
Abstract
A concurrent outbreak of infection by canine parvovirus 2b (CPV-2b) and Clostridium difficile producing A and/or B toxins occurred in Asian small-clawed otters (Amblonyx cinereus). The 5 clinically affected otters were 6- to 24-mo-old intact females that had severe diarrhea, dehydration, were acutely comatose, and died 1–4 d after the onset of clinical signs. Postmortem examination was performed in 3 of 7 otters. Macroscopically, the small intestine was diffusely reddened and contained red-to-brown, malodorous, watery digesta without formed feces (3 of 3). Histologic examination identified loss of enterocytes and necrosis of crypt epithelial cells. Denuded villi were often covered by mixed bacterial colonies with a predominance of gram-positive cocci to short rods in addition to larger gram-positive and -negative rods. There was also splenic lymphoid follicle depletion (2 of 3). Immunofluorescence assay revealed CPV antigen in enterocytes (2 of 3), mesenteric lymph nodes (3 of 3), and spleen (1 of 3). Immunohistochemistry revealed CPV antigen in enterocytes, lymphocytes, and dendritic cells of the Peyer patches and spleen (3 of 3), and lingual epithelial cells (1 of 2). CPV was isolated from tissues from 2 of 3 otters, and DNA sequencing identified CPV-2b for the 1 isolate tested. C. difficile producing A and/or B toxins were identified in the intestinal content by ELISA (3 of 3). To our knowledge, an outbreak of CPV-2b infection and C. difficile with clinically significant gastrointestinal disease has not been described previously in otters. The source of the viral infections remains unknown; however, these agents should be considered in otters and other mesocarnivores with similar clinical and pathologic findings.
The Asian small-clawed otter (Amblonyx cinereus) is a valuable exotic species kept in zoologic collections and as pets. In addition, Asian small-clawed otters are considered “vulnerable” on the IUCN Red List of Threatened Species.6,12,18 It is important to understand the effects that diseases can have on the health status of these animals, as well as the associated economic loss. Canine parvovirus 2 (CPV-2; family Parvoviridae, genus Protoparvovirus, species Carnivore protoparvovirus 1) causes gastrointestinal disease in several domestic and non-domestic carnivore species; the disease is of concern for mustelids, however, clinical disease has been reported only rarely.1,3,10 We describe an outbreak of CPV-2b combined with Clostridium difficile infection causing severe necrotizing enteritis in young and adult otters.
The outbreak occurred in an Asian small-clawed otter captive conservatory facility that housed 7 animals (5 females and 2 males), 6–36 mo old. Three offspring (3 mo old) were kept in a separate pen. All otters were fed whole fish and/or fillets (fish species were not identified), shrimp, and dry dog food. Five intact female otters (6–24 mo old) were affected clinically. Supportive treatment included fluid therapy that varied from 1.5 mL of dextrose in 1.5 mL of lactated Ringer solution (LRS) administrated intraperitoneally, followed by 50 mL of dextrose in 500 mL of LRS intravenously and/or subcutaneously, oxygen intubation for 2 h or until oxygen saturation reached 95%, and administration of broad-spectrum antimicrobials that included 5 mg/kg of enrofloxacin IM (100 mg/mL Baytril 2.27% injectable solution; Bayer Healthcare, Shawnee, KS). Clinical signs included bradycardia, hypoglycemia (glucose of 1.11 mmol/L [20 mg/dl] recorded only in a 6-mo-old, female otter; [reference interval: 2.8–10.1 mmol/L] 17 ), severe diarrhea to hematochezia, dehydration, ataxia, and acute coma. Despite treatment, 4 of 5 female otters died 1–4 d after the onset of clinical signs. Three of the 4 dead otters were submitted for postmortem examination.
All animals (3 of 3) were in fair nutritional condition and had reddened small intestines with red-to-brown malodorous, watery, digesta without well-formed feces (Fig. 1). Multiple tissue samples were fixed in neutral-buffered 10% formalin solution and processed routinely for: 1) routine histologic examination, 2) Gram staining, 3) CPV immunohistochemistry (IHC) using a specific rabbit polyclonal anti-parvovirus antibody (antigen retrieval: proteinase K; DAKO/Agilent, Carpinteria, CA), and 4) DNA extraction and real-time PCR (rtPCR) for C. difficile in formalin-fixed, paraffin-embedded (FFPE) intestinal tissue. In addition, selected fresh tissues underwent ancillary testing by: 1) immunofluorescence assay (IFA) for CPV and canine distemper virus (CDV) on lung, small intestine, mesenteric lymph node, and spleen; 2) virus isolation in A-72 cells and DNA sequencing of isolates of CPV-2 from the small intestine, mesenteric lymph node, and spleen; 3) aerobic culture and specific culture media for Salmonella spp. and Campylobacter spp. of fresh small intestine and mesenteric lymph node; 4) fecal flotation (parasitology) on colon and contents; and 5) C. difficile and C. perfringens toxin ELISA (Clostridium difficile Tox A/B II; TechLab, Blacksburg, VA) on intestinal contents. Nucleic acids for CPV-2 were extracted (Viral RNA kit; Qiagen, Germantown, MD) and the PCR was run (Platinum PCR super mix; Invitrogen, Carlsbad, CA). A portion of the CPV-2 VP2 gene was amplified, and a band of the appropriate size was purified and submitted for nucleotide sequence analysis. The edited sequence was aligned with CPV types 2a–c sequences to determine the genotype of the virus. CPV-2b was determined by the amino aspartic acid residue at codon 426 in VP2.
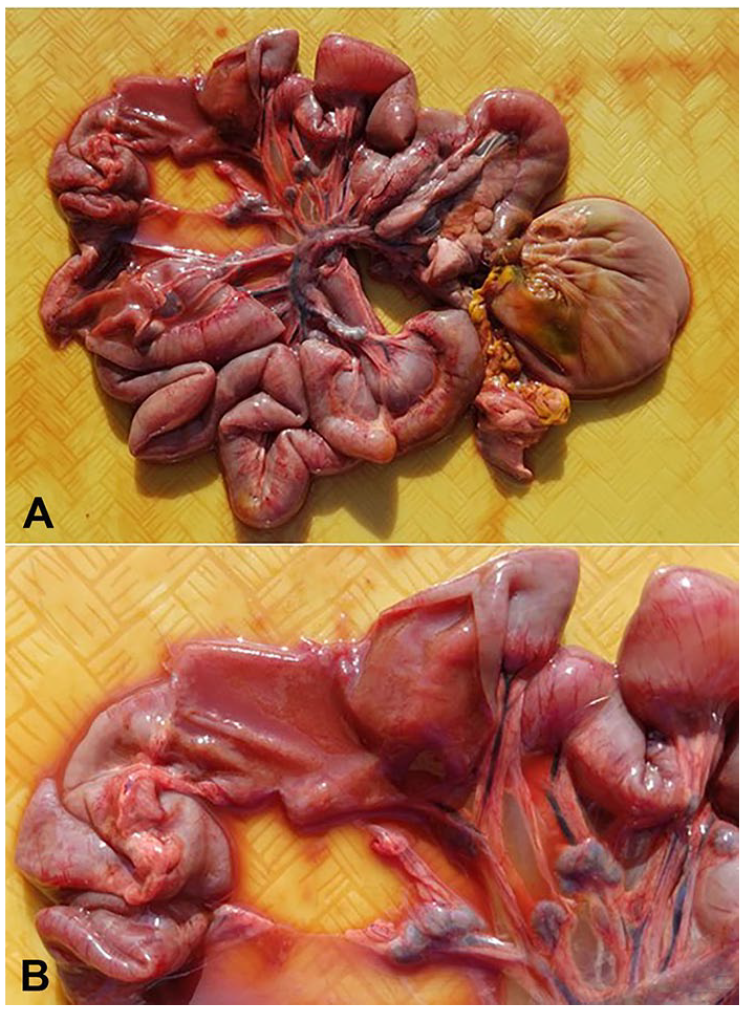
Parvoviral enteritis and Clostridium difficile infection in Asian small-clawed otters. The small intestine is diffusely distended by abundant malodorous, red, watery digesta. The mucosal and serosal surfaces are diffusely reddened. The mesenteric lymph nodes are mildly to moderately enlarged.
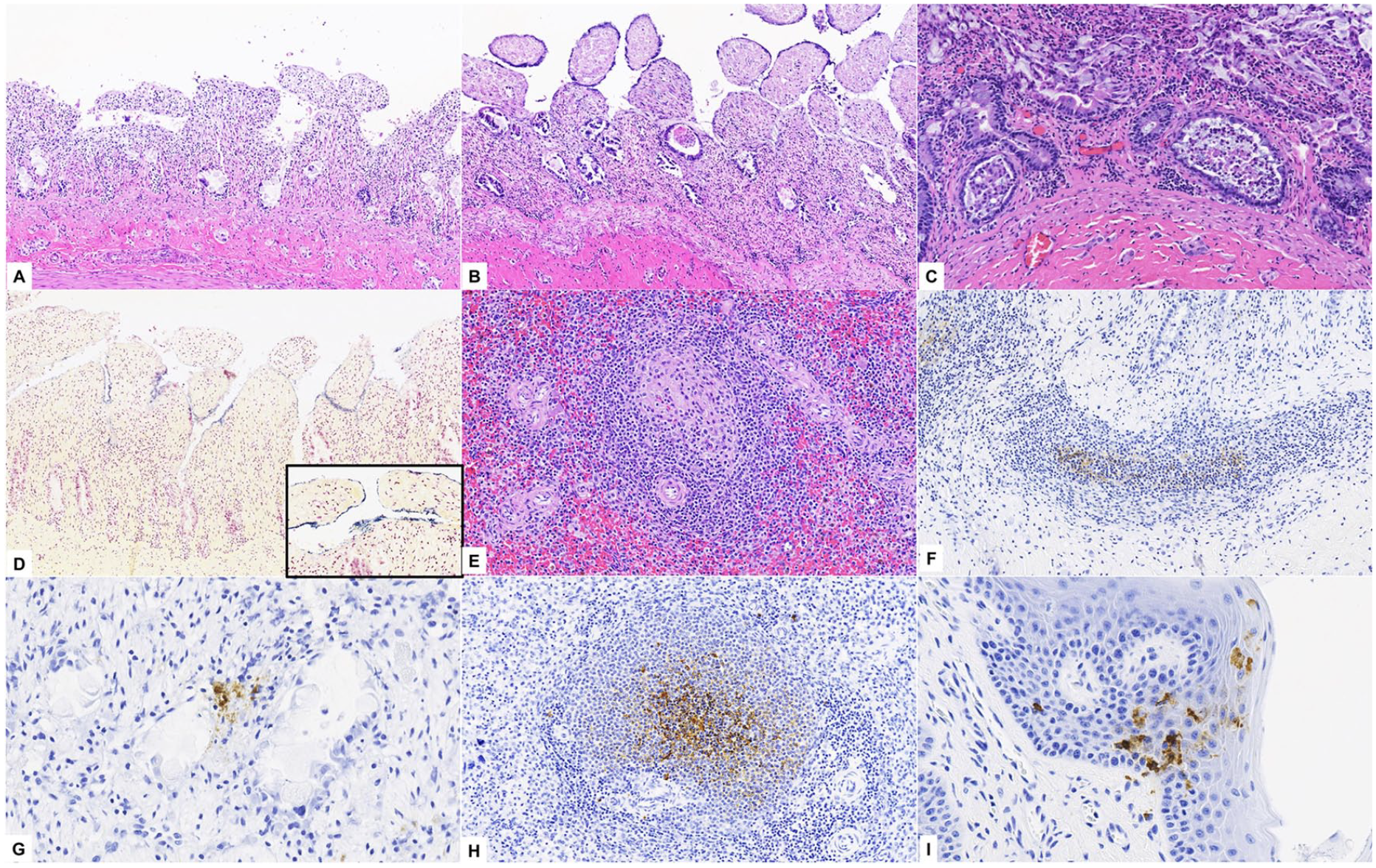
Histologically, there was frequent loss of luminal enterocytes and necrosis of crypt epithelial cells. Villi were shortened and blunted with occasional collapse of the mucosal architecture (Fig. 2A, 2B). Crypts were frequently dilated and lined by attenuated enterocytes with moderate-to-marked epithelial hyperplasia (Fig. 2C). Denuded villi were often coated by mixed bacteria with a predominance of gram-positive cocci to short rods in addition to larger gram-positive and -negative rods (Fig. 2D). Splenic follicles had lymphoid depletion (2 of 3; Fig. 2E). Lingual epithelial cells rarely had basophilic-to-amphophilic intranuclear inclusion bodies with chromatin margination.
Parvoviral enteritis and Clostridium difficile infection in Asian small-clawed otters.
Lesions noted during histologic examination were highly suggestive of an enteric form of CPV infection, and CPV was detected by IFA in the small intestine (2 of 3), mesenteric lymph nodes (3 of 3), and spleen (1 of 3). IHC revealed CPV antigen in the lymphocytes, dendritic cells of Peyer patches, and enterocytes (3 of 3; Fig. 2F, 2G), spleen (3 of 3; Fig. 2H), and lingual epithelial cells (1 of 2; Fig. 2I). CPV was isolated in cell culture from pooled samples (mesenteric lymph nodes, small intestine, and spleen) of 2 otters, and virus from 1 animal was determined to be CPV-2b by sequence analysis. In addition, ELISA demonstrated the presence of C. difficile toxins A and/or B in all tested intestinal content samples; no C. perfringens toxins were identified. Heavy growth of Escherichia coli (3 of 3), heavy-to-moderate growth of beta-hemolytic Streptococcus spp. (2 of 2), and heavy growth of Streptococcus dysgalactiae (1 of 3) were revealed by small intestine and mesenteric lymph node bacterial culture. Cultures for Salmonella spp. and Campylobacter spp. were negative. No CDV was detected by IFA in all tissues sampled (3 of 3), and no parasites or ova were identified through fecal flotation examinations (3 of 3).
In the United States, acquiring Asian small-clawed otters as an exotic pet is considered legal only in a few states. This practice should be strongly discouraged given that Asian small-clawed otters are categorized as “vulnerable” on the IUCN Red List of Threatened Species as a result of habitat loss, depletion of prey species, pollution, and exploitation.6,12,18 This species is challenging to maintain and care for as pets. In addition, diseases potentially contribute to the population decline of this otter. Studies reporting disease outbreaks in Asian small-clawed otters based on clinical, pathologic, immunohistochemical, virologic, and molecular characterization are scarce in the literature.
Only female otters were clinically affected in this outbreak. Whether or not this represents a true sex-based difference in susceptibility needs to be confirmed. The male and female offspring (3 mo old) and the adult males (36 mo old) showed no clinical signs of infection during the outbreak; the CPV infection status of these animals was not assessed by serologic or molecular testing and, thus, they may have remained free of CPV infection or were subclinically infected. In previously reported outbreaks of CPV in Asian small-clawed otters, the disease course was prolonged (25 d) 3 in comparison to our case (1–4 d), and the variant previously identified was CPV-2c rather than CPV-2b. Additionally, it was initially hypothesized that CPV-2 infection could predispose to another viral infection such as canine circovirus 1 as reported in dogs, resulting in more severe clinical disease 15 ; however, no other viral infections were noted histologically or detected by ancillary testing.
One neotropical river otter (Lontra longicaudis), considered “near-threatened” on the IUCN Red List of Threatened Species, 14 infected with CPV and incidentally parasitized by Dioctophyme renale is reported, 1 but virus isolation was not performed. The source of the viral infection in our outbreak remains undetermined, but it is suspected that other carnivores may have been the source, given that some reports suggest transmission between domestic and wild carnivores.5,11 Information on other carnivores, including wildlife, that may have been living in or around the same premises as the affected Asian small-clawed otters, was not available.
Once the diagnosis of CPV-2b infection was made, surviving and non-clinically affected otters were vaccinated with a vaccine for dogs (Merial, Athens, GA). No other gastrointestinal disease cases were observed after 4 mo of follow-up. The efficacy of vaccination was not serologically evaluated serologically; therefore, no conclusions can be drawn regarding seroconversion. However, inadequate seroconversion has been reported using killed vaccines for CPV-2 in North American river otters (Lontra canadensis).4,9
Infection with C. difficile, a gram-positive rod, can cause spontaneous disease in several animal species. 8 Antibiotic treatments should be considered as a predisposing factor because the infection can occur secondarily to dysbiosis of the intestinal microbiome. Disease caused by C. difficile is mediated by essential virulence factor toxins A and B. Therefore, the presence of these toxins in the intestinal contents is considered confirmatory for C. difficile infection in several domestic animals.8,13,16 The definitive role of C. difficile in cattle, dogs, and cats is uncertain. 16 Several bacteria observed histologically in our outbreak were suggestive of Clostridium spp., which were additionally highlighted by Gram stain. ELISA results confirmed the production of enterotoxin (toxins A and B) and cytotoxin (toxin B) by C. difficile. Accordingly, we speculate that the presence of toxins A and B could also be considered confirmatory for C. difficile infection in otters. However, further research is needed in order to support this hypothesis, as well as to determine the ability of these toxins to cause disease in this animal species. Therefore, whether the antibiotic treatment, CPV-2 infection, or a combination thereof contributed to the infection with multiple strains of C. difficile remains unclear.
Beta-hemolytic streptococci have been isolated from southern sea otters (Enhydra lutris nereis) stranded off the California coast. 7 In marine mammals, infection with these bacteria can be associated with pneumonia, sepsis, or opportunistic infections. The otters that we examined had either growth of beta-hemolytic Streptococcus spp. or S. dysgalactiae on bacterial culture of the small intestine and mesenteric lymph node, and we considered these bacteria to be most likely opportunistic invaders secondary to intestinal necrosis caused by the CPV-2b viral infection.
Wild North American river otters have tested serologically positive for CPV,4,9 CDV, 9 canine herpesvirus 1, 9 and Toxoplasma gondii. 2 Additionally, antibodies to Leptospira interrogans using the microscopic agglutination test have also been detected in free-ranging river otters. 2 Close proximity to humans, domestic animals, and rodent populations could facilitate river otter exposure to these pathogens. 2 We did not detect CDV infection in our Asian small-clawed otters. Nevertheless, proximity to other domestic animals on the property, including dogs, may have served as a source of exposure to CPV-2b.5,9,11
CPV-2b infection should be included on the list of differential diagnoses in cases of intestinal disease in mustelids. Parvovirus-induced disease could predispose to secondary infections as observed in our case, potentially increasing mortality rates.
Footnotes
Acknowledgements
We thank Robert P. Poston, Nicole W. Hazard, and Shiliang Liu (Louisiana Animal Disease Diagnostic Laboratory) for performing the IFA, ELISA, and bacterial culture, respectively, and Randall Renshaw (Animal Health Diagnostic Center, Cornell) for the CPV sequence analysis. We also thank Drs. Francisco (Paco) A. Uzal and Mauricio Navarro Badilla (California Animal Health and Food Safety Laboratory–San Bernardino branch, University of California, CA) and Drs. Nicola Pusterla and Samantha Barnum (Department of Medicine and Epidemiology, University of California, Davis, CA) for attempting a C. difficile PCR on FFPE intestinal tissue.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors declared that they received no financial support for their research and/or authorship of this article.