiple Linear Regression of MTHFR and MTRR Polymorphisms and Serum Lipid Levels Stratified by Smoking.
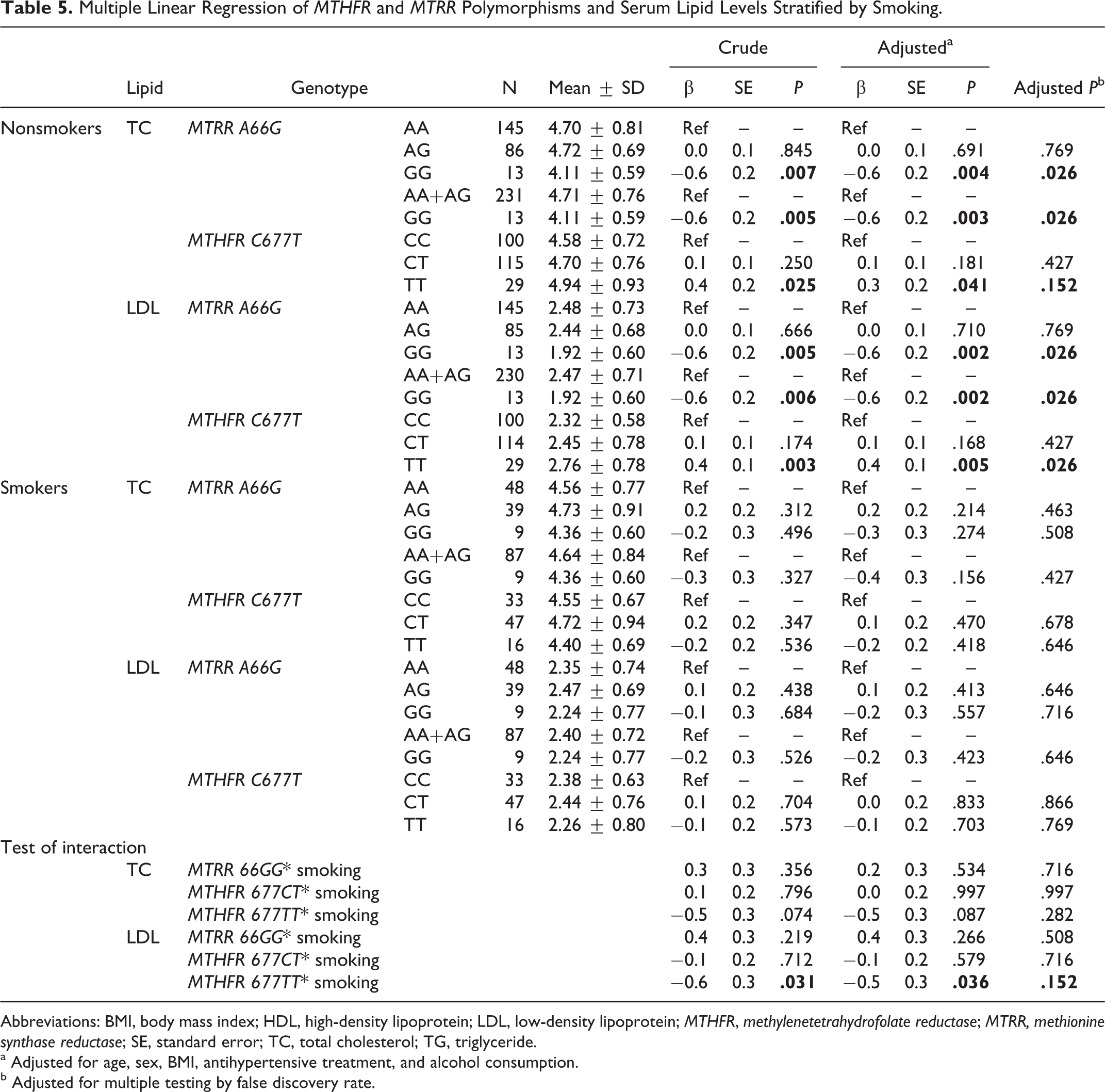
Abbreviations: BMI, body mass index; HDL, high-density lipoprotein; LDL, low-density lipoprotein; MTHFR, methylenetetrahydrofolate reductase; MTRR, methionine synthase reductase; SE, standard error; TC, total cholesterol; TG, triglyceride.
a Adjusted for age, sex, BMI, antihypertensive treatment, and alcohol consumption.
b Adjusted for multiple testing by false discovery rate.
Drinking-Specific Associations of Genotypes and Serum Lipid Levels
In nondrinkers, as shown in Table 6, multiple linear regression showed that compared with the AA genotype carriers, the MTRR 66GG genotype carriers had lower serum TC levels (adjusted β ± SE: −0.8 ± 0.2 mmol/L; P < .001) and LDL-C levels (adjusted β ± SE: −0.8 ± 0.2 mmol/L; P < .001). The FDR-adjusted P values were .0007 and .0007, respectively. The MTHFR 677TT genotype carriers had higher serum TC levels (adjusted β ± SE: 0.4 ± 0.2 mmol/L; P = .018) and LDL-C levels (adjusted β ± SE: 0.4 ± 0.2 mmol/L; P = .012) than the CC genotype carriers. The FDR-adjusted P values were .074 and .062, respectively. However, in drinkers, no significant differences in lipid levels among the 3 genotypes were observed.
Multiple Linear Regression of MTHFR and MTRR Polymorphisms and Serum Lipid Levels Stratified by Drinking.
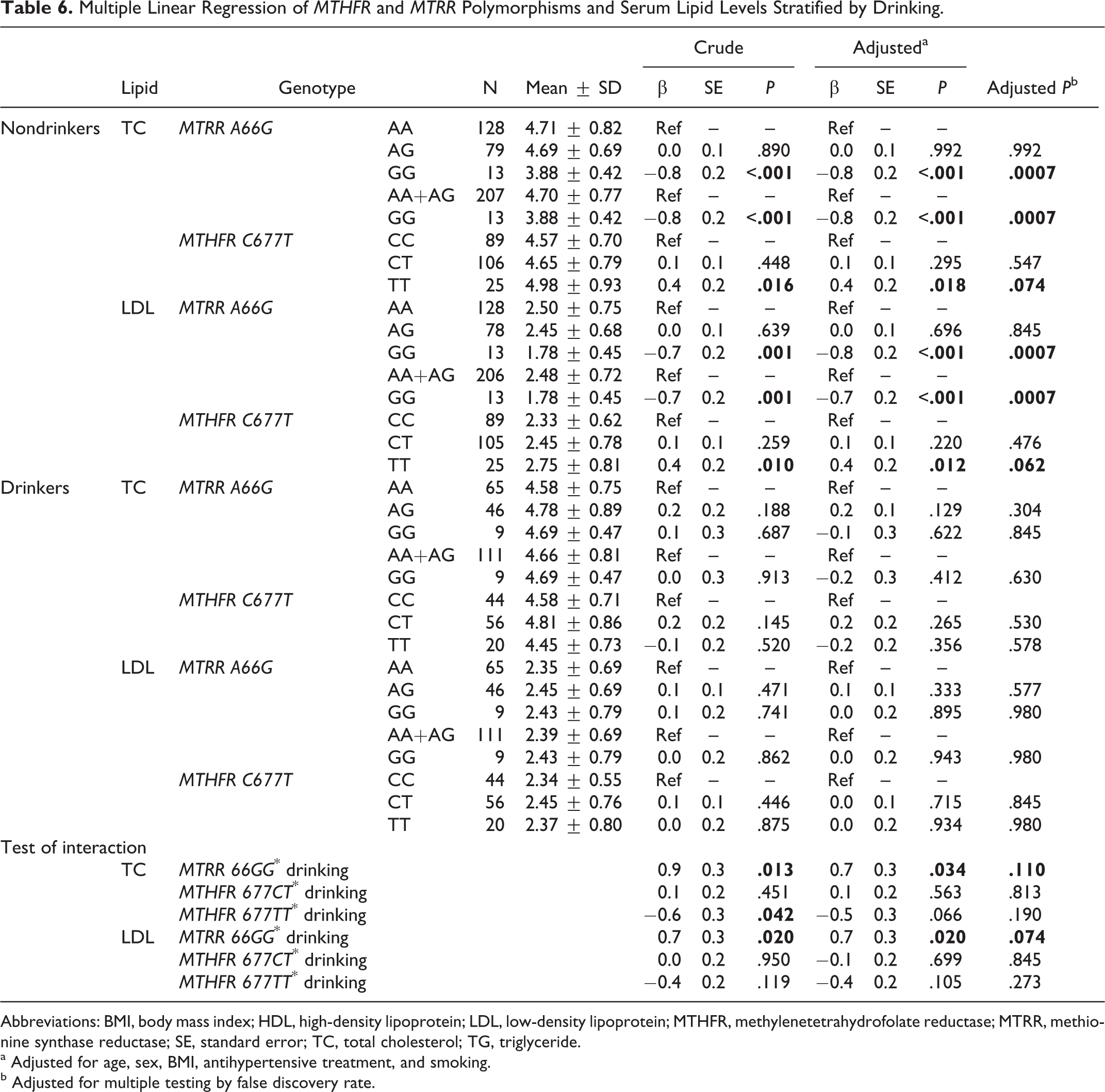
Abbreviations: BMI, body mass index; HDL, high-density lipoprotein; LDL, low-density lipoprotein; MTHFR, methylenetetrahydrofolate reductase; MTRR, methionine synthase reductase; SE, standard error; TC, total cholesterol; TG, triglyceride.
a Adjusted for age, sex, BMI, antihypertensive treatment, and smoking.
b Adjusted for multiple testing by false discovery rate.
Interactions of Modifying Factors and Genotypes in Serum Lipid Profiles
As shown in Tables 4, 5, and 6, we found significant interactions between genetic variants (MTRR A66G and MTHFR C677T) and modifying factors (sex, smoking, and drinking) in their associations with serum lipid levels. Table 4 showed that there was a significant interaction between MTHFR 677TT and sex in the association with serum LDL levels (P interaction = .02). A statistically significant interaction between MTHFR 677TT and smoking in the association with serum LDL levels was observed in Table 5 (P interaction = .036). We also identified the interactions between MTRR 66GG and drinking in the associations with serum levels of TC (P interaction = .034) and LDL (Table 6; P interaction = .02). However, after adjustment for multiple testing using the FDR, none of these interactions were significant.
Discussion
Our study suggests that MTHFR C677T and MTRR A66G gene polymorphisms may be important genetic determinants of serum lipid levels. There may exist significant interactions between MTHFR C677T and MTRR A66G variants and gender, smoking, and drinking consumption in the associations with serum lipid levels in Chinese hypertensive patients. However, after correction for multiple comparison tests, these interactions did not reach statistical significance. Our small samples may not have had sufficient power to estimate some of the interactions. It indicated that these findings may be validated in a larger sample.
The MTHFR C677T, one of the most common mutations, increases the risks of atherosclerosis, coronary heart disease, stroke, and other cardiocerebrovascular diseases. 21 –24 Hyperlipidemia is an independent risk factor for CVDs. The relationship between the MTHFR C677T and serum lipid profiles has been investigated in different populations. The MTHFR 677T allele is associated with unfavorable serum lipid profiles in a majority of studies 12,13,25 –29 but not in others. 30 –32 Yilmaz et al 12 genotyped MTHFR C677T polymorphism in 29 kidney transplant patients and found that the 677C allele has a protective effect on blood lipid levels, while the T allele has a harmful effect. Participants with the T allele compared to those with the CC genotype had significantly increased TC and LDL levels. Zhang et al 13 have shown that the levels of TC and LDL-C are different among the 3 MTHFR genotypes in both Bai Ku Yao and Han populations, the T allele carriers have higher serum TC and LDL-C levels as compared with the T allele noncarriers. Kawamoto et al 25 reported that the MTHFR gene polymorphism is a significant independent contributer to TC (P < .001) and HDL-C (P < .001). Zuntar et al 27 found that TC and TG are statistically different in MTHFR genotypes. Pitsavos et al 26 reported that oxidized (ox)-LDL levels were higher in those with the TT genotype compared to CC and CT genotypes. Before correction for multiple comparison tests, our findings consistently supported that the MTHFR 677TT genotype had higher LDL-C levels. However, the significant result disappeared after corrections. In short, apart from previous studies, our present study identified that the MTHFR 677TT genotype may be an independent risk factor for predicting higher serum lipid profiles.
The MTRR is another important enzyme in Hcy metabolism. The A66G mutation in this gene has been studied, but its role in Hcy regulation is still controversial. Several studies 14,15 reported that Hcy is higher in carriers of MTRR AA than in those carrying the other genotypes. Laraqui et al 33 found that the 66G allele contributes to a moderate increase in Hcy levels (P = .007). However, studies did show that Hcy concentration is not different among MTRR A66G genotypes. 34 Our study found that before corrections, the MTRR A66G locus AA genotype is associated with higher TC and LDL-C levels. After FDR corrections, these significant results were not observed. To our knowledge, this is the first report of associations between the MTRR A66G polymorphism and lipid levels.
Our present study possibly suggested that MTHFR 677TT may not only be related to higher TC and LDL-C levels but also proposed that the MTRR 66AA genotype may be a risk factor and predictor of higher TC and LDL-C levels. The potential underlying mechanism may involve (1) high levels of Hcy increase messenger RNA expression and enzyme activity of HMG-CoA reductase, 4 and this in turn activates 3 transcription factors, sterol regulatory element-binding protein 2 (SREBP-2), cyclic adenosine monophosphate response element-binding protein (CREB), and nuclear factor Y (NF-Y). Thus, HHcy-induced activation of SREBP-2, CREB, and NF-Y likely increases cholesterol biosynthesis by transcriptionally regulating HMG-CoA reductase expression in the liver, leading to hepatic lipid accumulation and subsequent hypercholesterolemia. (2) Elevated Hcy may induce endoplasmic reticulum stress and activate unfolded protein response and the SREBP in liver cells, endothelial cells, and arterial smooth muscle cells. 1,5 Activation of the SREBPs leads to increased expression of genes responsible for cholesterol/TG biosynthesis and uptake and intracellular accumulation of cholesterol. Our results seem to support that MTHFR C677T and MTRR A66G loci can affect serum TC and LDL-C metabolism through the influence of serum Hcy levels. However, a limitation of the present study was that it did not measure plasma Hcy levels, otherwise it could provide an effective and rational explanation for the hypothesis of the genotypes in association with serum lipid levels.
Other than MTHFR and MTRR, MS is a key enzyme of Hcy metabolic pathways. Huang et al’s study 28 suggested that compared with noncarriers, participants carrying MS 2756G alleles show a 3.1-fold risk of combined hyperlipidemia. The patients with hyperlipidemia with 2756G alleles even had a higher concentration of TC and LDL-C than those without 2756G alleles. The MS 2756G variant might increase the Hcy levels 35 and hence have a detrimental effect on lipid profiles. To fully understand the relation between Hcy-metabolizing genes and serum lipid profiles, further work is needed to investigate whether the MS 2756G variant is associated with serum lipid levels in our studied population.
It is well known that complex traits such as hyperlipidemia generally are caused by genetic factors and their interaction with the environment. Our study showed that the associations between MTHFR and MTRR polymorphisms and serum lipid levels may be gender specific. However, our models may not have had sufficient power to estimate some of the interactions with enough precision to achieve statistical significance, and these results require replication. Females with the TT genotype had significantly higher LDL-C levels when comparing to the 677CC genotype, but among men there was no apparent difference in LDL-C levels among the different genotypes. Although other gender by genotype interactions did not reach statistical significance, there were apparent trends toward interaction for MTHFR C677T genotypes and TC levels as well as MTRR A66G genotypes and both LDL-C and TC levels. For all of these, the effects of genotype appeared to be stronger among females than males. Gene by gender interactions could possibly result from different amounts of folic acid. Studies have consistently found that in women folic acid levels are significantly higher than in men, likely from the higher intake of folic acid. 36 The elevated folic acid in females can increase the difference in Hcy level between wild-type and mutant, which causes those with 677TT and 66AA to carry relatively higher Hcy levels and in turn produce higher lipid levels. Thus, in females, MTHFR 677TT and MTRR 66AA genotype loci can have more significant contributions to plasma Hcy levels and further lead to detrimental effects on serum lipid levels.
In this study, we investigated the relationship between the lipid levels and 2 environmental factors, smoking and drinking. Compared to those with the 677CC genotype, there was higher LDL-C concentration among those with the 677TT genotype among nonsmokers but not smokers. There was also a trend toward interaction between smoking and MTHFR 677TT in their associations with TC with a larger genotype effect among nonsmokers. However, after FDR corrections, the significant interactions between the MTHFR genotype and smoking in association with lipid levels disappeared. It suggested that our models may have lacked power to discriminate interactions with sufficient precision and require further replication. Nicotine and carbon monoxide can increase TG and free fatty acids, which pour into the liver and stimulate the synthesis of large amounts of TG and very-low-density lipoprotein cholesterol (VLDL-C). Decomposition of VLDL-C leads to the production of apo A, the main apolipoprotein of HDL-C. When the liver increases synthesis of VLDL-C, apo-A decomposition will increase, leading to decreased HDL-C levels. Thus, smoking-induced risk of hyperlipidemia can be at least due to the effect on HDL-C and LDL-C. 37 Smoking is an independent risk factor for HHcy, which was positively associated with increased Hcy concentrations. 18,19 Therefore, in the smoking population, smoking may increase Hcy level but weaken the relative differences in Hcy among the 3 genotypes and further distort associations between MTHFR C677T and MTRR A66G polymorphisms and lipid levels. In contrast, among nonsmokers, the independent and significant association between genotype and various indicators of lipid levels, such as TC and LDL, can be observed.
Additionally, we also observed that the effects of genotype appeared to be stronger among nondrinkers. Drinking can contribute to increased TC, accelerate the progress of atherosclerosis, and lead to increased incidence of CVDs risk. Wu et al 38 found that drinking causes an increased amount of TC, TG, and HDL-C values and a decreased LDL-C value. Drinking is associated with the metabolism of vitamin B6, vitamin B12, and folic acid, which will lead to plasma Hcy accumulation. Therefore, similar to smoking, drinking could also possibly mask the associations between genotype and the indicators of lipid levels, but for nondrinkers a significant association can more easily be observed.
In summary, our results seemed to support the contribution of both genetic and environmental factors in regulating the levels of lipids. Before FDR corrections, we found that participants with MTRR 66GG have significant lower TC and LDL levels, while those with MTHFR 677TT have significantly higher TC and LDL levels. We also have demonstrated evidence of interactions between MTHFR C677T and MTRR A66G polymorphisms and gender, smoking, and drinking consumption in their associations with serum lipid levels. However, these significant findings disappeared after corrections. Our study suggested that MTHFR C677T and MTRR A66G may be potential markers for lipid expression and can be good targets for applications such as disease risk prediction, individualized drug treatment and development.
Footnotes
Acknowledgments
We gratefully acknowledge the assistance and cooperation of the faculty and staff of the Anhui Medical University and thank all of the participants in our study. This study was conducted in accordance with the current regulations of the People’s Republic of China.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: National Natural Science Foundation of China (Grant No. 81141116 and 30700454).