
Abstract
An 8-y-old, intact female degu (Octodon degus) was presented with a slow-growing mass on the tail tip. The mass was completely removed by partial caudectomy. Histologically, the last coccygeal vertebra was replaced by a lobulated neoplasm composed of large clear polygonal cells embedded in a myxoid alcian blue–positive matrix with highly vacuolated cytoplasm (physaliferous cells) and intracytoplasmic periodic acid-Schiff–positive granules. The neoplasm exhibited the morphologic features of a “classic” chordoma of humans, which is 1 of 3 distinct chordoma subtypes. Immunohistochemistry revealed dual expression of cytokeratin AE1/AE3 and vimentin, consistent with a diagnosis of chordoma. Chordomas are uncommon slow-growing neoplasms in humans and animals, arising from notochordal remnants. Depending on their subtype and location, they can have a high local recurrence rate and metastatic risk. Chordoma should be included in the differential diagnosis of a soft tissue mass on the tail of a degu, similar to the clinical situation in ferrets.
Degus (Octodon degus) are becoming increasingly popular as pet rodents, but little is known about their diseases. Clinical reports and prevalence studies of neoplasms are sporadic in the veterinary literature. A case series of 189 degus 15 recorded only 7 tumors. A 2011 retrospective study of the clinical records of 300 degus reported 6 cases of neoplasia. 10 Two chordomas have been reported, as a brief communication 12 or conference abstract (Zadravec M, et al. Spontaneous tumours and non-neoplastic proliferative lesions in pet degus. Proc 3rd ICARE Conf; March 2017; Venice, Italy).
An 8-y-old, 202 g, intact female degu (Octodon degus) was presented to the Alfort University Veterinary Hospital (Maisons-Alfort, France) with an ~1-cm mass at the tip of the tail that had been present for several months and was slowly enlarging. The mass was firm, round, non-painful, and well-defined (Supplementary Figs. 1, 2). No other abnormalities were observed on physical examination, and vital signs were within normal reference ranges.
A fine-needle aspirate of the mass had few cells and was inconclusive, but ruled out a cyst or an abscess. Whole-body radiographs (ventrodorsal and laterolateral views) were performed under general anesthesia, mask-induced and maintained with isoflurane (1–3%, Iso-Vet, 1,000 mg/g, Piramal Healthcare, Morpeth, United Kingdom). Radiographs revealed lysis of the last caudal vertebra (Supplementary Fig. 2). No pulmonary metastases could be detected.
Surgical removal of the mass was elected as treatment with the owner’s informed consent. Given the lack of soft tissue around the mass and to ensure safe margins, a partial tail amputation was performed. Recovery was uneventful, and the degu was discharged 6 h after surgery. At the 7-d follow up after surgery, the tail was healing normally, and no complications were observed.
The amputated tail was fixed in 10% neutral-buffered formalin. Cut sections of the tail revealed a white-to-brown multilobulated mass (Fig. 1), 11 × 4 × 3.5 mm, growing along the last 3 caudal vertebrae and replacing the last vertebral body. The center of the mass contained a 5 × 2 × 2 mm necrotic cavity. The mass was processed routinely, and 4-µm thick tissue sections were stained with hematoxylin–eosin–saffron (HES), periodic acid–Schiff (PAS), and alcian blue (pH 2.5).
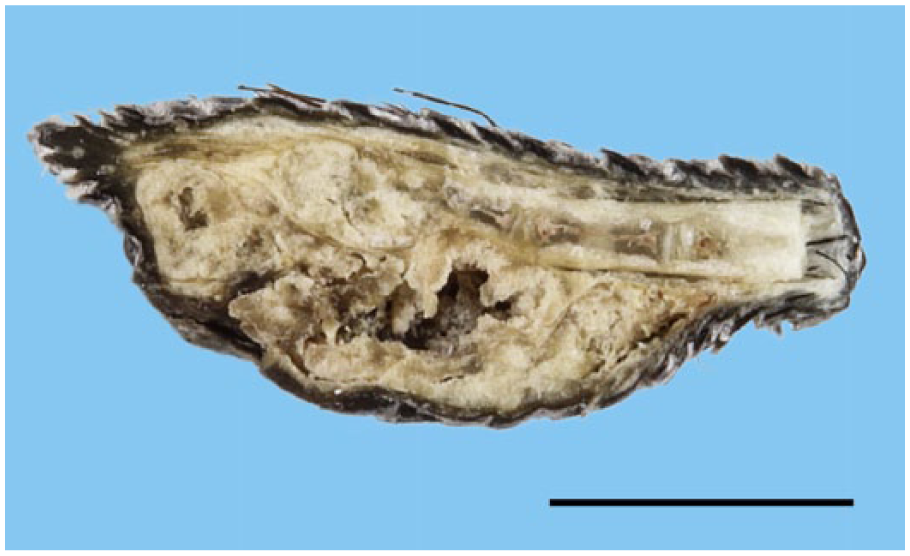
Longitudinal section of the amputated tail of a degu (Octodon degus) showing a white-to-brown multilobulated, cavitated mass expanding beneath and along the last 3 caudal vertebrae and replacing the last vertebra. Formalin-fixed specimen. Bar = 0.5 cm.
Immunohistochemistry was performed on serial sections using an automated slide stainer (Benchmark XT, Ventana Medical Systems, Tucson, AZ). Primary antibodies were specific for cytokeratins AE1/AE3 (diluted 1:100; Monoclonal rabbit anti-human, clone AE1/AE3, Dako, Ely, UK) and vimentin (prediluted; Monoclonal mouse anti-human, clone V9, Roche, Ely, UK). After antigen retrieval with Cell Conditioning CC1 solution (Ventana Medical Systems) at pH 8, primary antibodies were applied followed by a secondary biotinylated antibody; slides were then labeled by the streptavidin–biotin–peroxidase procedure, and immunoreaction was revealed with 3,3’-diaminobenzidine substrate using a commercial kit (iView DAB, Ventana Medical Systems). Sections were counterstained with hematoxylin.
Histologic examination of serial sections showed a well-circumscribed, non-encapsulated tumor replacing the last caudal vertebra, with subcutis and dermal involvement. The tumor was composed of ~40-µm polygonal cells arranged in a multilobular pattern and supported by a fine fibrovascular stroma. The neoplastic cells had distinct cell borders with up to 15 variably sized intracytoplasmic clear vacuoles (physaliferous cells; Fig. 2). These vacuoles frequently had inconspicuous PAS-positive granules (Fig. 3). Nuclei were located eccentrically, ovoid, and vesicular with a single nucleolus. There was moderate anisokaryosis and anisocytosis. Mitoses were rare (0 to 1 per ten 400× fields). Neoplastic cells were within a myxoid matrix that stained positively with alcian blue stain (Fig. 4) resembling chondroid matrix. Scattered foci of coagulative necrosis and hemorrhage were present in the mass. No lymphatic or vascular invasion was seen. The cells exhibited strong cytoplasmic immunopositivity for cytokeratin (Fig. 5) and vimentin (Fig. 6). The histologic and immunohistochemical diagnosis was chordoma.
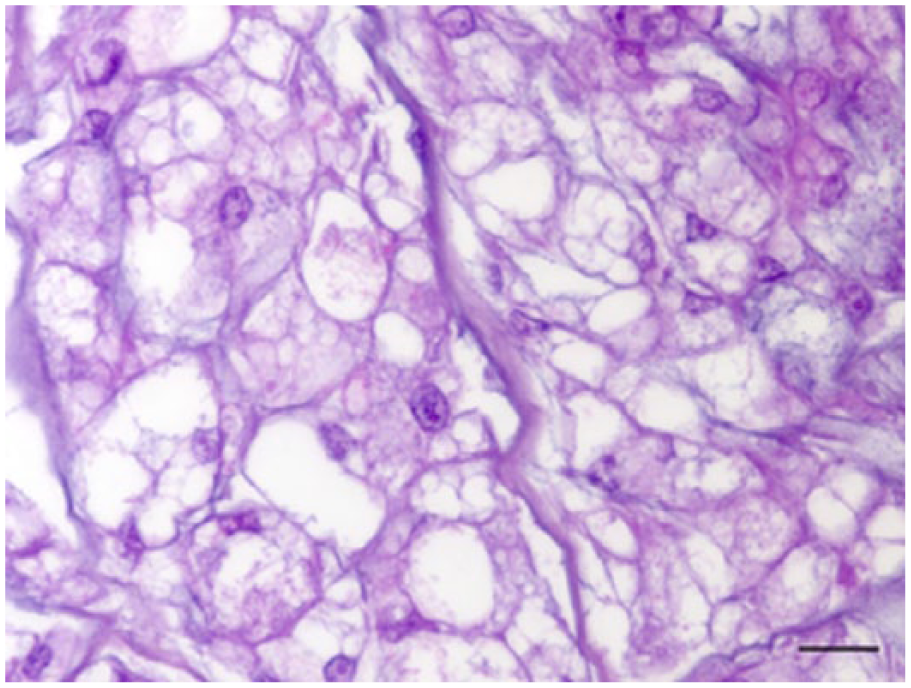
Histologic features of the coccygeal mass in a degu. Physaliferous cells display distinct cell borders with up to 15 variably sized intracytoplasmic clear vacuoles. Hematoxylin–eosin–saffron. Bar = 20 μm.
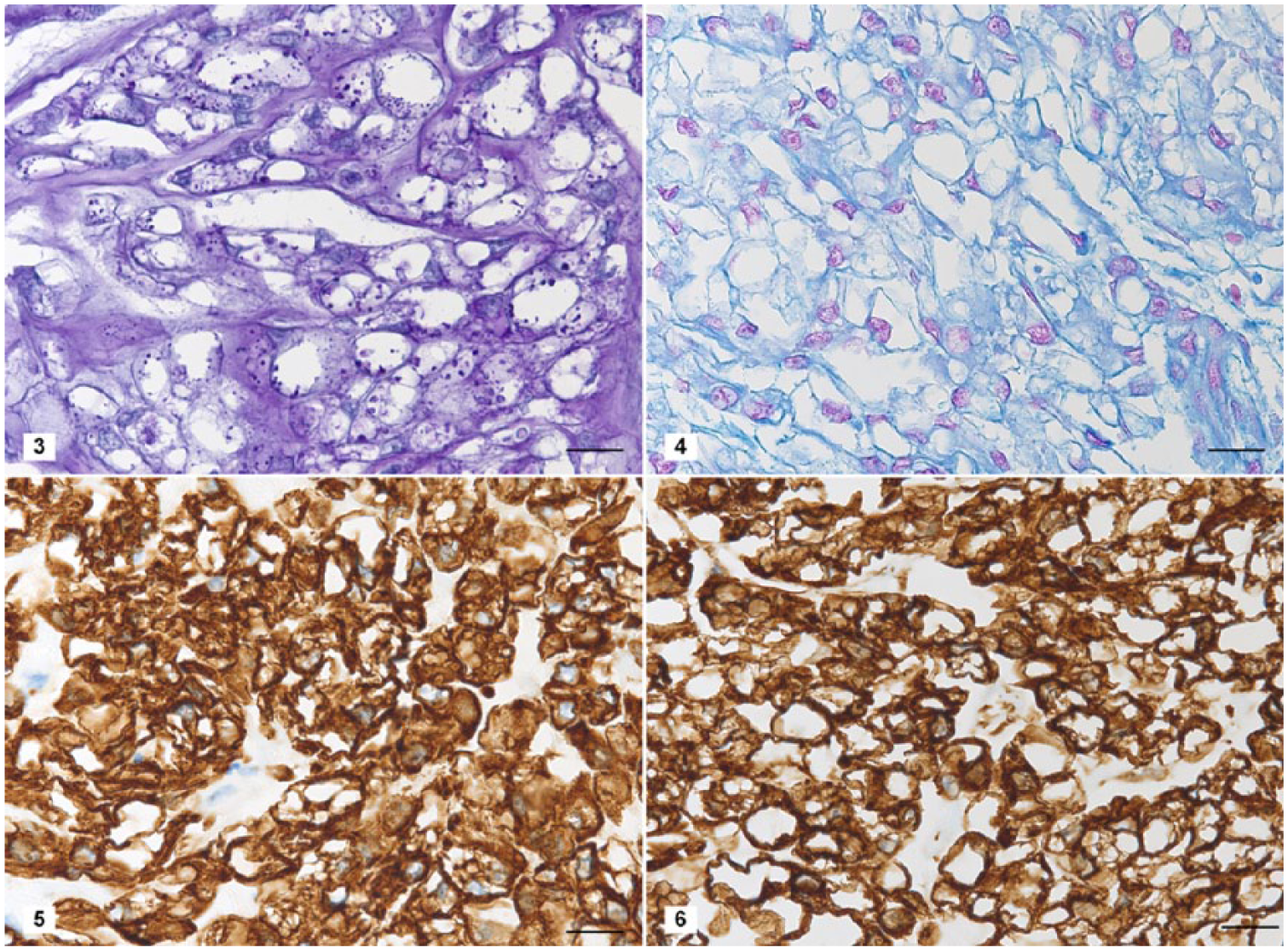
Special stains and immunohistochemical features of the coccygeal mass in a degu.
Tumors reported in degus include 8 epithelial, 13 mesenchymal, and 4 round cell tumors, and 2 chordomas (Supplementary Table 1). The age, sex, anatomic location, and outcome were not reported for one chordoma (Zadravec M, et al. Spontaneous tumours and non-neoplastic proliferative lesions in pet degus). The other chordoma was a 20 × 20 × 15 mm mass located at the base of the tail of a 6-y-old female degu that died from respiratory failure 3 mo after tail amputation. 12 The diagnosis was based on the histologic appearance of the excised mass. In both chordoma cases, the diagnosis was based on hematoxylin-and-eosin–stained sections of the amputated mass. Autopsies were not performed.
Chordomas arise from notochordal remnants and have a low incidence in humans and animals 18 (Supplementary Fig. 3). In animals, chordomas have been reported most commonly in ferrets 21 and less frequently in dogs, 14 rats, 17 cats, 2 mink, 8 zebrafish (Danio rerio), 4 Perdido Key beach mice (Peromyscus polionotus trissyllepsis), 19 and a tiger. 13 In human and animal cases, including our degu, chordomas typically occur in older adults (Supplementary Table 2). A few chordomas have been reported in children, 18 and one congenital chordoma has been described in a lamb. 11
The notochord regresses after completing its function of axial positioning in the developing embryo. It is then replaced by the nucleus pulposus of the intervertebral disks. However, a small portion of notochordal cells can still be found as notochordal remnants in adults. 20 The location of chordomas follows the anatomic distribution of notochordal remnants. Human chordomas most commonly occur at the cranial and caudal extremities of the axial skeleton. 18 The distal tail tip is the common location in ferrets. 5 In rodents, location of the tumor varies among the cranial, cervical, thoracic, lumbar, sacral, and coccygeal segments of the axial skeleton.17,19 There is a higher incidence in the lumbosacral area of Fischer 344 rats, 17 and in the thoracic vertebrae of Perdido Key beach mice. 19 Apart from metastases, the only extra-axial location of chordomas reported has been the intestine of aged zebrafish. 4
Chordomas in humans are slow-growing, locally aggressive tumors, with a high local recurrence rate after surgery, but with a low risk of distant metastases. 18 In ferrets, metastases have only been reported twice in the skin and once in the lungs, 7 and in cats, once in regional lymph nodes. 3 In contrast, there is a high rate of metastases in the lungs of Fischer 344 rats 17 and Perdido Key beach mice (Supplementary Table 2). 19 Tail tip chordomas have a good prognosis; there are no reports of metastasis associated with this location in mammals (Supplementary Table 2). Nevertheless, tail amputation and routine check for metastases (e.g., lung radiographs, abdominal ultrasound, palpation of regional lymph nodes) is recommended. In the degu of our report, the coccygeal chordoma was distant to vital structures, making surgical treatment ideal. At the 5-mo follow up, the degu was in a good body condition with no evidence of local recurrence or distant metastasis.
Histologic morphology and immunohistochemical analysis is the gold standard test for chordoma diagnosis. Fine-needle aspiration was inconclusive in our case given the firm consistency of the mass that did not allow many cells to be aspirated. However, cytology has proven useful as a primary diagnostic step for masses of the axial skeleton. 16
Our differential diagnoses included myxosarcoma and myxoid liposarcoma based on the histologic morphology of the tumor. Depending on the anatomic location, the predominant cell-type, and its histologic appearance, other differentials include chondrosarcoma, mucinous adenocarcinoma, sebaceous carcinoma, and signet ring cell carcinoma.1,14 However, the diagnosis in our case was made relatively straightforward by the distinctive PAS-positive cytoplasmic granules in the physaliferous cells, which has been described in chordomas of other species,8,14,17,19,21 and the dual immunohistochemical expression of epithelial and mesenchymal intermediate filaments. 5 Myxosarcomas and liposarcomas do not express cytokeratin and typically exhibit more malignant features such as nuclear atypia and mitoses. Furthermore, frozen sections stained with oil red O can help differentiate a chordoma from a liposarcoma, given that chordomas are devoid of lipids. 5
Other biological markers used in the diagnosis of human and animal chordomas, including the proteins S100 and neuron-specific enolase (NSE), 5 have been investigated and are frequently found in the immunohistochemical panel for diagnosing chordoma. The most exhaustive study to date on the immunohistochemical properties of ferret chordomas revealed that neoplastic cells express brachyury, 21 a protein and transcription factor that is the main regulator of notochord formation and a useful marker of human chordomas. 20 Brachyury has also been detected by immunohistochemistry in chordomas of Perdido Key beach mice. 19
Three distinct but overlapping histologic subtypes of chordoma are recognized in humans: 1) classic (also known as “not otherwise specified (NOS),” “conventional,” “typical,” or “myxoid”), 2) chondroid (a chordoma with a very focal-to-extensive cartilaginous component), and 3) dedifferentiated (a chordoma with a malignant spindle cell component’). 6 In humans, it is important to distinguish these histologic subtypes, at least the dedifferentiated from the others, because there is prognostic significance: the dedifferentiated chordoma carries the worst prognosis, compared to the well-differentiated subtypes (“classic” and “chondroid”). 19 Although the chondroid chordoma has long been associated with a better prognosis compared to the classic subtype, 9 to date, there is no good evidence that chondroid chordomas behave differently from classic chordomas. 6 Although prognostic human chordoma classification schemes 18 have not yet been adapted to mammalian animal chordomas, it is common to find standard human chordoma classifications (e.g., WHO Classification of Soft Tissue Tumours) 6 used in animals for histologic morphology.
Chordomas in Fischer 344 rats and Key Perdido beach mice are histologically most consistent with the human classic subtype and are highly malignant (Supplementary Table 2).17,19 Most chordomas in ferrets are reminiscent of the human chondroid subtype and are commonly benign (Supplementary Table 2). 5 The dedifferentiated subtype has never been characterized in non-human mammals, to our knowledge (Supplementary Table 2). Morphologically, the chordoma in our degu can be classified as a classic chordoma, as in Fischer 344 rats and Key Perdido beach mice, but no evidence of metastasis was seen. Additional data are needed to predict clinical behavior of chordomas in degus.
Supplemental Material
DS1_JVDI_10.1177_1040638718814584 – Supplemental material for Coccygeal chordoma in a degu: case report and review of the literature
Supplemental material, DS1_JVDI_10.1177_1040638718814584 for Coccygeal chordoma in a degu: case report and review of the literature by Lucile Sautier, David Guillier, Margaux Coste, Jean-Luc Servely, Charly Pignon, Eve Laloy and Thomas M. Donnelly in Journal of Veterinary Diagnostic Investigation
Supplemental Material
DS2_JVDI_10.1177_1040638718814584 – Supplemental material for Coccygeal chordoma in a degu: case report and review of the literature
Supplemental material, DS2_JVDI_10.1177_1040638718814584 for Coccygeal chordoma in a degu: case report and review of the literature by Lucile Sautier, David Guillier, Margaux Coste, Jean-Luc Servely, Charly Pignon, Eve Laloy and Thomas M. Donnelly in Journal of Veterinary Diagnostic Investigation
Supplemental Material
DS3_JVDI_10.1177_1040638718814584 – Supplemental material for Coccygeal chordoma in a degu: case report and review of the literature
Supplemental material, DS3_JVDI_10.1177_1040638718814584 for Coccygeal chordoma in a degu: case report and review of the literature by Lucile Sautier, David Guillier, Margaux Coste, Jean-Luc Servely, Charly Pignon, Eve Laloy and Thomas M. Donnelly in Journal of Veterinary Diagnostic Investigation
Supplemental Material
TS1-2_JVDI_10.1177_1040638718814584 – Supplemental material for Coccygeal chordoma in a degu: case report and review of the literature
Supplemental material, TS1-2_JVDI_10.1177_1040638718814584 for Coccygeal chordoma in a degu: case report and review of the literature by Lucile Sautier, David Guillier, Margaux Coste, Jean-Luc Servely, Charly Pignon, Eve Laloy and Thomas M. Donnelly in Journal of Veterinary Diagnostic Investigation
Footnotes
Acknowledgements
We thank Agnès Champeix, Dorine Chevalier, Emmanuelle Duhec, Sonita Ing, and Narcisse Towanou for technical assistance, James Walberg for reviewing the manuscript, and Yessica Servely for critically proofreading the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.