
Abstract
Cysticercus fasciolaris, the larval form of Taenia taeniaeformis, is commonly encountered in rodents. In our study, 287 wild rats (Rattus norvegicus) in South Korea were examined in 2010 and 2011. Of 287 rats, 97 (33.8%) were infected with C. fasciolaris. A strong positive correlation was found between the host body weight and prevalence in both sexes, regardless of the year of collection. The liver was the most common habitat of the parasite, and the lung was the most frequent ectopic region, followed by mesentery, pleura, abdominal wall, and kidney. The lesions of the affected organs were generally characterized by well-developed cysts, each containing a larva. However, the cysts within kidney and abdominal wall were poorly organized, filled with abscess, and lacked larvae. Collagen types I and III, but not type IV, played significant roles in constructing the cysts at differential stages, addressed by immunohistochemistry. During cyst wall development, both collagen types contributed equally to cyst formation at the early stage, whereas collagen type I was the major component at the late stage (p < 0.05). In early-stage cysts, distribution of collagens was interestingly differential depending on the development stage, as collagen type I was localized in the outer layer and type III was located in the inner layer. Our results suggest that an appropriate remodeling process of collagen fibers is necessary for C. fasciolaris to build the well-conditioned cysts in the target organs for survival.
Introduction
Cysticercus fasciolaris is the larval form of the cestode, Taenia taeniaeformis, also called a tapeworm, in cats. The metacestode inhabits the liver of the rodent, an intermediate host. 27 C. fasciolaris, whose definitive hosts are carnivores in the families Felidae and Canidae, commonly encounters wild rats (Rattus norvegicus) through contamination of feed and bedding by the embryonated eggs. 27 Once ingested by the wild rats, embryonated eggs hatch in the small intestine and migrate via the portal circulation into the liver, where they develop into cystic larvae. 27 Hatched embryos can also migrate into other ectopic organs, such as the lung and mesentery, and grow into a metacestode, albeit with low probability.22,26
Cysticercus fasciolaris infection in wild rats has been reported worldwide, and its prevalence varies depending on where and when the rats are captured. The reported prevalence is 100% in the Philippines, 5 33.3% in India, 26 29.9% in Serbia, 20 and 11% in rural England. 3 In the Republic of Korea, the prevalence has been reported to be 20% in Seoul, 23 15.2% in Chulwon, Kumhwa, Chungpyong, Pochun, and Paju, 24 and 65.1% in Chunchon. 25 However, the prevalence of C. fasciolaris infection has not been studied in recent decades in the Republic of Korea or other countries, and the presence of lesions in organs other than the liver has not been fully addressed. Larvae that have migrated to the liver via the portal circulation induce the liver to recruit inflammatory cells, including lymphocytes, neutrophils and eosinophils, which form a thick layer surrounding the larva and localize the parasite infection. 28 Although this is the defense mechanism of the host, the larvae take advantage of the host responses to build cysts that house the larvae, protecting them from the host inflammatory response and providing them a good environment to mature. 28 However, little information is available concerning the cellular and molecular mechanisms of cyst formation in the parasite-infected liver.
The cyst wall built in response to the parasites is generally composed of collagen-rich connective tissue and interspersed small neovessels with inflammatory cell infiltration, morphology similar to that of the granulation tissue. Thus, cyst formation by C. fasciolaris could be achieved by mechanisms similar to those in the repair process initiated following parasite-induced liver injury. The formation of granulation tissue in the liver consists mainly of serial stages of fibrosis initiated by the imbalance between the synthesis and degradation of extracellular matrix components. 11 Hepatic stellate cells (HSCs) have been postulated to play an important role in producing collagen in the injured liver. 12 Following injury of liver parenchyma, the proliferative and fibrogenic myofibroblasts originating from HSCs produce excessive collagen fibers via exocytosis.2,18,29 Various collagen types such as I and III contribute to liver fibrosis, and their localization is dependent on the stage of the fibrogenic process. In addition, collagen type IV, the major collagen type of basal lamina, could also be considered in cystic wall formation, as it has been reported to be expressed in HSCs.12,13
Thus, we hypothesized that fibrogenic mechanisms might be significantly involved in cyst formation by C. fasciolaris, which is critical for survival and development of cysticercus larvae in the liver. In our study, in addition to addressing the prevalence of C. fasciolaris infection in wild rats in Korea, we histopathologically assessed the changes, localization, and relative accumulation of collagen types during cyst formation.
Materials and methods
Animals, gross pathology, and parasite identification
A total of 287 wild rats were captured using wire traps from the areas around 10 selected cattle farms in Gyeonggie, Gangwon, and Chungbuk provinces, Republic of Korea, from 2010 to 2011. The rats were weighed and classified into 3 age groups on the basis of their body weight: immature (female: <100 g; male: <100 g), pubertal (female: 101–175 g; male: 101–200 g), and postpubertal (female: >175 g; male: >200 g; Table 1). 19 The wild rats were euthanized and autopsied, and location, number, and size of cysts together with the length of larvae were recorded. Following gross examination, all major parenchymal organs including the organs that contain cysts were fixed in 10% neutral buffered formalin, processed routinely for paraffin embedding, sectioned at 4 μm thickness, and stained with hematoxylin and eosin (HE). After removal from the cysts, the parasites were stained with Semichon acetocarmine to confirm the presence of C. fasciolaris. All animals used for this study were treated in accordance with the criteria outlined in the “Guide for the Care and Use of Laboratory Animals” of Kangwon National University, and this study was approved by the Institutional Animal Care and Use Committee of Kangwon National University.
Prevalence of Cysticercus fasciolaris in wild rats (Rattus norvegicus).*
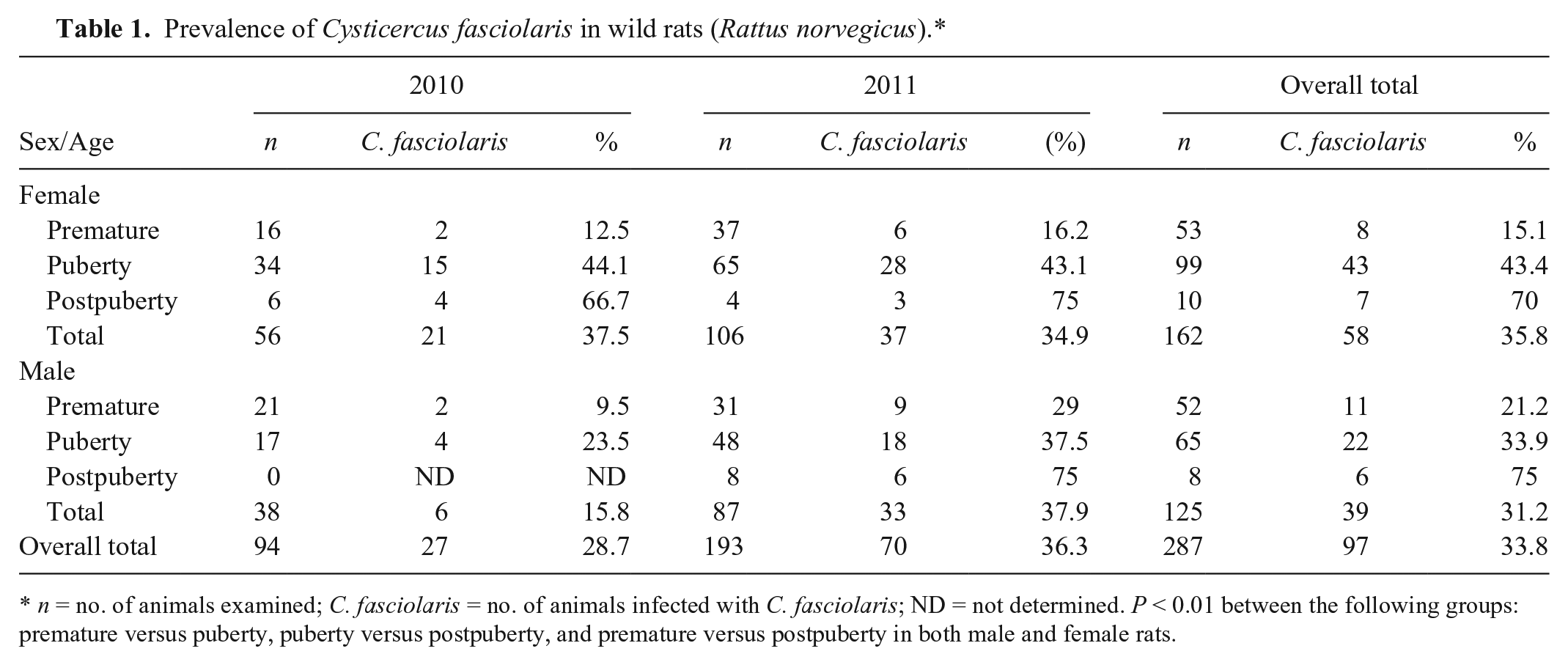
n = no. of animals examined; C. fasciolaris = no. of animals infected with C. fasciolaris; ND = not determined. P < 0.01 between the following groups: premature versus puberty, puberty versus postpuberty, and premature versus postpuberty in both male and female rats.
Classification of the stages of C. fasciolaris infection
We classified either early- or late-stage C. fasciolaris infection on the basis of the histomorphologic characteristics of larva or cyst. 28 The early stage of C. fasciolaris infection was characterized by a relative thick cyst wall with premature larva, whereas the late stage of infection was characterized by a thin cyst wall and mature parasite with well-matured organs.
Immunohistochemistry
The proportions of collagen types I and III in cyst walls at different stages of parasite infection were evaluated by immunohistochemistry (IHC). On the basis of histopathologic examinations, 5 and 13 fresh specimens were selected randomly for early and late infection stage, respectively, and sectioned at 6 μm thickness for IHC. All sections were deparaffinized in xylene, then rehydrated and subjected to antigen retrieval using citrate buffer. a After incubation with normal serum b for 1 h at room temperature to block nonspecific binding, tissue sections were then incubated overnight at 4°C with primary antibodies against collagen types I, III, and IV fibers [rabbit polyclonal anti-human collagen type I (1:500), III (1:500), and IV (1:250)]. c Tissue sections were then washed in phosphate buffered saline and incubated for 1 h at room temperature with biotin-conjugated anti-rabbit immunoglobulin G secondary antibodies, followed by incubation with avidin–biotin complex reagent for 30 min at room temperature. DAB (3,3′-diaminobenzidine) solution was used as a chromogenic substrate. Sections were counterstained with modified Mayer hematoxylin, then dehydrated, cleared in xylene, and mounted.
Morphometry of collagens in cysts
For morphometric analysis of collagen type I or III, images of the cyst walls were obtained using a light microscope at 400× magnification, and immunoreactive areas were measured using a digital imaging analysis system. d The proportions of areas immunostained for collagen type I or III were determined as the percentage of positive area in the entire cyst wall.
Statistical analyses
A chi-square test was used to determine if the prevalence of C. fasciolaris infection in wild rats was related to sex, year of collection, or host body weight. A Cochran–Mantel–Haenszel test was then used to evaluate the association between prevalence and host body weight. Morphometric data for collagen types I and III were analyzed using Student t-test to assess the significance of differences between early and late stages of infection, and a paired t-test was used to assess the significance of differences in the proportions of collagen types I and III at the same stage of infection. The SAS statistical software package e was used for statistical analyses. Significant differences between groups were determined at p < 0.05 or p < 0.01.
Results
Prevalence and pathological characteristics of C. fasciolaris infection
A total of 287 rats were captured from 2010 to 2011 (94 in 2010 and 193 in 2011; 162 females, 125 males; Table 1). On gross and microscopic examination, 97 of 287 rats (33.8%) were found to be infected with C. fasciolaris. There were no significant differences in prevalence between year captured (28.7% in 2010; 36.3% in 2011) or sex (35.8% in female; 31.2% in male; Table 1). We did find, however, that the prevalence of C. fasciolaris infection was significantly different among age groups, increasing with age and reaching a peak in the post-pubertal group in both sexes, regardless of the year of collection (p < 0.01; Table 1).
Cyst formation was found in the liver of all infected rats (Table 2). Table 2 shows that cysts were also present in other organs, including the lung (6.2%), mesentery (2.1%), pleura (2.1%), abdominal wall (1.0%), and kidney (1.0%). The lengths of larvae found in the cysts varied from 1 mm to 20 cm (Fig. 1A). The larvae were loosely coiled, characteristically lacked a neck, and had bell-shaped posterior proglottids (Fig. 1A). The scolex was large, with 2 rows of rostellar hooks (Fig. 1B). The morphology of the parasites corresponded to the larval form of T. taeniaeformis. 21 In a rat heavily infected with larvae, ~80% of the liver was replaced by cysts (Fig. 1A), and cysts in the abdominal wall (27 cysts) and kidney (1 cyst) were filled with purulent exudate and lacked larvae. Histologically, hepatic cysts from the liver parenchyma were well organized with granulation tissue (Fig. 1C, 1D). Although indistinct at the early stage (Fig. 1E), the well-developed cyst wall was divided into 3 layers: 1) an inner layer of cyst wall composed of a cytokeratin-immunoreactive squamous cell lining (Supplemental Fig. S1, available online at http://vdi.sagepub.com/content/by/supplemental-data), 2) attenuated hepatocytes, and 3) relatively thick connective tissue with infiltration of inflammatory cells, mainly lymphocytes and plasma cells and a few eosinophils (Fig. 1F). The cyst walls differed somewhat in thickness and density of connective tissue depending on the developmental stage of the larva, with the cysts of late-stage larvae having thinner and denser connective tissue than those of the early stage (Fig. 1C–1F). The cysts noted in other organs exhibited similar histological features as those in the liver. In parasite-containing cysts in the lung, hyperplastic bronchioles with mucous metaplasia surrounded the wall. The histological morphology of cysts in the abdominal cavity and pleura was similar to those in the liver and lung. Interestingly, cysts formed in the kidney and abdominal wall lacked a parasite and were instead filled with degenerate neutrophils (Fig. 1G). In the kidney, the cyst was well demarcated and separated from the renal parenchyma by a cyst wall composed of vesicular epithelial cells and thick connective tissue, with infiltration of predominantly lymphocytes and plasma cells with a few eosinophils (Fig. 1G). The cyst wall was poorly organized, with no inner squamous cell lining and no dense connective tissue layer, similar to that of cysts at the early stage of larval development (Fig. 1G).
Organs having the cysts in the wild rats infected with Cysticercus fasciolaris in Korea.*

Data are presented as number of cases/total number of animals with C. fasciolaris infection. Numbers in parentheses are percentages.
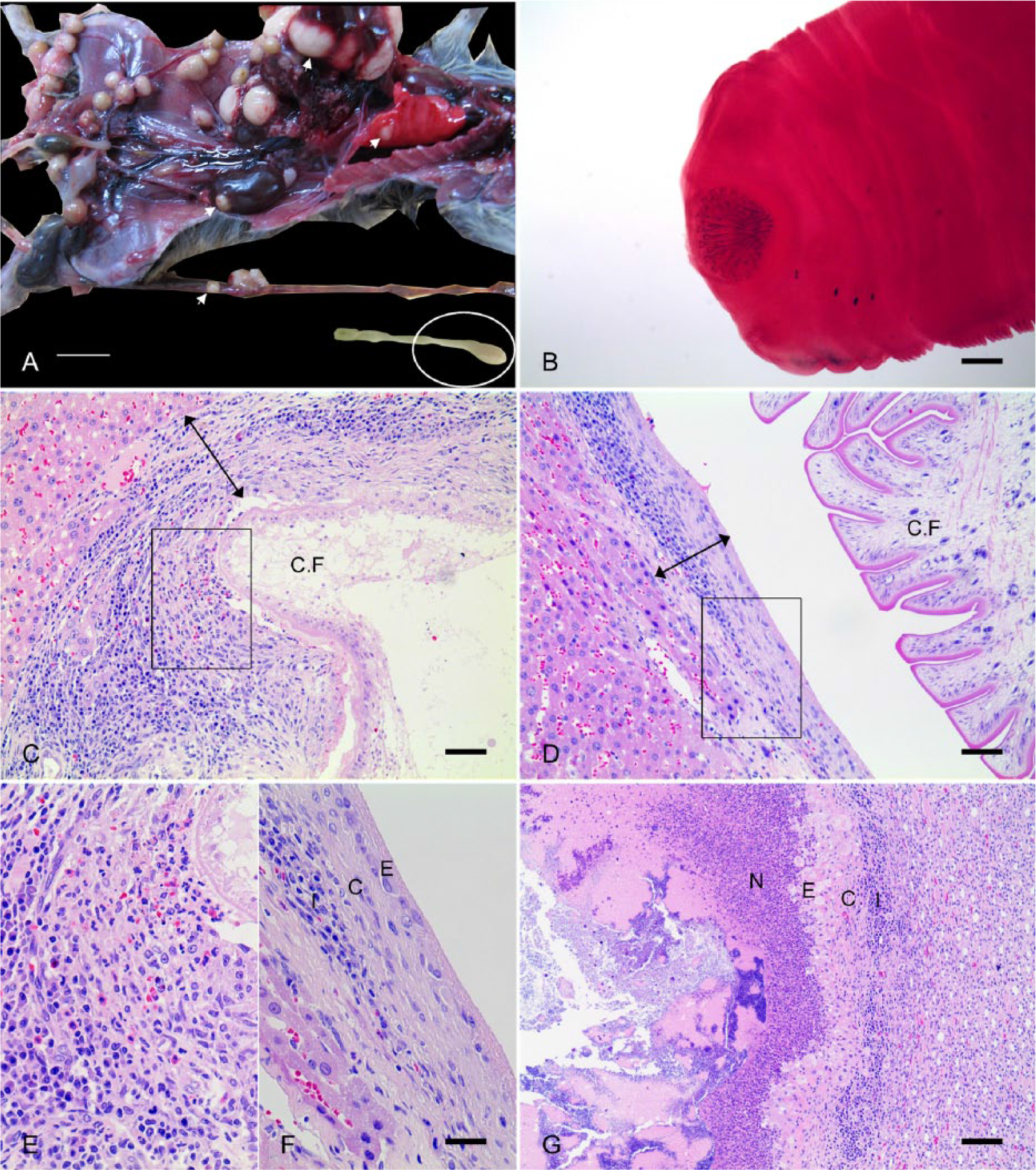
Pathology of Cysticercus fasciolaris infection.
Classification of infection stage and remodeling process of collagen types in the cyst wall
Parasite infections were classified as early or late stage, and the stage-dependent localization of collagen types I and III was determined. In the early infection stage, larvae were poorly developed, and the cyst wall (granulation tissue surrounding the larva) was thicker and looser than that of the late infection stage (Fig. 1C–1F). The collagen type I fibers were mainly localized in the outer layer of the cyst wall close to the liver parenchyma (Fig. 2A), whereas collagen type III fibers were more in the inner layer near the larva (Fig. 2C). The proportions of areas immunoreactive for collagen type I and III relative to the entire cyst wall were 56 ± 26.1% and 58 ± 8.4%, respectively, values that were not significantly different (Table 3; Fig. 2A, 2C).
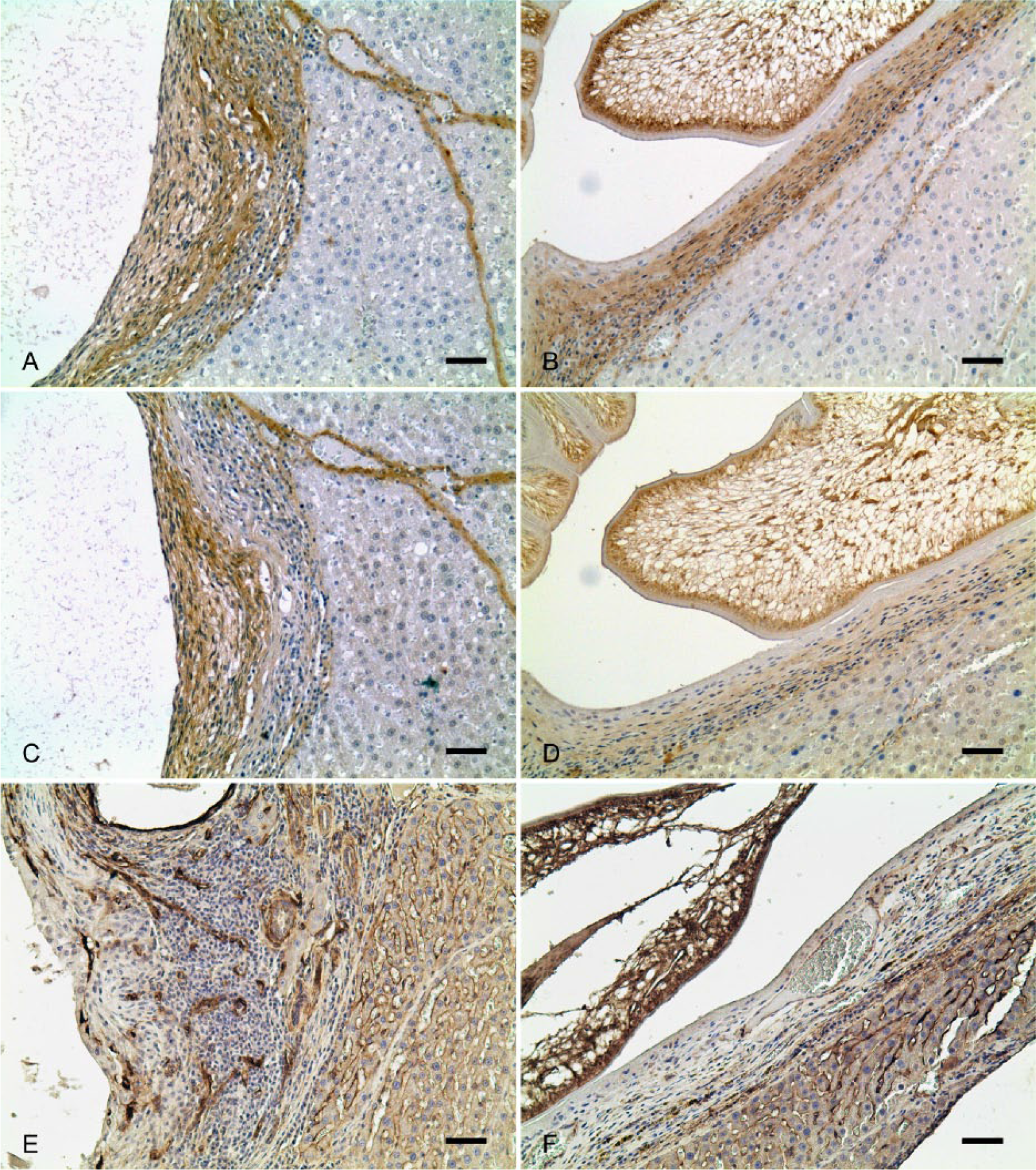
Immunohistochemistry (IHC) for collagen types I, III, and IV in the early and late stage of the parasitic cyst.
Morphometric analysis of collagen types I, III, and IV in cyst walls at early and late stages*

Data are presented as mean ± standard deviation. P < 0.05 between the type I and III groups.
In contrast, the late infection stage larval structures were well developed. These cysts were larger in diameter and thinner than those of the early stage (Fig. 1D, 1F). Collagen type I fibers were localized throughout the cyst wall, and constituted ~70% (69.6 ± 23.9%) of the total collagen (Table 3; Fig. 2B). Interestingly, collagen type III fibers covered only 40.8 ± 29.5% of the total area, and its expression was very weak (Table 3; Fig. 2D). Thus, the proportion of collagen type I accumulated in the late infection stage was significantly greater than that of collagen type III (Table 3; p < 0.05). On the other hand, collagen type IV fiber was not expressed in the cystic walls at either early or late stage of infection (Table 3; Fig. 2E, 2F). In normal livers, round cells positive for smooth muscle actin (SMA) were randomly distributed in the sinusoidal spaces, whereas SMA-positive cells were oval and flattened, and frequently distributed outside of the connective tissue layer of the cysts where there were abundant collagens (Supplemental Fig. S2, available online at http://vdi.sagepub.com/content/by/supplemental-data).
Discussion
In our study, the prevalence of C. fasciolaris infection in wild rats collected from 2010 to 2011 in the Republic of Korea averaged 33.8%. The prevalence trended higher in 2011 than in 2010 (a difference that did not reach statistical significance), but an estimate of ~30% in both years indicates that infection by this parasite in the Republic of Korea is endemic. This rate is higher than that in previous studies in 1964 23 (20%) and 1968 24 (15.2%), but lower than that in 1995 25 (65.1%). Prevalence and intensity of infection were positively correlated with body weight in both years of collection, but were independent of sex. This age-associated prevalence is in agreement with a prior report. 4 A previous report on albino rats suggested that 25- to 60-day-old rats are more susceptible to this parasite than younger or older animals. 14 However, the survival of helminth ova is known to be several months in the natural environment, and the parasites cannot escape from wild rodents—their intermediate hosts—throughout their lifetime.9,10 Thus, we suggest that the observed prevalence is more closely associated with the accumulation of infection with age rather than host age–dependent differences in susceptibility to the parasite.
Although histological features of cysts in infected livers have been well described in previous reports, migration into other organs, including lung and abdominal wall, and the histological features of these cysts have been rarely documented.7,22,26,28 The histological features of cysts with a larva in other organs were similar to those of cysts in the liver, suggesting that the larva can live and develop in the lung, mesentery, and pleura as well as the liver. The development of a larva in an infected lung induced glandular epithelial hyperplasia with mucous metaplasia. Cysts in the kidney and abdominal wall in 1 infected rat in our study were unusual; their walls were incomplete, with no inner squamous cell lining and a looser connective tissue layer, and they contained necrotic neutrophils instead of a larva. The reasons for the degeneration of ectopically migrated larvae and the mechanisms by which it occurs are unknown, but a prior report suggested that entry of the gram-positive bacillus, Corynebacterium kutscheri (previously C. murium), into the lesion with a migrated larva may lead to death of the larva. 17 This unusual rat was also heavily infected, with 75–80% of the liver parenchyma replaced by cysts. Therefore, larvae, which could not be located in the liver, may have exited the liver via the central vein and migrated into other organs via the systemic circulation, and the subsequent degeneration of larvae may have been a consequence of failure to construct appropriate living conditions in other organs.
Histologic examinations revealed that the composition of cyst walls surrounding larvae were similar to that of granulation tissue characteristic of tissues recovering after injury. The composition of cyst walls was further examined by IHC to assess the localization and density of collagen type I, III, and IV fibers accumulated within cyst walls. Our results indicated that collagen types I and III play significant roles in the formation of a cyst wall, whereas the collagen of basal lamina, type IV, was not significant in either early or late infection stage. According to the previous study, HSC is a unique cell, expressing SMA among the perisinusoidal cells of liver. 8 The inference is that migration of C. fasciolaris into the liver stimulates the activation of HSCs, inducing them to surround the larva and differentiate into myofibroblasts, which then produce collagen type I and III fibers. Collagen type III accumulated at the early stage of infection and were replaced by collagen type I fibers at the late stage. This sequence of events resembles the process by which granulation tissue forms following Mesocestoides vogae and Echinococcus multilocularis infection in the liver.6,15,16 In addition, collagen type I fibers are known to be a stronger form of the structural protein than collagen type III. 1 Therefore, as the larva develops, the cyst wall, composed of collagen networks, gradually becomes tougher. The possible role of HSCs in the cyst wall formation by cysticercus larvae in the liver was strongly suggested, as the SMA-immunoreactive cells surrounded the cyst walls (Fig. S2).
In general, the infection of C. fasciolaris in the liver induces an inflammatory response, which represents the host’s attempt to eliminate the larva. 28 To protect the liver parenchyma from the larva, the infected livers then form a capsule surrounding the larva, composed of infiltrated inflammatory cells and collagen fibers secreted by HSCs and/or myofibroblasts.15,28 As the larva develops, it becomes fully encapsulated, and the number of inflammatory cells decreases. Thereafter, the larvae are judged by the host to no longer be a severe irritant to the liver and the immune response abates. 15 Therefore, cyst formation by C. fasciolaris infection seems to represent a double-edged sword: it protects the infected liver against the larva, but it also creates a habitat that allows the larva to avoid the host immune response.
In summary, from 2010 to 2011, the prevalence of C. fasciolaris infection among wild rats was 33.8% in some provinces of Republic of Korea. The prevalence was positively correlated with body weight, presumably reflecting the accumulation of infection with age. The liver was the most common habitat of the parasite, and the lung was the most frequent ectopic site. C. fasciolaris was also found to migrate and form cysts in the mesentery, abdominal muscle, and kidney, but failed to survive in the abdominal muscle and kidney. The formation of well-developed cysts in liver following C. fasciolaris infection may involve a significant collagen type I, possibly produced by HSCs and/or myofibroblasts, resulting in mutual protection of the liver parenchyma against the larva and the larva against the host immune response.
Footnotes
Acknowledgements
We thank Mr. Byung-Tae Yoon for his excellent assistance in capturing wild rats in South Korea. The authors also thank Drs. Sul-Hwa Jeong and Hyung-Gi Kim for their great help in this study.
Authors’ contributions
BW Lee contributed to conception and design of the study; contributed to acquisition, analysis, and interpretation of data; drafted the manuscript; and critically revised the manuscript. BS Jeon contributed to conception and design of the study, and contributed to acquisition, analysis, and interpretation of data. HS Kim contributed to design of the study, and contributed to acquisition and analysis of data. HC Kim contributed to conception of the study; contributed to acquisition, analysis, and interpretation of data; and drafted the manuscript. BI Yoon contributed to conception and design of the study; contributed to analysis and interpretation of data; drafted the manuscript; and critically revised the manuscript. All authors gave final approval and agreed to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
a.
Scytek Laboratories Inc., West Logan, UT.
b.
Vector Laboratories Inc., Burlingame, CA.
c.
Abcam, Cambridge, MA.
d.
IMT iSolution Lite version 8.0, IMT Inc., Vancouver, British Columbia, Canada.
e.
SAS Institute Inc., Cary, NC.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the author ship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This study was supported by a grant (project code Z-AD21-2010-10-01) from the National Veterinary Research and Quarantine Service, Ministry of Agriculture, in 2010.