
Abstract
Increased doublecortin (DCX) immunolabeling at the tumor margins has been associated with tumor infiltration in human glioma and canine anaplastic meningioma. No association between DCX immunolabeling and glioma infiltration has been reported in dogs, to our knowledge. Here we compare the DCX immunolabeling in 14 diffusely infiltrating gliomas (gliomatosis cerebri) and 14 nodular gliomas with distinct degrees of tumor infiltration. Cytoplasmic DCX immunolabeling was classified according to intensity (weak, moderate, strong), distribution (1 = <30% immunolabeling, 2 = 30–70% immunolabeling, 3 = >70% immunolabeling), and location within the neoplasm (random or at tumor margins). Immunolabeling was detected in 6 of 14 (43%) diffusely infiltrating gliomas and 8 of 14 (57%) nodular gliomas. Diffusely infiltrating gliomas had moderate and random immunolabeling, with distribution scores of 1 (4 cases) or 2 (2 cases). Nodular gliomas had strong (6 cases) or moderate (2 cases) immunolabeling, with distribution scores of 1 (3 cases), 2 (3 cases), and 3 (2 cases), and random (6 cases) and/or marginal (3 cases) immunolabeling. Increased DCX immunolabeling within neoplastic cells palisading around necrosis occurred in 4 nodular gliomas. DCX immunolabeling was not increased at the margins of diffusely infiltrating gliomas, indicating that DCX should not be used as an immunomarker for glioma infiltration in dogs.
Gliomas are the most commonly diagnosed primary intraparenchymal CNS neoplasms of dogs. 7 Most gliomas are characterized as distinct masses with variable degrees of local infiltration into the adjacent neuroparenchyma. 9 However, rare subsets of gliomas (referred to as diffusely infiltrating gliomas or gliomatosis cerebri) infiltrate widely into the neuroparenchyma without causing distortion to its overall architecture. 12 Although highly invasive, diffusely infiltrating gliomas are morphologically and immunohistochemically identical to the more commonly diagnosed nodular gliomas. 12 In fact, it is likely that diffusely infiltrating gliomas represent a growth pattern of canine glioma rather than a separate entity (gliomatosis cerebri), as has been the case in human neuropathology.12,14
Doublecortin (DCX) is a microtubule-associated protein that plays an essential role in neuroblast migration in cerebrocortical development during embryonal life, but its expression has been also detected in subsets of adult subependymal and hippocampal neurons.1,4 Thus, DCX immunohistochemistry (IHC) has been utilized to detect neuronal progenitor cells and neuronal differentiation in human, canine, and feline gliomas.1,3,5,11 In addition, DCX immunolabeling has been advocated as an indicator of tumor infiltration, which is supported by the increased DCX expression and immunolabeling at the margins of invasive human CNS neoplasms, including gliomas. 1 A similar DCX immunolabeling pattern has been reported for anaplastic canine meningiomas and feline gliomas.2,6 In dogs, tumoral DCX immunolabeling has been reported regardless of glioma subtype (oligodendroglioma or astrocytoma) or grade, but, to our knowledge, no investigations have assessed the DCX immunolabeling pattern in gliomas with distinct degrees of infiltration into the neuroparenchyma. 3
We hypothesized that DCX immunolabeling should be more intense at the margins of invasive gliomas (such as diffusely infiltrating gliomas) compared to less infiltrative gliomas (compact or nodular gliomas). Here we compare the DCX immunolabeling profile in 28 canine gliomas with distinct degrees of infiltration into the neuroparenchyma.
Two sets of canine glioma were selected from the autopsy archives at the Athens Veterinary Diagnostic Laboratory, Department of Pathology, College of Veterinary Medicine, University of Georgia (Athens, GA, USA); Department of Anatomic Pathology, Schwarzman Animal Medical Center (New York, NY, USA); Department of Biomedical Sciences, Section of Anatomic Pathology, College of Veterinary Medicine, Cornell University (Ithaca, NY, USA); Department of Veterinary Pathobiology, College of Veterinary Medicine & Biomedical Sciences, Texas A&M University (College Station, TX, USA); and Department of Microbiology, Immunology, and Pathology, College of Veterinary Medicine and Biomedical Sciences, Colorado State University (Ft. Collins, CO, USA).
We selected and included 28 cases in our study. The first case set consisted of 14 cases of diffusely infiltrating glioma (gliomatosis cerebri) that have been published as part of a different investigation and were utilized because of their highly infiltrative nature. 12 The second case set consisted of 14 gliomas characterized by a compact and often nodular growth pattern with distinct degrees of infiltration. All tumors were examined histologically and graded according to a canine glioma diagnostic and classification system. 9 Although not predictive of tumor grade, tumor infiltration into the adjacent neuroparenchyma was recorded. For the diffusely infiltrating gliomas, tumor infiltration was characterized as diffuse by definition and consisted of widespread neoplastic cell infiltration in >2 cerebral lobes (with or without involvement of deep gray matter structures) or infiltration of deep gray matter structures, brainstem, and cerebellum. 12 For the nodular gliomas, tumor infiltration was recorded as absent, focal (compact tumor with focal or multiple areas of infiltration), or diffuse (compact tumor with diffuse areas of infiltration into the adjacent neuroparenchyma). 9 All tumors were subjected to IHC for oligodendrocyte transcription factor 2 (OLIG2) and glial fibrillary acidic protein (GFAP) for diagnostic confirmation, and subsequently to IHC for DCX (Table 1). 12 OLIG2, GFAP, and DCX immunolabeling was recorded as positive or negative. When positive, DCX immunolabeling was assessed according to intensity (weak, moderate, or strong), distribution (1 = <30% of neoplastic cell immunolabeling, 2 = 30–70% immunolabeling, 3 = >70% immunolabeling), and predominant location within the neoplasm (random or at tumor margins). 2
Details of immunohistochemical stains performed on sections of canine glioma.
In all cases, antigen retrieval was by use of citrate solution 10× (BioGenex), 1:10 dilution, 15 min at 110°C; antibody dilution was 1:4,000, the chromogen used was diaminobenzidine, performed in an autostainer (Nemesis 3600; Biocare).
DCX = doublecortin; GFAP = glial fibrillary acidic protein; NA = not applicable; OLIG2 = oligodendrocyte transcription factor 2.
All gliomas were diagnosed and graded using tumor morphology aided by IHC for OLIG2 and GFAP (Table 2). 9 Diffusely infiltrating gliomas consisted of low-grade astrocytomas (4 of 14; 28%), high-grade astrocytomas (4 of 14; 28%), high-grade undefined gliomas (3 of 14; 21%), low-grade oligodendrogliomas (2 of 14; 14%), and high-grade oligodendroglioma (1 of 14; 7%). Nodular gliomas consisted of high-grade oligodendrogliomas (7 of 14; 50%), low-grade oligodendrogliomas (2 of 14; 14%), low-grade astrocytomas (2 of 14; 14%), high-grade astrocytomas (2 of 14; 14%), and high-grade undefined glioma (1 of 14; 7%). Tumor infiltration was diffuse in all diffusely infiltrating gliomas and focal (7 cases), diffuse (5 cases), and absent (2 cases) in the nodular gliomas. Regardless of glioma category (diffusely infiltrating vs. nodular glioma), subtype, or grade, DCX immunolabeling (Figs. 1–7) was observed in 14 neoplasms (50% of cases), and occurred mainly in high-grade oligodendrogliomas (6 cases), followed by low-grade astrocytomas (3 cases), high-grade undefined glioma (3 cases), and high-grade astrocytomas (2 cases).
Signalment, diagnosis, tumor infiltration, and immunohistochemistry profile of 28 canine gliomas.
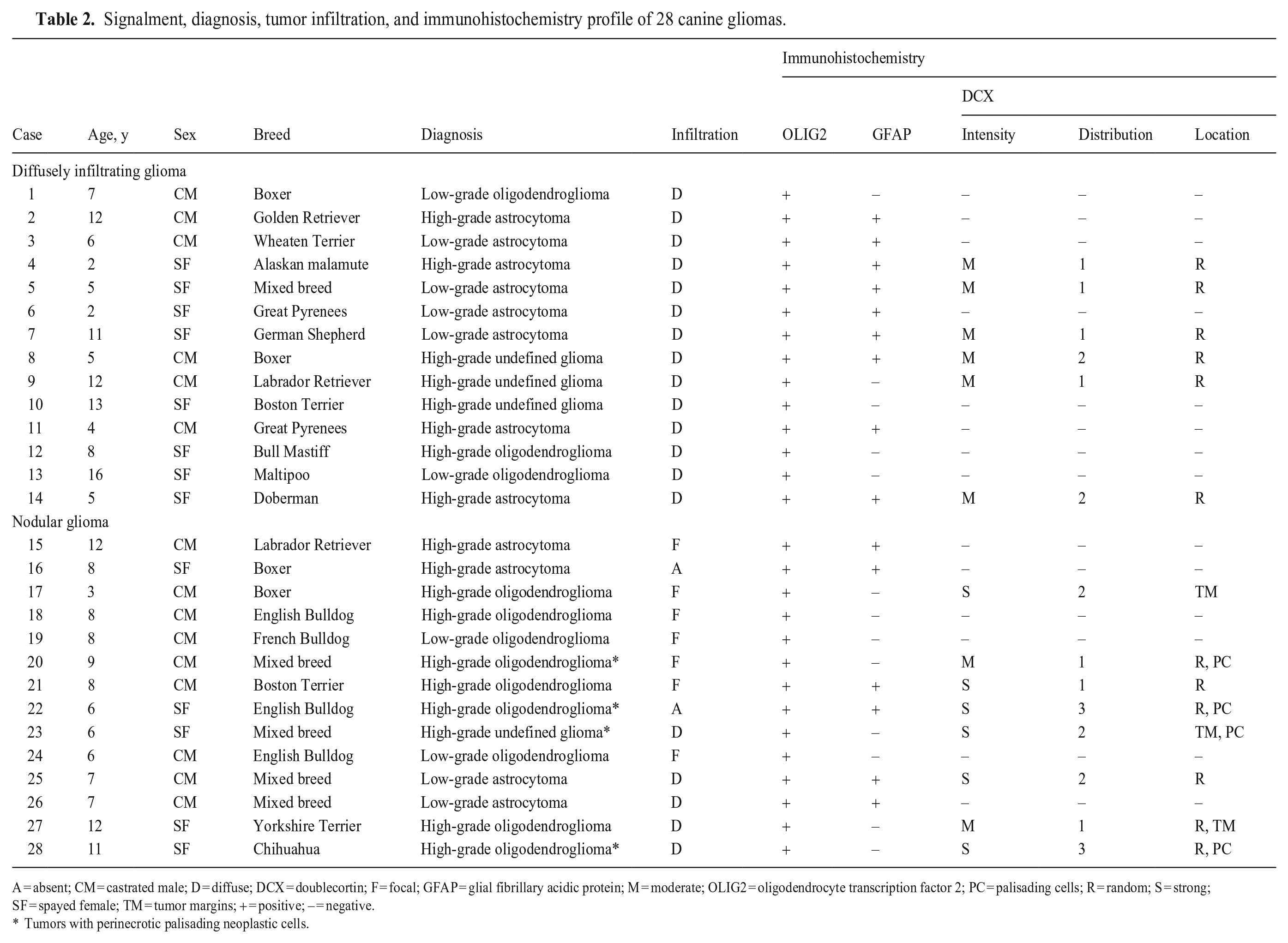
A = absent; CM = castrated male; D = diffuse; DCX = doublecortin; F = focal; GFAP = glial fibrillary acidic protein; M = moderate; OLIG2 = oligodendrocyte transcription factor 2; PC = palisading cells; R = random; S = strong; SF = spayed female; TM = tumor margins; + = positive; – = negative.
Tumors with perinecrotic palisading neoplastic cells.
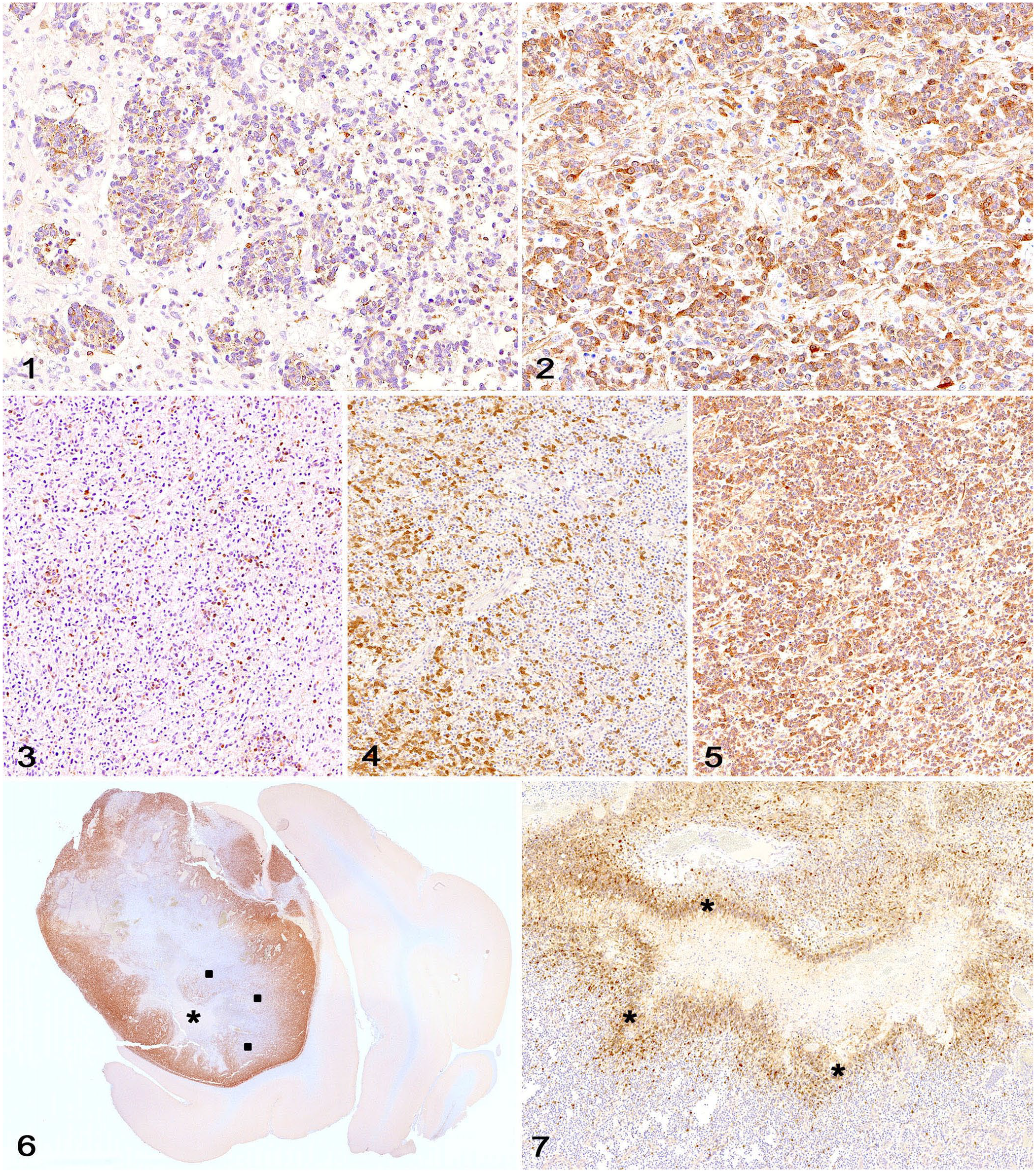
Doublecortin (DCX) immunolabeling in canine glioma.
Cytoplasmic DCX immunolabeling was observed in 6 of 14 (43%) diffusely infiltrating gliomas. The intensity of immunolabeling was moderate and random in all 6 cases, with distribution scores of 1 (4 cases) and 2 (2 cases). DCX immunolabeling was detected in 8 of 14 (57%) nodular gliomas and occurred in tumors with diffuse (4 cases), focal (3 cases), or no infiltration (1 case). The intensity of immunolabeling was strong (6 cases) or moderate (2 cases) in the nodular gliomas, with a distribution score of 1 (3 cases), 2 (3 cases), and 3 (2 cases). Immunolabeling was random (6 cases) and/or more pronounced at the margins (3 cases) in the nodular gliomas. Increased DCX immunolabeling was observed within neoplastic cells palisading around areas of necrosis in 4 of 4 nodular gliomas in which this feature was present. DCX immunolabeling was confined to neoplastic cells and normal adjacent neurons.
DCX immunolabeling has been utilized to detect neuronal differentiation by neoplastic glial cells in human, canine, and feline gliomas.1–3 Increased levels of DCX expression and immunolabeling at the margins of invasive human CNS neoplasms (including gliomas) has been also suggested as evidence that DCX-positive neoplastic cells may have a higher propensity to infiltrate the adjacent neuroparenchyma. 1 Therefore, DCX has been proposed as a potentially useful immunomarker to detect infiltrating neoplastic cells in small surgical specimens in human neuropathology. 10
In veterinary medicine, increased DCX immunolabeling has been detected in canine anaplastic meningioma, also supporting its use as a potential indicator of tumor infiltration. 6 A similar immunolabeling pattern has been observed in feline gliomas, mainly astrocytomas. 2 Given that astrocytomas tend to be more invasive than oligodendrogliomas, these findings may also support the role of DCX in tumor infiltration. However, this correlation was not as clear in other studies, which have suggested that DCX expression by glioma cells may not necessarily reflect their infiltrative potential. 10 Associations between DCX immunolabeling and tumor infiltration have not been investigated in canine gliomas, to our knowledge.3,5,8
DCX immunolabeling was observed in 50% of the canine gliomas in our case set, regardless of tumor subtype or grade, similar to reports of previous investigations.3,5,8,10 The DCX immunolabeling pattern in our diffusely infiltrating gliomas was random, with no increased staining at the tumor margins, which refutes our hypothesis that these tumors would have increased peripheral immunolabeling because of their highly infiltrative nature. Given that these gliomas are characterized by widespread tumor infiltration throughout the neuroparenchyma, our findings indicate that tumor invasion in these cases is not necessarily associated with DCX and may rely on other mechanisms. 10 Although diffusely infiltrating gliomas have been historically considered to be highly aggressive in humans, magnetic resonance imaging has revealed that their incidence may have been underestimated because many tumors have an indolent, slowly evolving behavior. 13 To date, the degree of heterogeneity and potential differences in biologic behavior among these neoplasms and other types of glioma are unknown in companion animals.
In contrast, DCX immunolabeling in our nodular gliomas was similar to that reported in canine and feline gliomas,2,3 with strong and random cytoplasmic immunolabeling in most cases. Increased staining at the tumor margins was observed in 2 high-grade oligodendrogliomas with focal and diffuse infiltration and 1 high-grade undefined glioma with diffuse infiltration. Our results confirm that neoplastic glial cells in canine gliomas can undergo neuronal differentiation, as demonstrated previously,3,7 but also reveal that DCX immunolabeling may not be a reliable indicator of tumor infiltration, as suggested by studies on human gliomas. 10
One of our findings that has not been reported in human and veterinary medicine was the increased DCX immunolabeling within neoplastic cells palisading around areas of geographic necrosis in 4 gliomas (3 high-grade nodular oligodendrogliomas and 1 high-grade nodular undefined glioma). Geographic necrosis with or without palisading cells, together with microvascular proliferation, is a key feature of high-grade gliomas. 9 Given that perinecrotic palisading cells are thought to reflect neoplastic cell migration away from an increasingly hypoxic area of tumor tissue, it is possible that DCX expression could have a role in the movement of neoplastic cells away from necrotic foci. 14 Another possibility is that the immunolabeling may have been artifactual given its location around extensive areas of necrosis. However, the cytoplasmic staining pattern was similar to that observed in the other areas of the DCX-positive neoplasms, which suggests that the perinecrotic immunolabeling was genuine.
Footnotes
Acknowledgements
We thank Nicole Young, Philip C. Dixon, and Jennifer Kempf (Histology Laboratory, Department of Pathology, College of Veterinary Medicine, University of Georgia) for their support with immunohistochemistry.
Declaration of conflicting interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors declare that they received no financial support for their research and/or authorship of this article.