
Abstract
We developed a PCR assay for the detection of Babesia odocoilei based on the 18S rRNA gene. Multiple specimens of B. odocoilei were examined, and the assay consistently produced a small specific PCR product of 306 bp. The PCR assay was also challenged with DNA from 13 other Babesia species and 2 Theileria species, originating from 10 different host species; however, nonspecific DNA amplification and multiple banding patterns were observed, and the amplicon banding patterns varied between different isolates of the same species. Sensitivity was determined to be 6.4 pg of DNA, and an estimated 0.0001% parasitism. This assay can be utilized for species-specific differential detection of B. odocoilei.
Keywords
Babesia odocoilei is an intraerythrocytic protozoan parasite, transmitted by Ixodes spp. ticks, that has been identified in cervids within Canada since 2013.13,16 B. odocoilei infection can manifest as subclinical infection, chronic debilitating disease, or acute fatal hemolytic anemia.3,6,17 Diagnosis can be achieved through visualization of organisms in peripheral blood smears, detection of antibodies using serology, and PCR testing. PCR has the advantage of high sensitivity, and the ability to precisely identify the organism, which is particularly important when there is the possibility of infection with morphologically similar organisms such as Theileria spp. or other species of Babesia.1,2,19,20
To date, B. odocoilei is the only piroplasm known to infect cervids in Canada, but this is not the case in other countries in which several different Babesia species must be considered, and coinfections are common. 9 Cervid babesiosis is considered an emerging disease in Canada. The geographic distribution in wild and captive cervids, and the role of non-cervid hosts in regional B. odocoilei epidemiology, is uncertain. 14 Surveillance data are limited, leaving many gaps in our understanding of the risk to captive and free-ranging cervids in Canada. 14 A specific test for B. odocoilei that could be used on a variety of samples, including formalin-fixed tissue, would facilitate both prospective and retrospective surveillance of this emerging pathogen in Canada.
We evaluated sensitivity and confirmed the specificity of a 306-bp PCR assay for B. odocoilei when challenged with DNA from a variety of Babesia and Theileria spp. For cervids, fresh or frozen blood, spleen, or velvet antler are generally preferred for PCR testing, with follow-up sequencing confirming the specificity of the test. 10 However, there are many circumstances in which fresh or frozen tissue may not be readily available and testing is limited to formalin-fixed samples. It has been well established that formalin causes chemical modifications and crosslinking of proteins, thereby reducing amplicon length or rendering samples unsuitable for PCR. 12 With this in mind, a set of primers has been designed to only anneal with the B. odocoilei sequence, providing specificity, as well as producing a smaller product (306 bp) more suitable to the reduced amplicon length often associated with formalin fixation. 10 Achieving specificity of this assay was important because the ability to confirm Babesia species through sequencing was lost given the smaller product size and loss of the annealing site during trimming and editing. In a previous study, this PCR was shown to differentiate known B. odocoilei–positive and –negative samples, and was found to be effective in detecting B. odocoilei in tissues stored frozen at −20°C for 1 y, and in 10% neutral-buffered formalin for up to 6 mo. 10
DNA samples (extracted from mammalian blood [n = 35] and ticks [n = 10]) and reference strains (n = 3) of a variety of Babesia and Theileria spp. were procured from the Department of Parasitology, Institute of Genetics and Microbiology, University of Wrocław (Wrocław, Poland); the Department of Veterinary Pathobiology, Texas Veterinary Medical Center, Texas A&M University (College Station, TX, USA); and the authors’ laboratories to assess the specificity of the PCR assay (Table 1).
Host, geographic origin, method of identification, GenBank accession, and result of our PCR assay of 48 samples of Babesia and Babesia-like organisms isolated and purified from mammalian blood (n = 35), tick (n = 10) samples, or obtained as reference strains (n = 3).
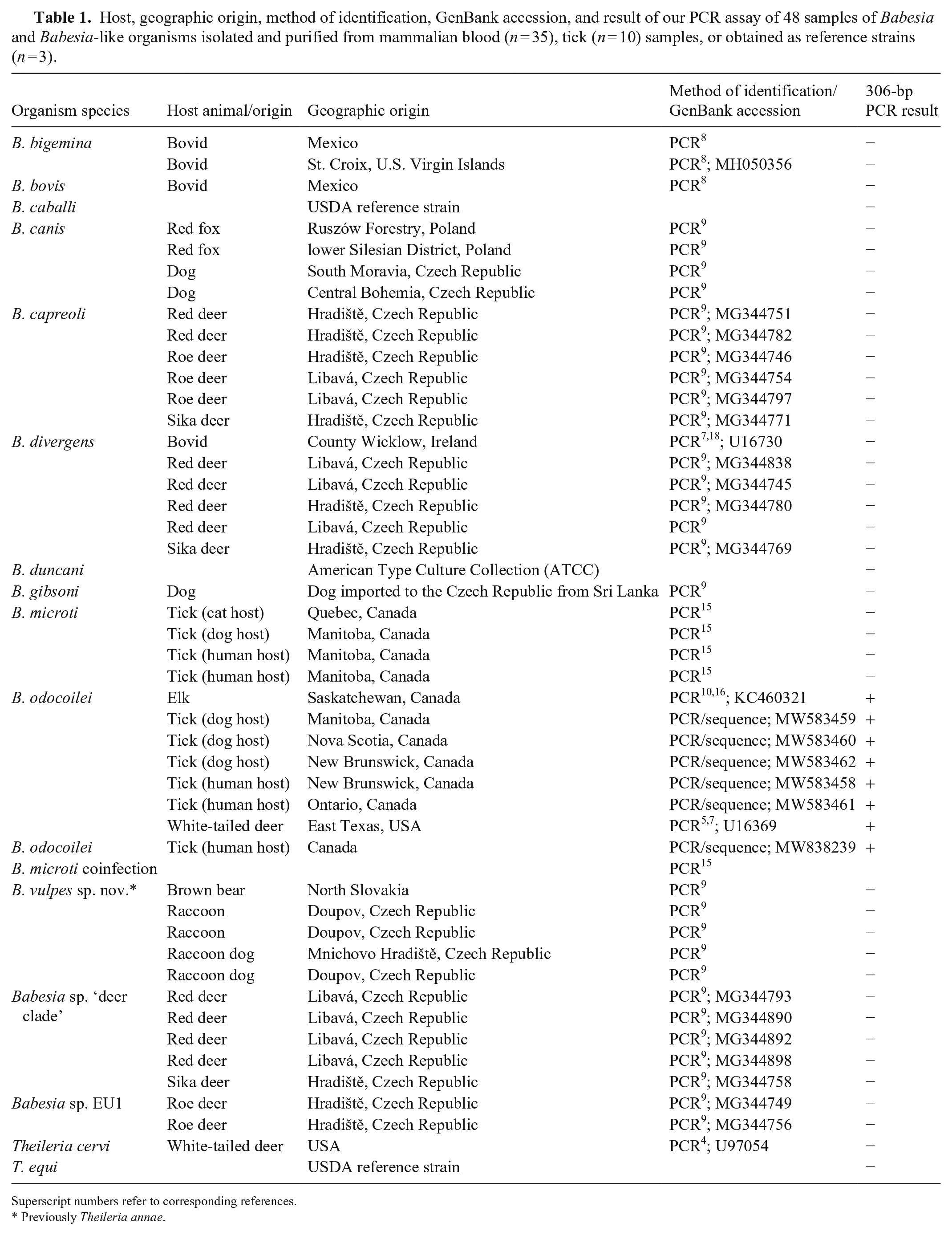
Superscript numbers refer to corresponding references.
Previously Theileria annae.
Each of the 48 samples, used to assess specificity, was analyzed in duplicate using the 306-bp PCR assay. Pellets were resuspended in 30 µL of Tris-EDTA buffer. Prior to each PCR, adequate DNA concentration (50–1,000 ng/µL) was confirmed (NanoDrop spectrophotometer; Thermo Fisher). The primers used for this PCR (Millipore Sigma), with nucleotide sequence listed from 5′ to 3′, were Bab CF (AGGCAGCAACGGGTAACG) and Bab 306R (AATACGGTGACGCAGAAA). As described previously, the primers were designed (Geneious prime; Biomatters) based on the B. odocoilei DNA sequence (GenBank U16369), with the forward primer (Bab CF) starting at position 318, and the reverse primer (Bab 306R) ending at position 623. 10 The annealing site of the reverse primer was in the region of the DNA sequence where the 32-position difference between B. odocoilei and B. divergens was located.7,10 The PCR assay was optimized using a final concentration of 4 mM MgCl2 and 1.25 units of Taq polymerase at an annealing temperature of 58.8°C. Amplification was performed in a 50-µL PCR reaction containing 33.2 µL of water, 5 µL of 10 × PCR buffer, 4 µL of 50 mM MgCl2, 2.5 µL of Bab CF (20 µM), 2.5 µL of Bab 306R (20 µM), 0.5 µL of 25 µM dNTP, 0.25 µL of Accu-start Taq (5 U/µL; Quantabio), and 2 µL of DNA. A Mastercycler pro (Eppendorf) was used, and the thermocycler parameters were as follows: initial denaturation for 3 min at 96°C, then 40 cycles at 94°C for 10 s, 58.8°C for 30 s, and 72°C for 30 s. The final extension was at 72°C for 10 min, and hold at 10°C.
Seven B. odocoilei samples consistently produced a single band of the expected size (306 bp). Only nonspecific binding occurred when the PCR assay was challenged with 40 samples comprising 12 different Babesia spp. and 2 Theileria spp., with no dominant bands present at the 306-bp mark. One sample originated from a tick coinfected with B. odocoilei and B. microti. The sample showed the expected dominant band of 306 bp, which was confirmed as B. odocoilei by sequencing, and a second pale band of larger size. Interestingly, this finding indicates that the primers preferentially bind to the specific binding sites on the B. odocoilei template DNA in samples collected from ticks or animals coinfected with B. microti. The second band likely reflected nonspecific binding, as expected with B. microti (Fig. 1). With the exception of B. odocoilei, there was no consistency in the banding pattern for Babesia spp. that had >1 sample available (8 species, 34 samples), reinforcing the nonspecific nature of the primer binding in these cases.
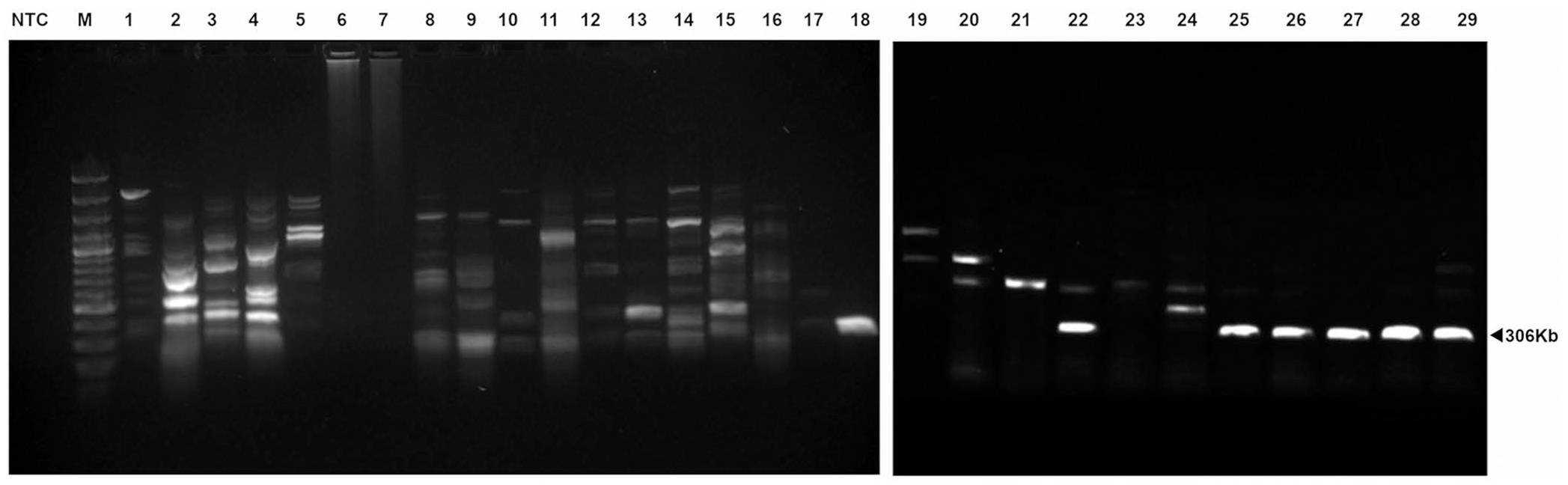
Gel electrophoresis of our PCR assay products using total DNA extracted from samples infected with different species of Babesia. In the case of B. odocoilei, only a single dominant band (306 bp) was amplified consistently. A secondary larger faint band is observed in samples coinfected with B. odocoilei and B. microti. In some cases, DNA from samples infected with B. microti produce a single band of ~650 bp, but this is not consistent from sample to sample. PCR of template DNA from other Babesia spp. resulted in multiple dominant bands of equal intensity and variable size, as well as other multiple faint bands. Lanes: M = marker; NTC = no template control; 1 = B. canis; 2–5, 15 = Babesia sp. “deer clade”; 6–9, 16 = B. vulpes; 10–12 = B. capreoli; 13, 14 = Babesia sp. EU1; 17 = B. caballi; 18, 25–29 = B. odocoilei; 19 = B. duncani; 20, 21, 23, 24 = B. microti; 22 = coinfection with B. odocoilei and B. microti.
To assess the limit of detection for the PCR assay, DNA was extracted from EDTA-blood from an elk infected with B. odocoilei (GenBank KC460321) by digestion using a lysis buffer containing 100 mM NaCl, 500 mM Tris, and 10% sodium dodecyl sulfate. Proteinase K (50 mg) was added, the samples were incubated overnight at 56°C, protein was extracted with phenol–chloroform, and DNA was precipitated with ethanol. 11 Pellets were resuspended in 30 µL of Tris-EDTA buffer. The DNA concentration of the resulting sample was measured (104 ng/µL; NanoDrop spectrophotometer, Thermo Fisher), and then serially diluted with Tris-EDTA buffer. A 2-µL volume was used, with the total amount of DNA for each dilution as follows: 20.8 ng, 4.16 ng, 0.83 ng, 0.17 ng, 32 pg, 6.4 pg, and 1.28 pg. PCR was performed in duplicate on the original sample, and each dilution, producing a single band of the expected size (306 bp) for all dilutions up to and including the sample with 6.4 pg of DNA. No band was observed for the sample with 1.28 pg DNA (Fig. 2).
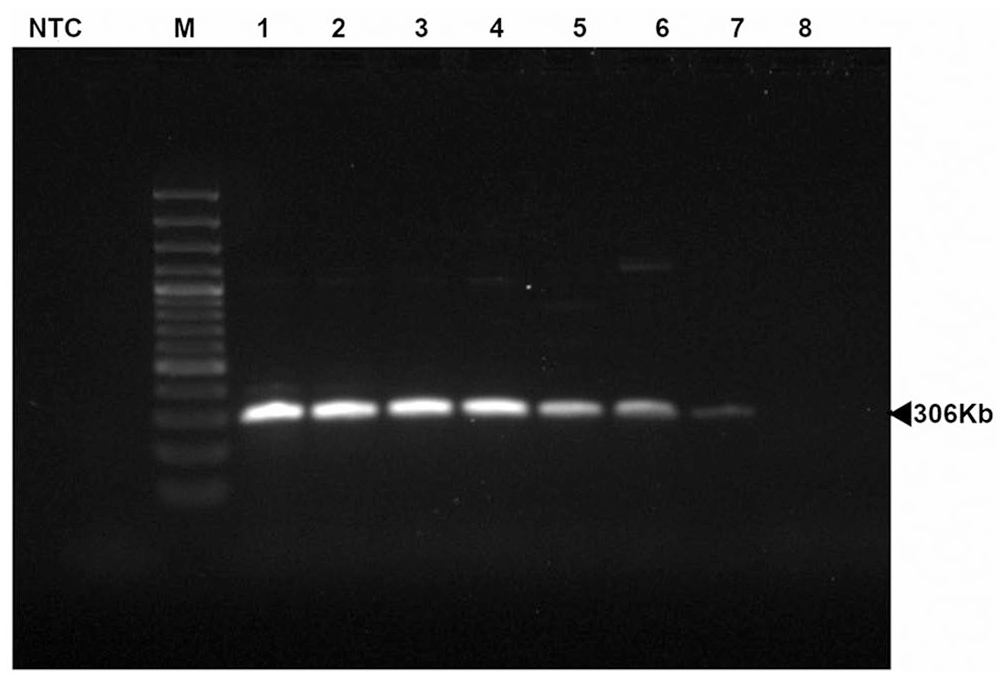
Products of our Babesia odocoilei PCR assay performed using 5-fold diluted total DNA extracted from blood samples for the detection of B. odocoilei. Lanes: M = marker; NTC = no template control; 1 = 104 ng/µL; 2 = 20.8 ng/µL; 3 = 4.16 ng/µL; 4 = 0.83 ng/µL; 5 = 0.17 ng/µL; 6 = 32 pg/µL; 7 = 6.4 pg/µL; 8 = 1.28 pg/µL.
The PCR assay that we used has been shown to successfully amplify B. odocoilei DNA in fresh-frozen tissue, as well as tissue stored in 10% neutral-buffered formalin for up to 6 mo. 10 This provides an advantage for cases where samples are limited to formalin-fixed tissues, as well as those cases in which tissue processing may be delayed, such as field studies or cases awaiting chronic wasting disease test results. In making the product size of this assay small enough to accommodate formalin-associated protein modifications, the ability to definitively confirm the Babesia spp. through sequencing was lost, making the specificity of the primers integral to the utility of this assay. When challenged previously with B. microti DNA, only nonspecific binding occurred, and there was no band present at the 306-bp mark. 10 However, given that cervids can be infected by a number of Babesia spp. (including B. capreoli, Babesia sp. EU1, B. divergens, and “deer-clade”), 9 further investigation into the specificity of this primer set was necessary.
We evaluated here the ability of this PCR to differentiate B. odocoilei from 12 other Babesia spp. (6 of which infect cervids8,9) and 2 Theileria spp. (one of which infects cervids 4 ). As mentioned, cervid babesiosis is a rare occurrence in Canada, and other infectious disease agents that cause similar clinical signs are not recognized in this geographic region. The species of Babesia and Theileria used to challenge the specificity of this PCR assay were chosen to reflect those that may be present in cervids alongside B. odocoilei in other geographic regions and to exclude cross-reactivity with other similar species regardless of their host specificity or geographic origin. Given that only B. odocoilei produced a positive result on the PCR assay, we were able to demonstrate the exclusivity of this assay. The B. odocoilei samples that we used originated from 3 different hosts (elk, tick, and white-tailed deer) from a variety of different geographic regions (USA and 6 provinces in Canada), with consistent positive results demonstrating inclusivity within this species of Babesia.
Based on our results, it is possible for additional faint, nonspecific bands to accompany the 306-bp dominant band in cases of coinfection, and in these cases further investigation into infection with additional Babesia spp. is warranted. Sensitivity was determined to be 6.4 pg of total DNA. Given that our PCR assay is not quantitative, we were unable to determine sensitivity specifically for Babesia DNA. Many publications have indicated sensitivity in terms of percentage parasitism. On evaluation of the EDTA-blood smears used in the determination of sensitivity, parasitism was 1.5% of erythrocytes. 16 The estimated percentage parasitism for the final dilution would be 0.000096%, indicating a highly sensitive PCR. This level of sensitivity is equal to that found for a nested PCR designed to amplify the small subunit rRNA gene for B. gibsoni. 2
Although PCR provides numerous advantages in the detection of infectious disease agents, sample storage conditions can limit performance. The primers that we evaluated have expanded the repertoire of sample types available for B. odocoilei testing, and the high sensitivity and specificity of the primers have made this a useful test without the need for confirmation by follow-up sequencing.
Footnotes
Acknowledgements
We thank Agnieszka Perec-Matysiak, from the Department of Parasitology, Institute of Genetics and Microbiology, University of Wrocław, for the generous donation of B. canis DNA; and Patricia Holman, from the Department of Veterinary Pathobiology, Texas Veterinary Medical Center, Texas A&M University for her generous donation of Babesia divergens, B. bigemina, B. bovis, B. caballi, B. odocoilei, Theileria cervi, and T. equi DNA.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Our research was supported financially by the Ministry of Education, Youth and Sports of the Czech Republic under project CEITEC 2020 (LQ1601)