
Abstract
Bovine viral diarrhea virus (BVDV) infects cattle populations worldwide, causing significant economic losses though its impact on animal health. Previous studies have reported the prevalence of BVDV species and subgenotypes in cattle from the United States and Canada. We investigated the genetic diversity of BVDV strains detected in bovine serum samples from 6 different Mexican regions. Sixty-two BVDV isolates from Mexico were genetically typed based on comparison of sequences from the 5′ untranslated region (5′-UTR) of the viral genome. Phylogenetic reconstruction indicated that 60 of the samples belonged to the BVDV-1 genotype and 2 to the BVDV-2 genotype. Comparison of partial 5′-UTR sequences clustered 49 samples within BVDV-1c, 8 samples within BVDV-1a, 3 samples within BVDV-1b, and 2 samples clustered with the BVDV-2a subgenotypes. Our study, combined with information previously published on BVDV field strain diversity in the United States and Canada, benefits the development of effective detection assays, vaccines, and control programs for North America.
Bovine viral diarrhea virus (BVDV) is associated with a wide range of clinical manifestations in cattle, including severe respiratory disease, gastrointestinal disorders, hemorrhagic syndrome, and reproductive problems such as reabsorption, abortion, infertility, stillbirth, congenital malformations, mummification, and the birth of weak calves or persistently infected animals, as a result of a transplacental infection in the first trimester of gestation. 23 The reproductive disorders resulting from BVDV infection lead to significant economic losses to the cattle industry. 9
BVDV is a member of the Pestivirus genus (family Flaviviridae), which includes 4 recognized species: Bovine viral diarrhea virus 1 (BVDV-1), Bovine viral diarrhea virus 2 (BVDV-2), Border disease virus, and Classical swine fever virus. 23 In addition, 4 putative species of Pestivirus have been proposed: bovine viral diarrhea virus 3 (BVDV-3) or “HoBi-like” virus, Bungowannah virus, pronghorn virus, and giraffe virus, but these remain officially unrecognized. 12 One of the putative species, HoBi-like virus, is a bovine pathogen that produces clinical disease similar to that seen with infections of BVDV-1 or -2. To date, HoBi-like viruses have been detected in South America, Europe, and Asia, but not North America. The viruses grouped as BVDV are highly heterogeneous and encompass viruses belonging to 2 different species and 2 different biotypes, cytopathic (CP) and noncytopathic (NCP), based on their effect in cultured cells. 17 The 2 BVDV species are subdivided into subgenotypes. At least 11 subgenotypes of BVDV-1 25 and 3 subgenotypes within BVDV-2 are reported. 6
The first reported isolation of BVDV in North America was in the United States in 1957. 11 Subsequently, 2 subgenotypes of BVDV-1 (BVDV-1a and -1b) and 1 subgenotype of BVDV-2 (BVDV-2a) have been reported in bovine samples from the United States and Canada.15,19 In Mexico, the first report of BVDV was in 1975 based on a study to detect antibodies in cows with clinical history of infertility and abortions, showing a seropositivity of 75%. 3 Other studies have also revealed the presence of antibodies against BVDV, showing a seroprevalence of 14–29%.8,21 However, none of these studies provide information regarding the genetic diversity of BVDV in Mexico. Because there is no information available about the BVDV genotypes and subgenotypes currently circulating in Mexican cattle, the aim of our study was to detect and characterize BVDV-1 and -2 and survey for HoBi-like viruses in bovine serum samples in cattle from 6 different regions of Mexico, and survey the diversity of BVDV present in Mexican cattle.
A total of 402 serum samples from 6 farms located in 6 different regions of Mexico including Hidalgo, Morelos, Querétaro, Veracruz, Tamaulipas, and Chihuahua were used for this study. The herd sizes varied from 56 to 134 animals and included both dairy and beef breeds.
Total RNA was extracted from a 140-µL aliquot of serum from each animal, using a robotic workstation (QIAcube, Qiagen, Hilden, Germany) for automated RNA purification by a spin column system (QIAamp viral RNA mini kit, Qiagen) according to the manufacturer’s instructions. RNA was eluted in 50 µL of RNase-free water and stored at −70°C.
The RNA prepared from serum samples and positive controls, consisting of RNA prepared from the reference strains BVDV-1a Singer, BVDV-2a 296c, HoBi-like virus Italy-1/10-1, were submitted to reverse transcription–polymerase chain reaction (RT-PCR). The set of primers 324-326, HCV90-368, 5UTR/START, and HoBi N2/R5 were used as described previously in order to amplify a fragment of the 5′ untranslated region (5′-UTR).2,13,19,24 The PCR products were detected by electrophoresis in 1% agarose gel and then purified using a commercial kit (QIAquick gel extraction kit (Qiagen) following the manufacturer’s recommendations. The nucleotide sequences of the PCR products were sequenced, (ABI Prism 3130xl genetic analyzer, Applied Biosystems/Hitachi, Forest City, CA), and all sequencing reactions were done in duplicate and in both directions.
Phylogenetic analysis was based on the comparison of a 270 nucleotide region of the 5′-UTR using the maximum likelihood method with a Kimura 2-parameter 10 substitution model using commercial software (BioEdit v7.1.9, Tom Hall Ibis Biosciences, Carlsbad, CA; MEGA 6 v.6.0, Center for Evolutionary Medicine and Informatics, Tempe, AZ).
Sixty-two of the 402 samples were positive using the 324-326, HCV90-368, or 5UTR/START primers (Table 1). None of the serum samples tested was detected as positive by RT-PCR using the primer set N2/R5, specific for HoBi-like virus. Thirty-two of the 62 sequences showed a 100% identity, represented with sample C6487 in the tree (Fig. 1). These 32 samples belong to herds from Veracruz, Tamaulipas, Queretaro, Morelos, and Chihuahua. Based on the nomenclature of BVDV groups suggested previously, 25 the positive samples could be divided into 4 different subgenotypes; 49 clustered within BVDV-1c, 8 were typed as BVDV-1a, 3 as BVDV-1b, and 2 were characterized as BVDV-2a. No HoBi-like virus was found.
Genetic typing of the 62 Mexican bovine viral diarrhea virus samples used in the current study.

Samples represented by C6487 are shown in boldface.
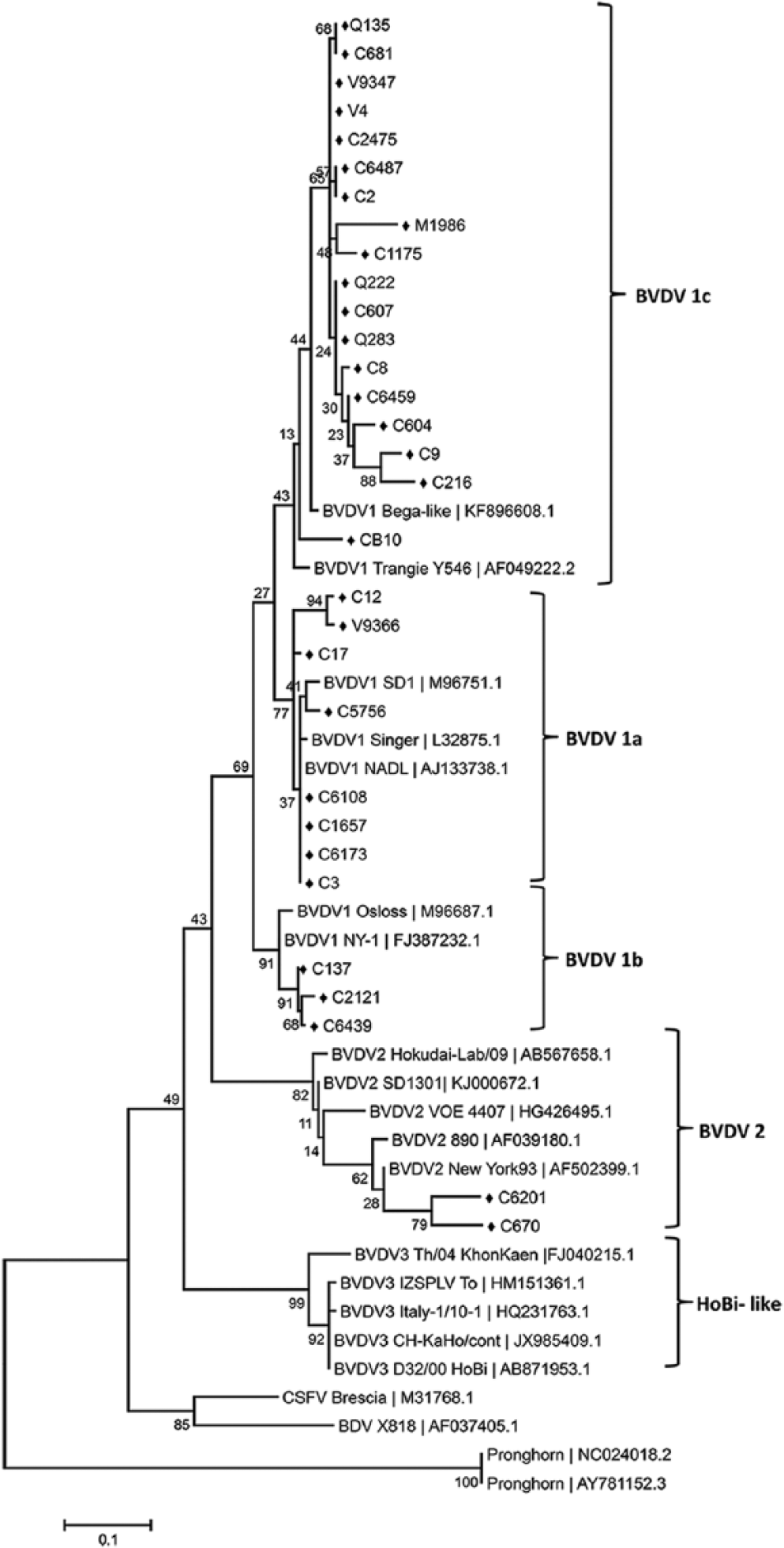
Phylogenetic tree based on the 5′ untranslated region. Phylogeny inference was conducted in MEGA 6 (Center for Evolutionary Medicine and Informatics) using maximum likelihood method. Distances were computed using the Kimura 2-parameter model. Reference sequences are identified by GenBank accession. The Mexican nucleotide sequences are labeled with the symbol “♦”.
Our investigation describes the genetic diversity of BVDV from cattle in Mexico, and shows that at least 3 BVDV-1 (BVDV-1a, -1b, and -1c) and 1 BVDV-2 (BVDV-2a) subgenotypes were present in the surveyed population. Subgenotypes BVDV-1a and -1b and BVDV-2a have been detected in cattle in the United States or Canada.7,20 The BVDV-1c subgenotype is the most prevalent subgenotype circulating in Australia, but to date has not been detected in cattle from the United States or Canada. 20 No HoBi-like viruses were identified in serum samples during the investigations of our study. This correlates with a previous survey of fetal bovine sera originating in North America that failed to detect HoBi-like viruses. 2 However, it is not definitive proof that HoBi-like viruses are not circulating in Mexico. Further surveillance studies sampling larger numbers of animals originating across Mexico are necessary to demonstrate the absence of HoBi-like viruses.
Our survey revealed that, among the 62 viral samples analyzed, BVDV-1c was the virus detected most commonly, followed by BVDV-1a, -1b, and -2a. Thus, the mix of BVDV circulating in Mexico is unique in North America. In addition to being the predominant BVDV subgenotype in Australia, 13 BVDV-1c has also been detected in Germany 22 and Spain. 1 The presence of this subgenotype might be associated with cattle trade between Mexico and Australia because Mexico is the only country in the Central American region to import live Australian animals. In contrast, the BVDV-1b subgenotype appears to be the most common BVDV subgenotype found in the neighboring countries of the United States 19 and Canada. 15 In the United States, BVDV-1b strains have been shown to break through the protection provided by vaccines containing BVDV-1a and -2a viruses. 5 Previous studies have shown antigenic variation between BVDV-1a and -1c that are statistically similar to that seen between BVDV-1a and -1b strains. 20 Antigenic differences between BVDV-1b strains and BVDV-1a and -2a strains have led to the suggestion that vaccines would be more effective if BVDV-1b strains were included. 4
Only 2 subgenotype 2a–positive samples were detected, showing a low frequency. BVDV-2b, -2c, and -2d strains, which have been reported in South America, 6 were not detected. The low frequency of BVDV-2 strains is similar to findings in the United States, Australia, and the United Kingdom. 20 The BVDV-2a subgenotype appears to be more common than the other subgenotypes worldwide. 14
Our results provide information about the genetic diversity of BVDV, suggesting that at least 4 subgenotypes are currently circulating in Mexican cattle. Antigenic differences 16 among BVDV subgenotype strains are associated with vaccine failure. 4 Several commercial vaccines containing BVDV-1a and -2a strains are currently being used in Mexico. It has been suggested that vaccine efficacy could be improved by including in vaccines the BVDV strains prevalent in the region in which the vaccine is going to be used. 18 Monitoring the diversity among BVDV strains is recommended, especially to drive the design of new vaccines able to promote a protective response against the virus, to contribute to the development of effective eradication and control strategies, and when designing accurate detection assays to determine likely infection sources. To date, there is not enough data on BVDV epidemiology and distribution in Mexico to allow the informed design of vaccines, testing, and control programs.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflict of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this project was provided by Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT, IT201516).