
Abstract
Bovine respiratory disease complex (BRDC) is a multi-etiologic disease that significantly impacts the cattle industry. Nasopharynx-associated lymphoid tissue (NALT) is the nasal mucosal immune system that protects against various pathogens; however, reports on its pathology are limited. We compared the pathologic impact of BRDC-related viruses on the NALT and lungs of 10 pneumonic and 3 negative control Japanese black (JB) calves. Three bovine viral diarrhea virus (BVDV) persistently infected Holstein calves were examined as positive control; all 3 calves had BVDV in situ hybridization (ISH)-positive signals in NALT follicular macrophages and lymphocytes, suggesting viral replication in these cells. NALT and lungs of pneumonic JB calves had weak-positive BVDV results in PCR and negative results in ISH, indicating late-stage transient BVDV infection. The finding of BVDV in unvaccinated pneumonic JB calves confirmed the involvement of a BVDV field strain. PCR detection of bovine coronavirus (BCoV) and bovine respiratory syncytial virus (BRSV) in NALT along with ISH BCoV-positive signals in NALT epithelial cells confirms infection by those viruses. Pneumonic JB calves had nasopharyngitis and pneumonia, with the same bacteria present in nasopharyngeal swabs and lungs, indicating that, in these animals, the bacteria proliferating in the nasopharynx probably migrated to the lungs via inhalation. BVDV transient infection in the NALT may induce local immunosuppression; BCoV and BRSV infections are thought to damage epithelial cells, facilitating bacterial infection of the NALT and lungs. Our results confirm that bovine NALT is a location for viral replication and may be associated with BRDC development in cattle.
Keywords
Bovine respiratory disease complex (BRDC) refers to a range of clinical diseases and lesions of the respiratory tract of cattle. 41 BRDC has a multifactorial etiology involving complex interactions among numerous factors, including environmental stressors and both viral and bacterial pathogens. 17 The diversity of diseases associated with BRDC has made it difficult to determine the roles of each factor, because although several factors appear necessary, they are insufficient to generate BRDC on their own. Other studies have suggested that infections involving multiple pathogens, including viruses and bacteria, may occur simultaneously or sequentially, preceded by stress.16,26,41 Stressors, such as long-distance transportation or overcrowding, raise the risk and severity of BRDC through compromised respiratory defense mechanisms that allow pathogens to emerge.16,26,61 Viruses that are implicated most frequently in BRDC include bovine viral diarrhea virus (BVDV; Flaviviridae, Pestivirus), bovine coronavirus (BCoV; Coronaviridae, Betacoronavirus 1), bovine respiratory syncytial virus (BRSV; Pneumoviridae, Orthopneumovirus bovis), bovine parainfluenza virus 3 (BPIV3; Paramyxoviridae, Respirovirus bovis), bovine adenovirus 3 and 7 (BAV3, Adenoviridae, Mastadenovirus bostertium; BAV7, bovine adenovirus F), and bovine alphaherpesvirus 1 (BoAHV1; Orthoherpesviridae, Varicellovirus bovinealpha1).23,27,33,71 Primary infection with one or more viruses alters the host’s innate and adaptive immune responses and/or damages the respiratory mucosa. 23
Impairment of the immune response of the host via stress and/or primary viral infection enables bacteria that are normal nasopharyngeal commensals to colonize deeper respiratory tissues, replicate, and secrete toxins in the lower airways that cause tissue destruction and inflammation, subsequently leading to the development of BRDC. 9 Commensal bacterial pathogens that are isolated frequently in association with BRDC include Mycoplasmopsis (Mycoplasma) bovis, Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni.36,46 Although mycoplasmas can be the predominant bacteria in healthy calves, they can also predominate in calves diagnosed with BRDC. 35 The prevalence of M. bovis infection is increasing in Japan. 27
BVDV is a ubiquitous infectious agent that causes multi-organ infection due to its ability to infect and replicate in immune cells such as macrophages and lymphocytes.2,9 BVDV infection can occur as either transient infection (TI) or calves may be persistently infected (PI). 69 TI is a postnatal infection, which may be subclinical, but clinical signs, such as mild fever, leukopenia, anorexia, and decreased milk production, can occur. 69 Animals infected with BVDV in utero during pregnancy can be born as PI animals.34,69 PI animals are significant virus reservoirs and play a crucial role in maintaining BVDV in the cattle population, given that they continuously shed large amounts of virus in all body secretions.25,69 PI calves have viral antigens in various tissues, especially lymphoid tissue, which may not be associated with lesions.31,32 PI calves may appear normal, but they are usually weak, stunted, and susceptible to many diseases, such as pneumonia, due to compromised immune responses.25,69
BRSV and BCoV are important pathogens associated with BRDC.24,33,38 BRSV replicates predominantly in ciliated respiratory epithelial cells and type II pneumocytes.65,66 As a primary pathogen, BRSV by itself can induce severe respiratory disease and pneumonia in calves, potentially accompanied by infection with secondary bacterial pathogens.50,74 BRSV increases the adherence of P. multocida to epithelial cells. 56 In cattle, BRSV infection causes atypical interstitial pneumonia with severe bronchitis, bronchiolitis, epithelial necrosis, and syncytia formation.52,60
BCoV is a pneumoenteric virus known to cause 3 different clinical diseases: severe calf diarrhea, winter dysentery in adult cattle, and respiratory tract infections, including BRDC, in cattle of various ages.10,49,54 BCoV has been detected in nasal secretions and lungs of cattle with pneumonia and/or from feces.20,21,62 BCoV has been identified as an important contributor to BRDC in Japan.27,38
Mucosal surfaces, such as those of the respiratory and digestive systems, are constantly exposed to a variety of pathogens and have a specialized immune system known as mucosa-associated lymphoid tissue (MALT). 40 Nasopharynx-associated lymphoid tissue (NALT), a type of MALT, is found in the nasal mucosa of rodents, pigs, and humans, and plays a vital role in the defense against airborne pathogens.12,51 NALT is a lymphoid structure located in the roof of the nasopharynx in pigs and in the floor of the nasal cavity in rats, hamsters, and mice.30,42 NALT lymphoid follicles are positioned close to the mucosal surface and are comprised of B lymphocytes interspersed with interfollicular T lymphocytes and dendritic cells (DCs).11,30 NALT lymphoid follicles are covered by follicle-associated epithelium (FAE), which is heavily infiltrated by lymphocytes, and includes morphologically distinct M cells that are specialized in the uptake of antigens. 70 M cells are among the most important epithelial cells involved in the adhesion, uptake, and sampling of antigens at mucosal surfaces.30,53,70 Luminal antigens are transported across the epithelium by M cells and then captured by DCs that reside beneath M cells to initiate the mucosal immune response and production of antigen-specific IgA by B cells.30,53,70
NALT is considered a constitutive structure of the nasal immune system. NALT has been identified in small laboratory animals, pigs, sheep, and humans of different ages without relation to antigen stimulation and plays a vital role in host defense against airborne pathogens.12,30,42,76 NALT lymphoid follicles have also been detected in healthy and respiratory-diseased pigs. 42 The NALT of rats serves as a highly effective route for inducing mucosal immunity because it is the location of IgA antibody responses at the mucosal effector site.53,72 Human NALT is the first lymphoepithelial barrier and exerts “gate control” of airborne antigens, including respiratory viruses such as SARS-CoV-2. 15 Histologic analysis of the upper respiratory tract (URT) mucosa, the gateway for pathogen invasion, is useful for characterizing the pathogenesis of infectious diseases. However, the nasal mucosa of most farm animal species has not been investigated, as it is surrounded by several bones that make it difficult to separate from the nasal cavity. 30 In addition, the preparation for histology involves demineralization steps that further inhibit detection of pathogen-associated antigens and nucleic acids. 30 The difficulty of gaining access to the NALT makes histologic examination of this tissue challenging.
The NALT has been described as the initial site for foot-and-mouth disease virus (FMDV) replication, as this is where FMDV antigen was observed. 8 Porcine circovirus 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) have been detected in the NALT of pigs, and these viruses are thought to induce local immunosuppression in the URT.42,64 PCV2 and PRRSV target macrophages.39,42,64
Here we focused on BVDV, which also targets macrophages, 47 as the virus is thought to induce local immunosuppression in the URT, similar to that caused by PCV2 and PRRSV. BVDV replication within the lymphoid follicles reduces the function of macrophages and induces apoptosis of lymphocytes. 1 However, although the NALT is considered a critical location for early viral replication, we found no reports on the role of NALT in the pathogenesis of BRDC-related pathogens. Thus, we analyzed the pathologic impact of BRDC-related viruses in the nasopharynx, including NALT. We also attempted detection of BRDC-related viruses (BVDV, BCoV, and BRSV) in the NALT and lungs, and attempted isolation of pathogenic bacteria from nasopharyngeal swabs and the lungs to further elucidate the pathogenesis of BRDC.
Materials and methods
Animals and sampling
Ten pneumonic Japanese black (JB) calves, 3-wk- to 18-mo-old, were collected from several farms in Miyazaki Prefecture, Japan, between February 2022 and May 2023 and examined at autopsy. Six calves were unvaccinated, and 4 had been vaccinated with respiratory 5-component live vaccines, including attenuated BVDV1, BRSV, BoAHV1, BPIV3, and BAV7. Three Holstein PI calves, 1–5-mo-old, 1 male and 2 females, were examined as positive controls for BVDV. The 3 calves were clinically healthy and had been culled from their herd after being confirmed as PI calves. For PI calf confirmation, a BVDV antigen ELISA was performed on ear punch samples of newborn calves, and the whole blood of positive calves was retested after 2 wk. After testing positive in the retest, the calf was considered a PI calf. They also did not have lesions at autopsy. Positive controls for BCoV and BRSV were unavailable, as these viruses are not persistent in animals, unlike BVDV. As a negative control group, NALT and lungs of 3 JB, 3–16-mo-old calves without respiratory signs were collected from farms in Miyazaki Prefecture, Japan, and tested negative for the investigated viruses.
To facilitate NALT sampling, we used a unique dissection procedure. First, an incision was made at the caudal end of the hard palate to expose the nasopharynx, then the incision was extended to the soft palatine tonsil. The NALT was located on the roof of the nasopharynx. NALT and lung samples were fixed in 4% paraformaldehyde (PFA), processed routinely, and sections cut for histologic examination and in situ hybridization (ISH). ISH was performed to confirm the distribution of infected cells in the NALT. Fresh NALT and lung tissues were stored at −80°C and examined for BRDC-related viruses by reverse-transcription PCR (RT-PCR). Nasopharyngeal swabs and lung samples were processed for bacteriologic examination.
Histologic examination
Tissue sections (4-µm thick) were stained with H&E and examined using a light microscope. NALT was examined for goblet cell hyperplasia, loss of cilia, exocytosis, and formation of micro-abscesses to determine the severity of nasopharyngitis (Suppl. Table 1). To determine the type of pneumonia, lung tissue was assessed for the type of exudate, bronchial and alveolar epithelial degeneration, thickening of the interstitial tissue, and the extent of necrosis in the lung parenchyma.
ISH
ISH probes were produced via amplification of the nonstructural protein NS3 region of BVDV and the spike (S) glycoprotein gene, as described previously.24,73 The sequence of the T7 promoter region was added to the 5′-region on the anti-sense primers. 75 PCR products were in vitro transcribed using T7 RNA polymerase. Digoxigenin (DIG)-labeled cRNA probes were prepared using an RNA labeling kit (Roche).
ISH for BVDV and BCoV was performed as described previously, 59 with slight modifications. Briefly, deparaffinized sections were digested with proteinase K, postfixed in 4% PFA, treated with 0.2 N HCl, and acetylated with 0.25% acetic anhydride in 0.1 M triethanolamine. The sections were then dehydrated, air-dried, and hybridized with a 10-fold diluted DIG-labeled RNA probe for 16–18 h at 37°C. After hybridization, the sections were immersed in 50% formamide/2× saline–sodium citrate buffer (2× SSC) and then washed sequentially in 2× SSC, 0.2× SSC, and 0.1× SSC. Biotin tyramide working solution (Akoya Biosciences) was used for sensitization. The sections were then washed with Tris–NaCl–Tween buffer, and the deposited biotin was visualized using anti-DIG horseradish peroxidase–labeled streptavidin (Dako) in Tris–NaCl blocking buffer. Finally, color was developed using a DAB kit, and the specimens were counterstained with hematoxylin.
To evaluate the specificity and sensitivity of the BVDV probe in ISH, serial sections of the tracheobronchial lymph nodes of a Holstein PI calf were prepared and analyzed using ISH, with hybridization buffer alone without DIG-labeled probe for one section and RNase treatment before hybridization for another section. One section was used for competitive inhibition analysis using a DIG-labeled probe and a fluorescein isothiocyanate (FITC)-labeled probe with the same sequence as the DIG-labeled probe and 20 times the amount of the DIG-labeled probe. All reactions were then compared to the usual protocol. The specificity and sensitivity of the BCoV probe were evaluated using the same method.
RT-PCR
To detect BRDC-related viruses, RNA was extracted from 10% homogenates of fresh NALT and lung samples. Two PCR primer pairs were used to detect BVDV, as described previously.48,68 BCoV- and BRSV-specific primers were determined based on previous reports.24,65 Holstein PI calves were used as a positive control for BVDV; the respiratory 5-component live virus vaccine was used as a positive control for both BVDV and BRSV.
Bacteriologic examination
Nasopharyngeal swabs and lung tissues of 6 JB calves were subjected to bacteriologic examination according to a previously reported method.4,6,7,28,55,63 In brief, the samples were inoculated onto Columbia agar medium with 5% bovine-defibrinated blood or trypticase soy agar as a basal medium. The inoculated samples were cultured in 2 ways: aerobically or with 5% CO2 gas at 37°C for 48–72 h. The isolated bacteria were purified and subjected to bacterial identification tests. For Mycoplasma isolation, the same specimen was inoculated into 0.002% DNA Hayflick medium, modified Taylor–Robinson medium, and BHL medium, and the tubes were capped tightly. After the medium changed color, the samples were spread on agar medium and cultured with CO2 gas at 37°C under high-humidity conditions for 3–5 d. The isolated bacteria were then characterized.
Results
Anatomically, bovine NALT is located in the roof of the nasopharynx and dorsal side of the soft palate (Fig. 1A). Macroscopically, NALT is a cauliflower-like structure of longitudinal raised folds of the surface epithelium of the nasopharynx, with the soft palatine tonsil located laterally (Fig. 1B, 1C). The NALT folds are longer in the rostral portion of the nasopharynx (nasal side) than the caudal portion (pharyngeal side), reflecting the higher density of NALT on the nasal side. Histologically, NALT is covered with stratified columnar epithelium (FAE), which covers lymphoid follicles (Fig. 1D). NALT crypts are lined by stratified columnar epithelium (Fig. 1E).
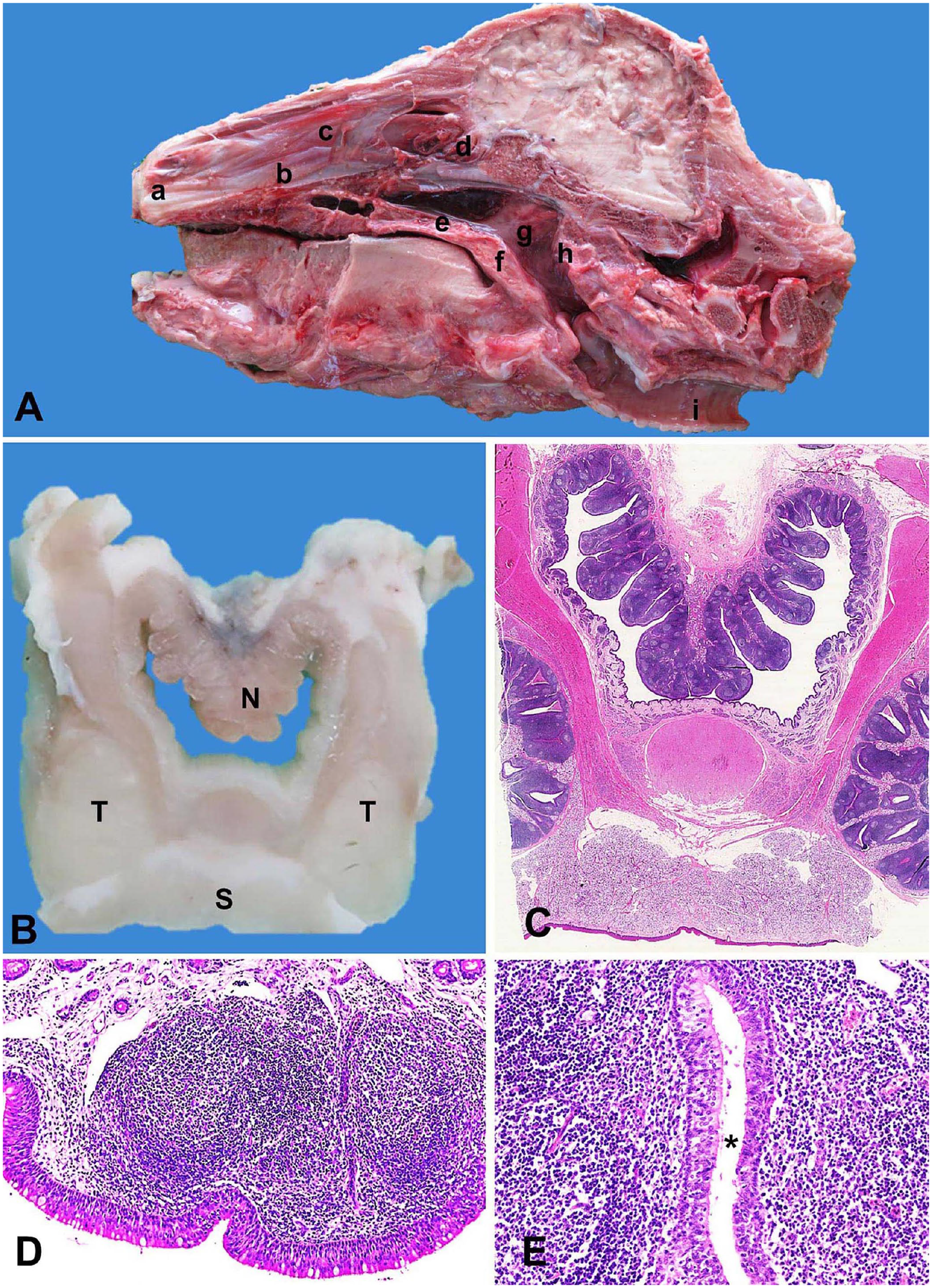
Bovine nasopharynx-associated lymphoid tissue (NALT).
Histologic analysis of the nasopharynx, including NALT, revealed moderate-to-severe nasopharyngitis in the pneumonic JB calves 1–10; negative control JB calves 11–13 did not have signs of nasopharyngitis (Table 1). Severe nasopharyngitis in calf 7 was marked by complete loss of cilia, with formation of micro-abscesses (Fig. 2A). Conversely, in negative control calf 13, the epithelium was intact with normal cilia and minimal exocytosis (Fig. 2B). In calf 3, severe nasopharyngitis was observed, with copious mucus mixed with several types of inflammatory cells, particularly neutrophils and mononuclear cells, accumulated in the NALT crypts, with goblet cell hyperplasia and degeneration of the epithelial cells lining the crypts (Fig. 2C). In contrast, the crypts of negative control calf 11 were clear and without exudate (Fig. 2D). In addition, follicular hyperplasia with active germinal centers was observed in calf 4 (Fig. 2E), in contrast to negative control calf 11 (Fig. 2F).
Histopathologic findings in nasopharynx-associated lymphoid tissue (NALT) and lung tissue of pneumonic (1–10) and negative control (11–13) Japanese Black calves.
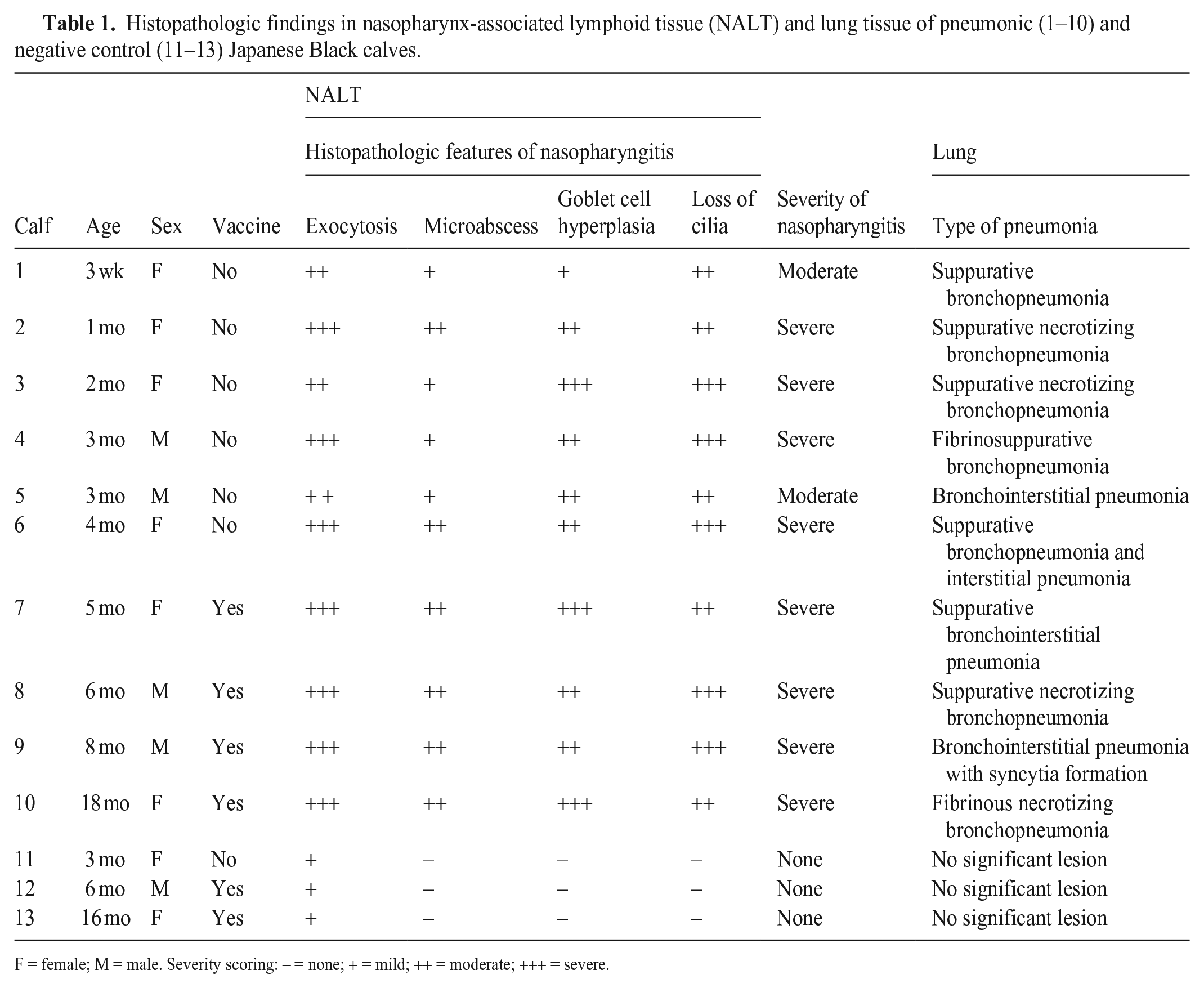
F = female; M = male. Severity scoring: – = none; + = mild; ++ = moderate; +++ = severe.
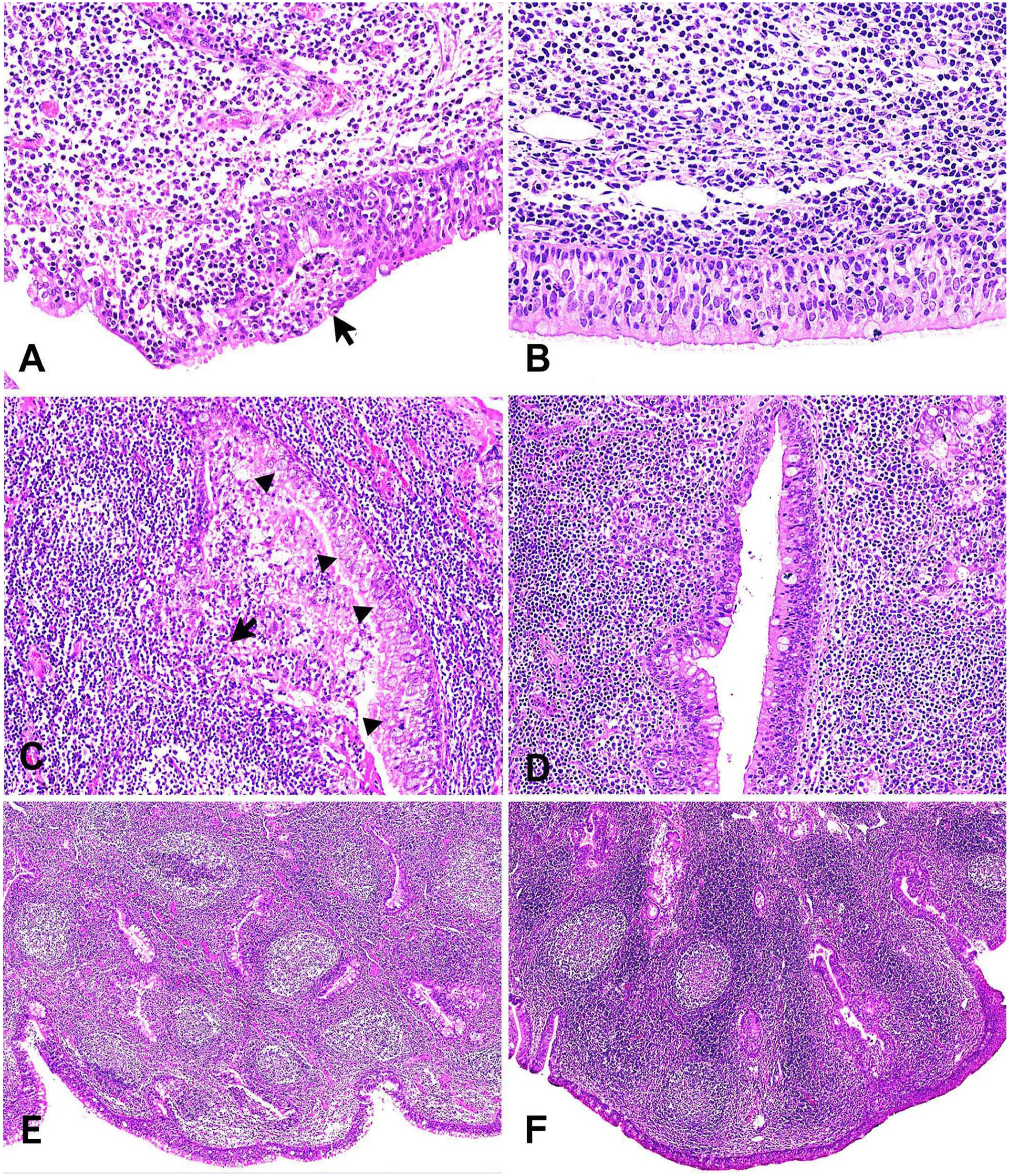
Histopathologic findings in the nasopharynx-associated lymphoid tissue (NALT) of pneumonic and negative control Japanese Black calves. H&E.
No BVDV-positive signals were detected in the NALT of any of the pneumonic JB calves using ISH. In contrast, the NALT of all 3 Holstein PI calves had BVDV-positive signals in the center of lymphoid follicles (Fig. 3A, 3B). Moreover, the BVDV-positive cells were morphologically similar to macrophages. BVDV-positive signals were also detected in the tracheobronchial lymph nodes of Holstein PI calves. BCoV-positive signals were detected in sloughed epithelial cells in the NALT crypts of calf 2 (Fig. 3C, 3D). The NALT epithelial cells in calf 10 had ciliary extinction with exocytosis (Fig. 3E, 3F). Testing of the specificity and sensitivity of the BVDV-ISH probe revealed clear BVDV-positive signals in the section of a tracheobronchial lymph node of a Holstein PI calf in which normal concentrations of DIG-labeled probe and hybridization buffer were used in accordance with the usual procedure (Suppl. Fig. 1A). However, no positive signals were detected using the other treatments (Suppl. Fig. 1B–D). Testing of the specificity of the BCoV-ISH probe had equivalent results.
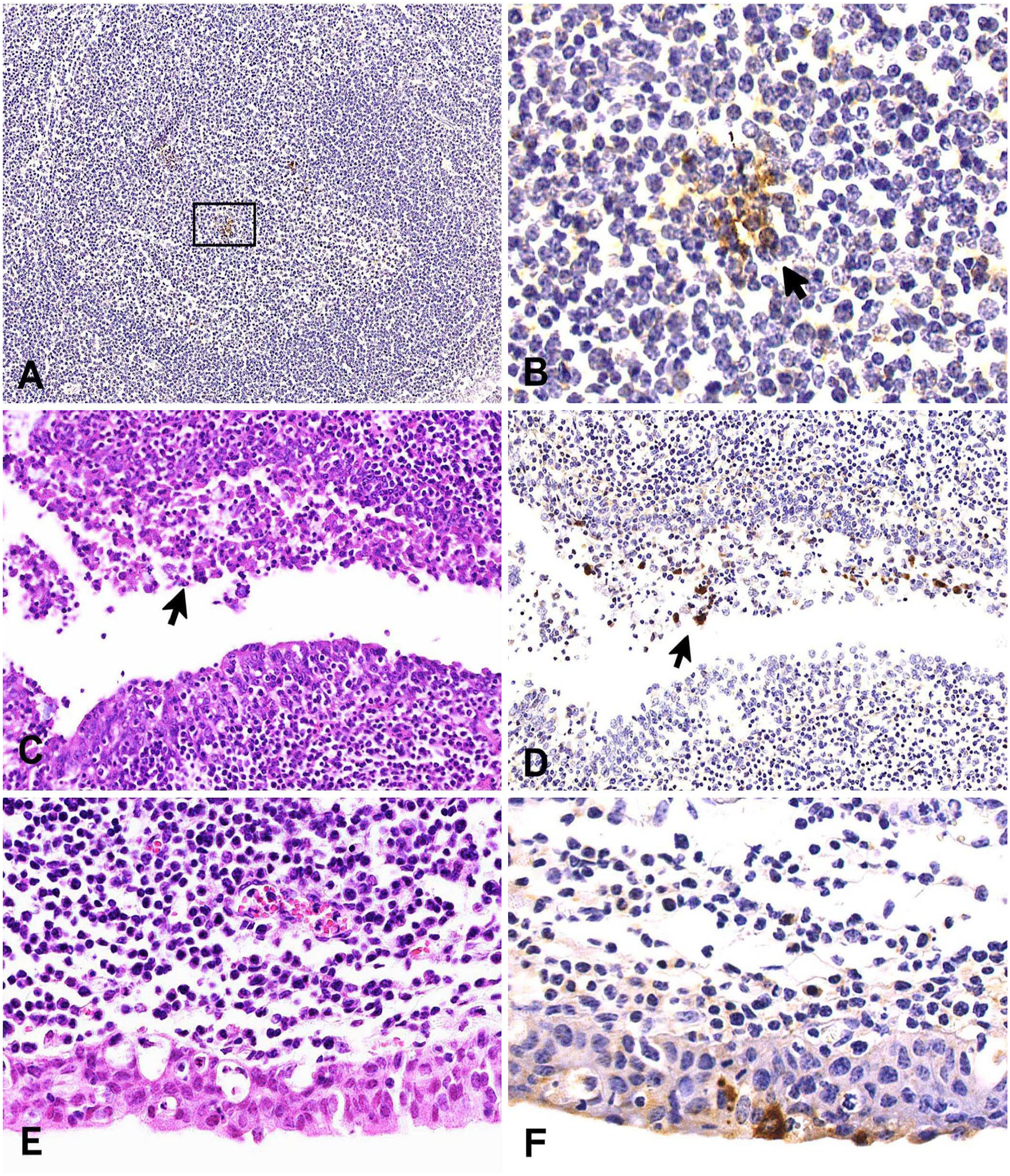
Histopathologic findings and in situ hybridization (ISH) results for analyses of the nasopharynx-associated lymphoid tissue (NALT) for BVDV (A, B) and BCoV (C–F).
The lungs of all pneumonic JB calves had lesions, whereas the lungs of negative control calves had no significant lesions (Table 1). Bronchial-associated lymphoid tissue (BALT) hyperplasia was only clearly evident in one of the pneumonic JB calves. Normal lungs with clear alveoli and bronchioles were observed in negative control calf 13 (Fig. 4A). Features observed in calf 1 included suppurative bronchopneumonia with abundant neutrophils, macrophages, and cellular debris within the lumens of the bronchi, bronchioles, and alveoli, and BALT hyperplasia (Fig. 4B). Calf 4 had fibrinosuppurative bronchopneumonia in the form of fibrinocellular exudates and distension of the interlobular septa. Accumulation of bacterial clusters with cellular debris and fibrin in the alveoli was also observed in calf 4 (Fig. 4C). Suppurative necrotizing bronchopneumonia with areas of caseous necrosis filled with hypereosinophilic granular debris resembling classical lesions associated with M. bovis infection was observed in calves 2, 3, and 8 (Fig. 4D). In calf 6, suppurative bronchopneumonia and interstitial pneumonia resulted in an abundance of neutrophils within the lumen of the bronchi, bronchioles, and alveoli. Other features in this calf included expansion of the alveolar wall by large numbers of macrophages and lymphocytes, and pneumocyte type II hyperplasia (Fig. 4E). Calf 9, with bronchointerstitial pneumonia, had large multinucleate syncytia characteristic of BRSV in the alveolar spaces and bronchiolar lumens (Fig. 4F). In most of the pneumonic JB calves, BVDV was detected in both the NALT and lungs, excluding calves 2 and 4, in which BVDV was detected solely in the NALT; calf 9 was negative in both tissues (Table 2). However, gel electrophoresis analysis of NALT samples from unvaccinated JB calves had a notably weak band unlike the positive control (vaccine and Holstein PI calves; Fig. 5). BRSV testing was positive in the NALT of 4 calves and lungs of 3 calves; BCoV was positive in the NALT of 2 calves and lungs of only 1 calf.
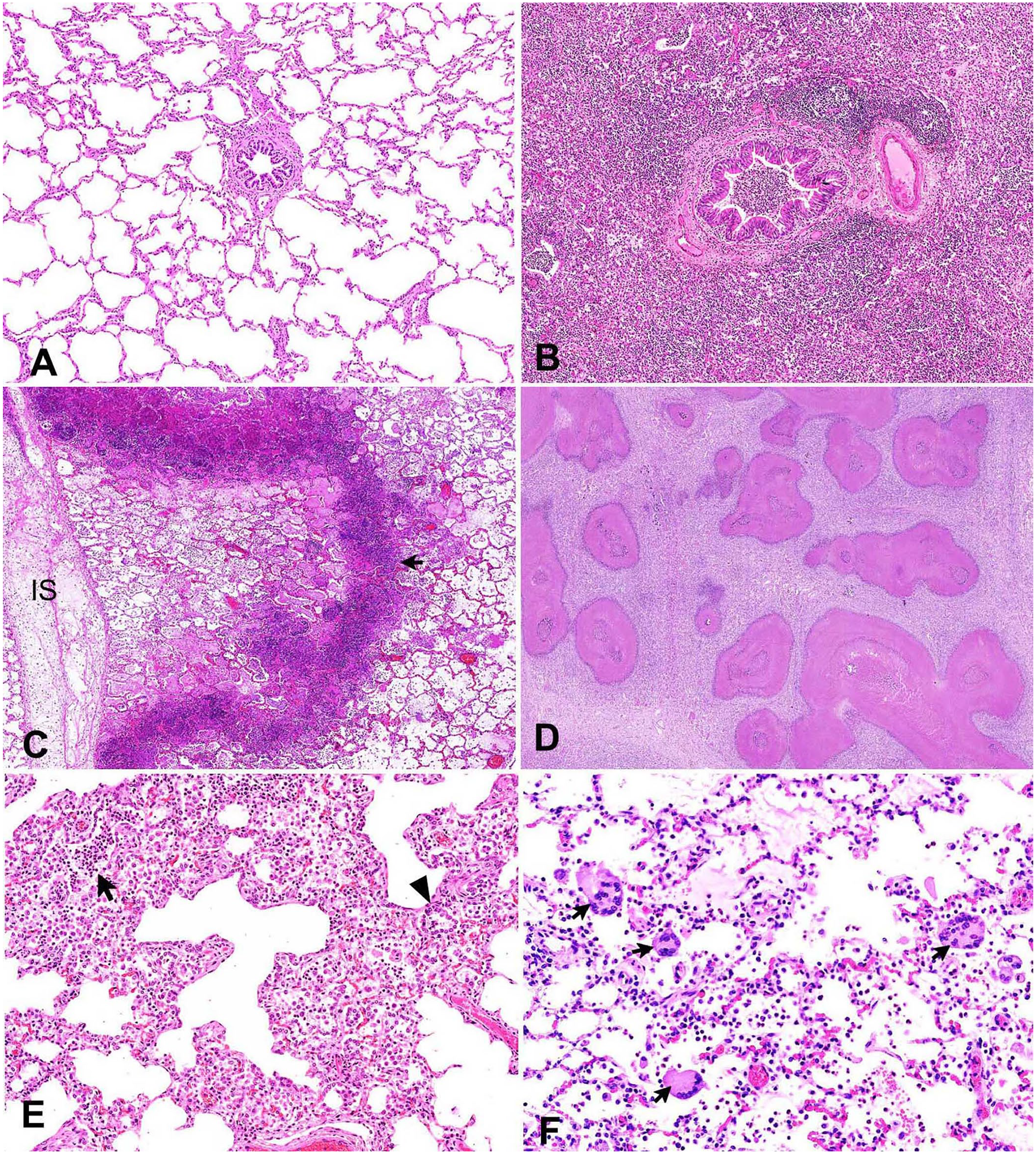
Histopathologic findings in the lungs of the pneumonic and negative control Japanese Black calves. H&E.
RT-PCR of bovine respiratory disease complex–related viruses in nasopharynx-associated ly mphoid tissue (NALT) and lung tissue of pneumonic (1–10) and negative control (11–13) Japanese Black calves.

BCoV = bovine coronavirus; BRSV = bovine respiratory syncytial virus; BVDV = bovine viral diarrhea virus; RT-PCR = reverse-transcription PCR; + = positive; ± = weak positive; – = negative.
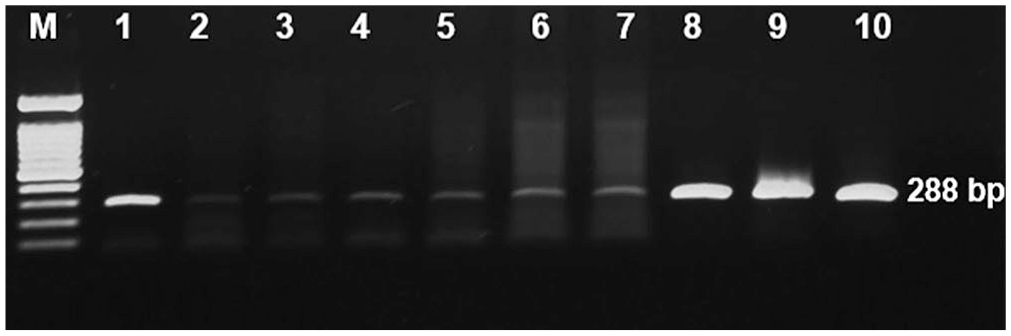
Electrophoretic analysis of RT-PCR amplification products of BVDV. Comparison of the band intensity in nasopharynx-associated lymphoid tissue (NALT) samples between unvaccinated pneumonic Japanese Black (JB) calves and positive control (Holstein PI) calves. Lanes: M = molecular weight markers (1.5 kb); 1 = vaccine (amplified product, 288 bp); 2–7 = NALT of unvaccinated pneumonic JB calves; 8–10 = NALT of Holstein PI calves.
M. bovis was detected in nasopharyngeal swabs and lungs of all pneumonic calves, except calf 9 (Table 3). M. haemolytica was detected along with M. bovis in calf 4. In addition, T. pyogenes and Streptococcus suis were isolated from the nasopharyngeal swab and lung samples of calf 8. Finally, Moraxella sp. and Pseudomonas aeruginosa were detected in the nasopharyngeal swab and lung samples of calves 9 and 10, respectively.
Bacteriologic examination of nasopharyngeal swab and lung tissue of pneumonic Japanese Black calves.

= positive; – = negative.
Trueperella pyogenes.
Streptococcus suis.
Moraxella sp.
Pseudomonas aeruginosa.
Discussion
We found that the bovine NALT appeared as a cauliflower-like structure in the roof of the nasopharynx, similar to the location previously described for bovine and swine NALT.37,42 A previous study that analyzed the bovine nasopharyngeal mucosa to examine NALT determined that the exact location of the NALT is not easily identifiable. 37 Our sampling method was unique, and our dissection procedure facilitated the complete excision and analysis of NALT without bones. We found that the NALT had a higher density in the rostral portion of the nasopharynx, as described previously. 37 Histologically, NALT consists of lymphoid follicles covered by stratified columnar epithelium in rodents, humans, and pigs and plays an important role in nasal mucosal immune defense.22,42,44 We suggest that bovine NALT exerts a similar immune defense function.
All pneumonic JB calves had nasopharyngitis. Bacteriologic examination revealed M. bovis, M. haemolytica, T. pyogenes, S. suis, Moraxella sp., and P. aeruginosa in nasopharyngeal swabs and lungs. These bacteria are thought to proliferate in the nasopharynx and cause secondary nasopharyngitis preceded by viral infection. 5 We detected M. bovis in nasopharyngeal swabs and lungs in 5 of 6 pneumonic calves, suggesting that M. bovis could proliferate in the nasopharynx and then migrate to the lungs via inhalation, leading to suppurative necrotizing bronchopneumonia, the classical lesion associated with M. bovis infection, which we observed in 3 cases. In one report, M. bovis was detected in 16 of 50 nasopharyngeal swabs from pneumonic JB cattle. 29 M. bovis has been also detected in JB calves from local farms in south Kyushu, the same area from which we collected our cases, that exhibited a variety of conditions associated with M. bovis infection, including otitis and pneumonia.57,58
Commensal bacteria in the URT can be opportunistic pathogens and are commonly associated with BRDC.36,46 Various virulence factors, including biofilms, capsules, adhesins, toxins, and enzymes, enhance the ability of these bacteria to colonize the lower airways, evade the immune system, resist antimicrobial treatment, cause tissue destruction, and provoke an intense inflammatory response. 46 Impairment of the host immune response due to stress and/or primary viral infection enhances the ability of commensal bacteria to colonize the deeper pulmonary tissues with the aid of virulence factors, subsequently leading to the development of BRDC. 9 We propose that virulence factors enable commensal bacteria to cause tissue damage and inflammation not only in the lower respiratory tract but also in the URT, leading to the development of nasopharyngitis.
We detected BCoV and BRSV in the NALT of pneumonic JB calves with nasopharyngitis. BCoV-positive signals were detected by ISH in sloughed epithelial cells in the NALT crypts. The detection of BCoV and BRSV in NALT suggests that these viruses could damage epithelial cells, thereby leading to nasopharyngitis. BCoV-positive signals have also been reported in the respiratory epithelium of the lower respiratory tract using the ISH test. 49
We detected BVDV by RT-PCR in 9 of 10 NALT and 7 of 10 lungs of the pneumonic JB calves. Six of these calves were unvaccinated; therefore, at least for these calves, the involvement of a TI BVDV field strain should be considered. To date, few studies have examined the prevalence of BVDV infection in JB cattle populations in the Kyushu region. In blood samples collected from 1,075 JB cattle at slaughterhouses in Miyazaki Prefecture, only 2 samples (0.18%) were positive for BVDV by antigen ELISA. 3 It is important to note that we used NALT and lung tissue samples. When comparing the results of antigen ELISA using various samples from PI animals, tissue biopsy samples, such as ear notch and tail skin, provide more reliable results than serum.13,67
Most BVDV-positive samples from pneumonic JB calves that we examined were weakly positive by RT-PCR, and all were negative by ISH. In a report in which Holstein or mixed-breed calves were infected with BVDV, RT-PCR assays detected the viral gene only on days 4–8 after BVDV infection. 14 An immunohistochemical analysis using skin biopsy material collected on days 10–14 after BVDV infection in Holstein and mixed-breed cattle revealed that many negative results could be attributed to the lower viral load in experimentally infected cattle compared to PI cattle. 43 Therefore, the weak-positive RT-PCR results that we found in many pneumonic JB calves, as well as negative ISH results, indicated that the calves were not PI. Instead, these calves may have been in the late stages of TI, in which the virus load is low. Studies have suggested that the depletion of lymphoid tissues that occurs during BVDV TI is also transient and is followed by recovery and repopulation of lymphoid follicles.31,32 The pneumonic JB calves that we examined had lymphoid hyperplasia, supporting the conclusion that these calves were in the late stages of TI. Although the BVDV load in the pneumonic JB calves was assumed to be low, replication of BVDV in the NALT may have a suppressive effect on mucosal immunity during the peak infectious viral load. BVDV reduces the phagocytic and bactericidal activities of macrophages and induces apoptosis of lymphocytes. 1 BVDV may also disrupt the harmonious interaction between the host and the normal bacteria that inhabit the airways of healthy cattle. 23
We found lymphoid follicles in NALT in both pneumonic and negative control calves. Follicular hyperplasia was seen in the pneumonic calves only, suggesting that these calves were in the late stages of transient BVDV infection. By contrast, BALT is not constitutively present in farm animals or humans but rather very dynamic in these species. 30 The presence and extent of BALT are dependent on age and microbial stimulation. 30 In pigs, BALT hyperplasia is clearly noticeable in bronchointerstitial pneumonia associated with Mesomycoplasma (Mycoplasma) hyopneumoniae infection. 18 Conversely, M. bovis infection in cattle produces a classical lesion characterized by suppurative necrotizing bronchopneumonia with foci of caseous necrosis, 45 which we observed consistently. We did not observe BALT hyperplasia in pneumonic calves except in calf 1.
We detected BVDV in the NALT of PI calves using RT-PCR, and BVDV-positive signals were observed by ISH at the center of NALT lymphoid follicles, which suggests that BVDV might replicate within macrophages and lymphocytes within the NALT. Previous studies indicated that PI calves have BVDV antigens in a variety of tissues, particularly lymphoid tissue, but these antigens are not associated with lesions as the immune system does not recognize the virus as foreign.31,32 An immunohistochemical study found BVDV antigens in macrophages and lymphocytes within ileal Peyer patch lymphoid follicles of PI dairy calves. 32 The lungs of all pneumonic JB calves had various forms of bronchopneumonia, including suppurative, suppurative necrotizing, fibrinosuppurative, fibrinous necrotizing, and bronchointerstitial. Bronchopneumonia is a characteristic lesion of bacterial infection, including by M. bovis. 19 In addition, the bacteriologic examination of 6 pneumonic JB calves revealed the same bacteria in nasopharyngeal swabs and lung samples from these calves. Our findings suggest that bacteria proliferating in the nasopharynx infect the lungs via inhalation, ultimately causing pneumonia. Therefore, a plausible hypothesis is that BVDV TI causes local immunosuppression within the NALT, and BCoV and BRSV damage NALT epithelial cells, which allows the proliferation of pathogenic bacteria that then are transferred from the NALT to the lungs via inhalation, causing pneumonia.
We detected BVDV, BCoV, and BRSV in the NALT and lungs of almost all pneumonic JB calves, suggesting that NALT is a prime location for virus replication. These viruses may also be associated with the development of interstitial pneumonia, which we observed in 4 cases. Rarely, we detected viruses (BVDV in 2 cases, BCoV in 1 case, and BRSV in 1 case) in the NALT but not in the lungs, which could be attributed to the sampling area of the lung, low virus load in the lung, or the stage of infection at the time of sampling. However, the detection of BRDC-related viruses and bacteria in both the NALT and lungs of pneumonic calves emphasizes the role of NALT in the pathogenesis of BRDC-related pathogens. We retrieved no reports describing the role of bovine NALT in the development of BRDC, in a search using Google, PubMed, CAB Direct, Web of Science, and Scopus, with the search terms “cattle” or “bovine”, “NALT”, and “respiratory viruses” or “upper respiratory tract lesions”, suggesting that the pathogenesis of BRDC-related pathogens has not been discussed in the bovine NALT. Our findings suggest that BVDV TI in the NALT could lead to local immunosuppression; BCoV and BRSV infection in the NALT are thought to damage NALT epithelial cells. Consequently, local immunosuppression and disruption of the epithelial barrier in the NALT facilitates the colonization of deeper pulmonary tissues by bacteria that normally inhabit the nasopharynx, leading to the development of BRDC.
Supplemental Material
sj-pdf-1-vdi-10.1177_10406387251318415 – Supplemental material for Detection of bovine respiratory disease complex–related pathogens in nasopharynx-associated lymphoid tissue
Supplemental material, sj-pdf-1-vdi-10.1177_10406387251318415 for Detection of bovine respiratory disease complex–related pathogens in nasopharynx-associated lymphoid tissue by Asmaa A. Hegazy, Mutsumi Nakai, Naoyuki Fuke, Amaal E. Hussein, Hiroaki Kondo and Takuya Hirai in Journal of Veterinary Diagnostic Investigation
Footnotes
Acknowledgements
We thank the Ministry of Higher Education of the Arab Republic of Egypt for providing a scholarship to Asmaa A. Hegazy.
Declaration of conflicting interests
The authors declared no conflicts of interest with respect to the research, authorship, or publication of this article.
Funding
Our study was funded by a research grant from Itokinen-Zaidan (108, Takuya Hirai). The funder had no role in the study design, data collection, analysis, interpretation, or manuscript preparation.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.