
Abstract
Fetal tissues and placenta from a third trimester Mediterranean miniature donkey (Equus asinus) abortion were submitted to the Washington State University, Washington Animal Disease Diagnostic Laboratory for abortion diagnosis. Microscopic examination of formalin-fixed tissues revealed multifocal necrotizing placentitis. Several cells within the necrotic foci contained large, eosinophilic, intranuclear inclusions. Virus isolation from fresh, frozen placenta identified a cytopathic, syncytia-forming virus. Polymerase chain reaction (PCR) from the cultured virus using degenerate universal herpesvirus primers amplified a 699–base pair portion of the DNA polymerase gene. The PCR amplicon had 96.7% nucleotide identity with the DNA polymerase gene of Equid herpesvirus 7 (EHV-7; asinine herpesvirus 2), a gammaherpesvirus. An identical sequence was obtained when the same degenerate herpesvirus primers were used for PCR on the formalin-fixed placenta. Additionally, the amplicon had complete identity with short sequences of asinine herpesviruses that have been published in association with interstitial pneumonia in donkeys. EHV-7 has previously been isolated from nasal secretions of normal donkeys and mules. Our report describes a case of abortion associated with EHV-7 or a similar virus.
Gammaherpesviruses are lymphotropic viruses that are characterized as slow-growing in vitro. There are several important human gammaherpesviruses, such as Human herpesvirus 4 (Epstein–Barr virus) and Human herpesvirus 8 (Kaposi’s sarcoma–associated herpesvirus). In contrast, the role of gammaherpesviruses in disease of equids is less obvious. Equid herpesvirus 2 and 5 (EHV-2 and -5) are gammaherpesviruses that infect horses, but because they can be isolated in the absence of disease, their true pathogenic potential is unclear. However, EHV-2 has been isolated from juvenile and adult horses with upper respiratory tract disease and conjunctivitis; the virus has also been associated with development of pneumonia.2,8,10 Gammaherpesvirus infections have also been reported in donkeys and mules: A virus similar to asinine herpesvirus 5 was isolated from a pharyngeal swab of a donkey with neurologic disease, 15 and asinine herpesviruses 4 and 5 were identified in donkeys with interstitial pneumonia. 11 Another gammaherpesvirus, Equid herpesvirus 7 (EHV-7; order Herpesvirales, family Herpesviridae, subfamily Gammaherpesvirinae, genus Unassigned), also known as asinine herpesvirus 2 (AHV-2), has been isolated from the blood of a healthy donkey and from the nasal secretions of a mule following an outbreak of respiratory disease.1,3 Unfortunately, sequence information is not available for the first AHV-2 described in 1988, which was isolated from a healthy donkey, but produced rhinitis when 2 weanling donkeys were inoculated with the virus. 3 A small survey found that EHV-7 was recovered from the nasal secretions of ~8% of healthy mules (n = 114) and donkeys (n = 13). 1 We describe a third trimester abortion in a Mediterranean miniature donkey (Equus asinus) from which a gammaherpesvirus similar to EHV-7 was isolated from the placenta. The microscopic lesions are unique among herpesvirus abortions in equids and, to our knowledge, placentitis and abortion has not been described in association with EHV-7 or any similar gammaherpesviruses in equids.
In April 2014, a 3.9-kg, preterm, female Mediterranean miniature donkey fetus was submitted for autopsy to the Animal Disease and Food Safety Laboratory (Sparks, Nevada). The dam was reported to be current on vaccines and was 1 of 11 equids on a property that housed horses, miniature donkeys, and miniature zonkeys (zebra–miniature donkey hybrids). The dam previously had a full-term abortion 2 years prior. The previous abortion occurred on an adjacent property while the jenny was housed with a pony, now deceased. There were no other reproductive losses on either farm. The fetus was submitted along with an intact allantoamnion and allantochorion. The stage of pregnancy was estimated to be early to mid-third trimester. No gross lesions were noted on the fetus or the allantoamnion. The allantochorion was covered with multifocal, small, yellow-gray regions of altered villi with a gritty consistency; these villi grossly appeared necrotic. Formalin-fixed tissues and fresh, frozen placenta were submitted to the Washington State University, Washington Animal Disease Diagnostic Laboratory (Pullman, Washington) for additional diagnostic evaluation.
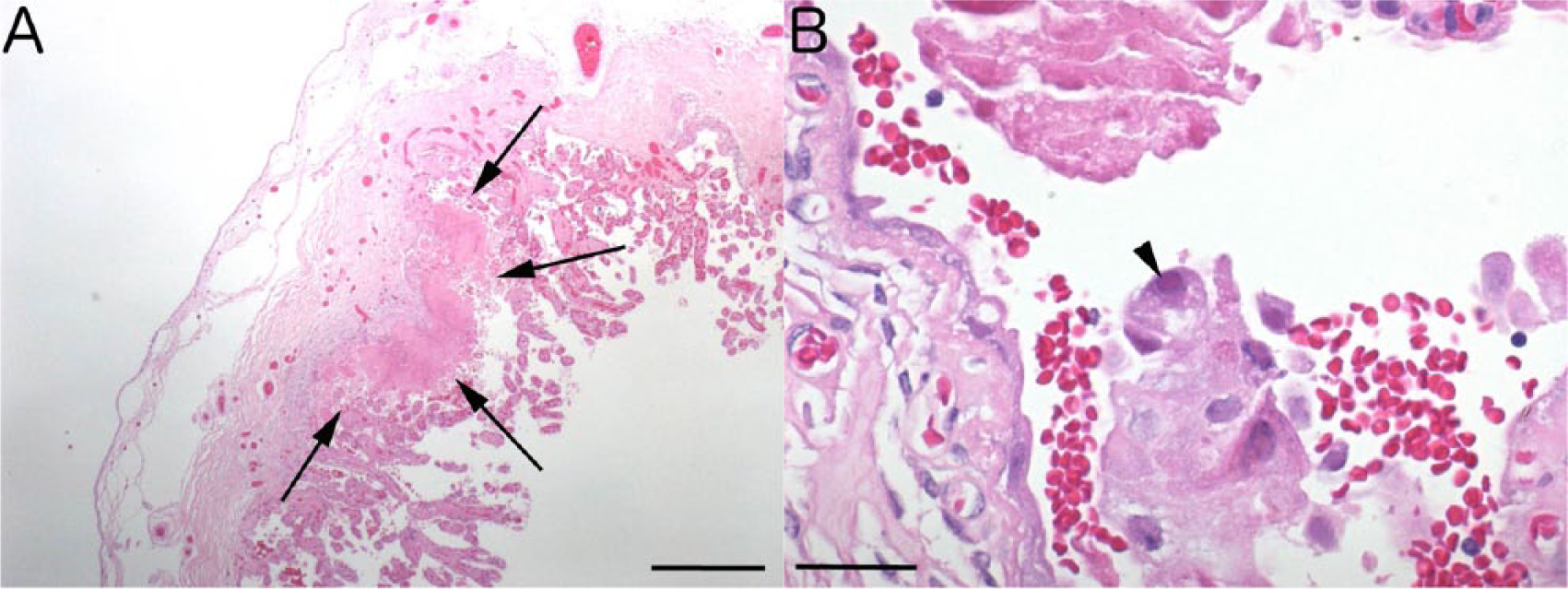
Microscopic evaluation of the placenta revealed scattered hypereosinophilic foci within the chorionic villi accompanied by loss of cellular and nuclear detail (Fig. 1). Chorionic stroma subjacent to these foci was occasionally infiltrated with small numbers of lymphocytes and macrophages. Occasional cells within and adjacent to the necrotic foci had eosinophilic, intranuclear inclusions (Fig. 1B). The fetal heart, spleen, liver, kidney, lung, thymus, ovary, and umbilicus were examined and were considered microscopically normal.
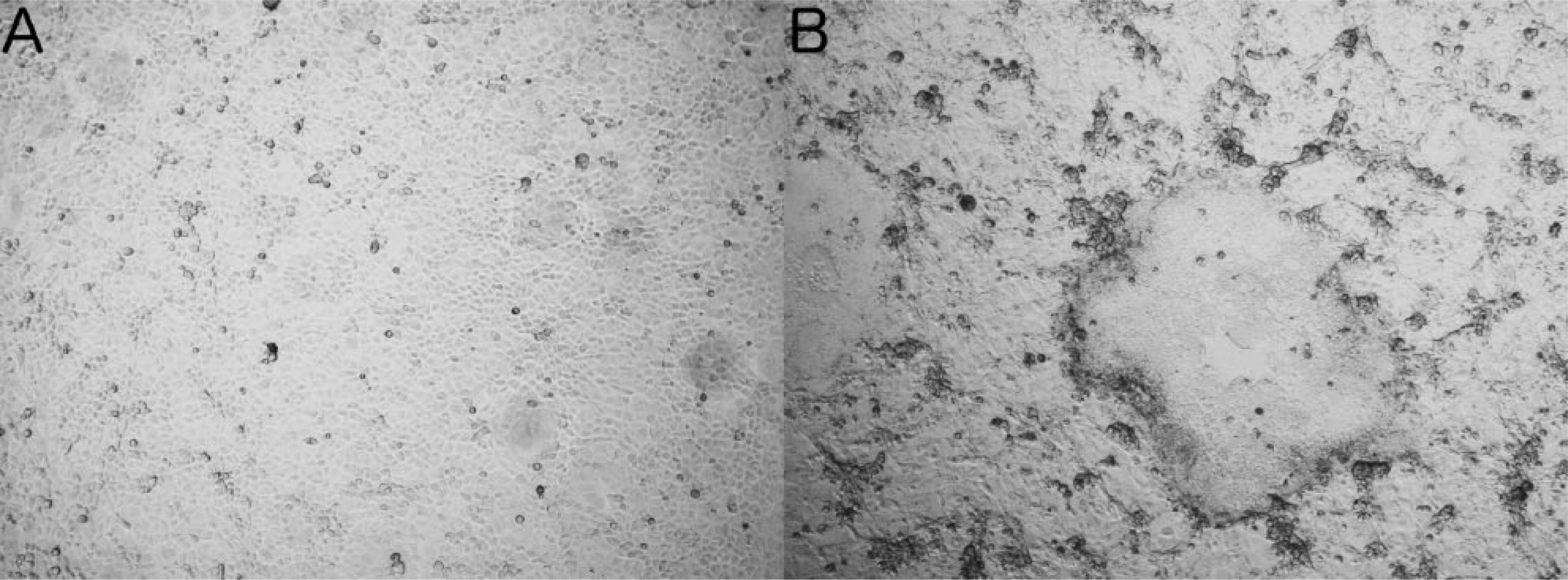
Based on the placental lesions, virus isolation was performed on frozen, fresh placenta using standard detection techniques on multiple cell lines. 4 Processed tissue supernatant was inoculated onto confluent monolayers of rabbit kidney–derived (RK)-13B and primary equine kidney cells. Positive controls consisted of Equid herpesvirus 1 (EHV-1) in parallel cell cultures. Negative controls consisted of cell culture media (minimum essential media) without virus. After day 5 postinoculation, the cell cultures were passaged, and the second passage was observed for 5–10 days. Subsequent passage was performed only on RK-13B cell cultures because demonstrable cytopathic effect (CPE) was observed (Fig. 2B). Negative control cell lines did not show any CPE (Fig. 2A). The CPE on infected RK-13B cells was progressive over 5–7 days and was characterized by syncytia with enlarged, rounded, and refractile cells arranged in loose clusters (Fig. 2B). Infected RK-13B cells had a propensity to exhibit CPE on passage quite early, within the first week postinoculation, whereas no CPE was observed in the infected equine kidney cells. Chloroform sensitivity was performed at passage 3 by mixing CHCl3 with viral cell culture supernatant at a 1:1 ratio for 10 min at room temperature. Treatment with chloroform prevented CPE on RK-13B cells, indicating that the virus was enveloped. The viral isolate was identified as WADDL 2014-5403.
Supernatant from the virus amplified in RK-13B cells showing CPE was prepared for ultrastructural analysis using standard electron microscopic negative stain. 7 Examination of uranyl acetate–stained grids revealed roughly hexagonal nucleocapsids ~100 nm in diameter, surrounded by electron-sparse material interpreted as envelope, consistent with herpesvirus ultrastructural morphology 5 (Fig. 3).
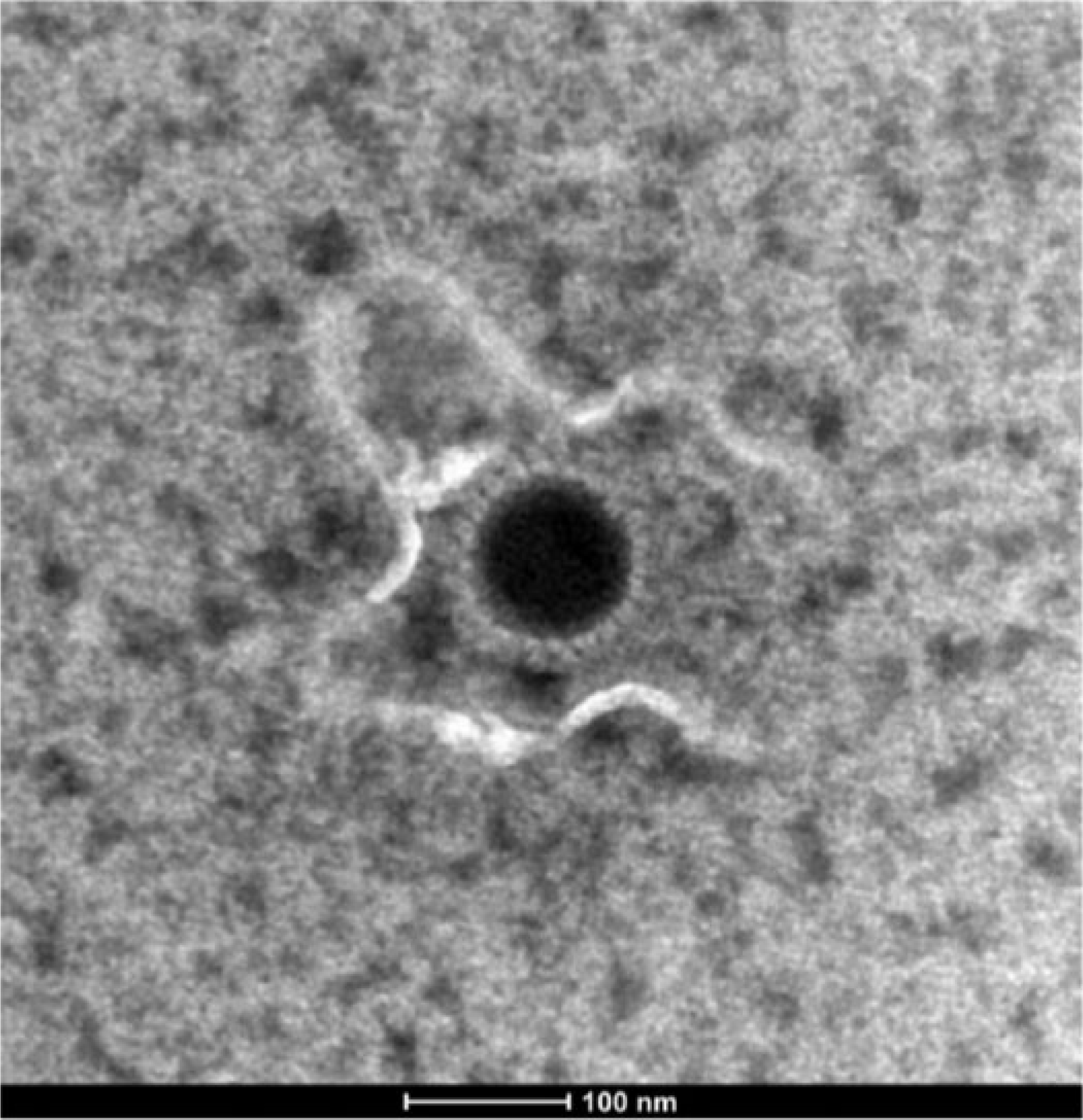
Electron photomicrograph of the virus isolated from the placenta of a miniature donkey abortion. Nucleocapsid is surrounded by electron-sparse material, consistent with a viral envelope. Uranyl acetate stain. Bar = 100 nm.
The supernatant fluids from infected RK-13B cell cultures were subjected to polymerase chain reaction (PCR) followed by sequencing. Specifically, DNA was extracted using a commercial kit following manufacturer directions. a A single-round PCR for amplification of a portion of the DNA polymerase gene was performed using herpesvirus-specific degenerate primers as previously described. 14 Positive results were obtained from the infected cell line as compared with the negative control, and a 699–base pair (bp) sequence was obtained from the infected RK-13B cell culture DNA extraction. Sequences were compared with data available in the National Center for Biotechnology Information database (GenBank). The partial sequences of the herpesvirus DNA polymerase gene clustered with equid gammaherpesviruses (Fig. 4). 6 The virus had 96.7% sequence similarity to a portion of the DNA polymerase gene from EHV-7 (GenBank accession EU165547) and 98–100% sequence similarity to short (166 bp) sequences of the DNA polymerase gene of asinine herpesvirus 4 (GenBank accession AY054991.1, AY054992.1, AY054990.1). Sequence similarity, along with moderately high (≥95%) sequence coverage, was 91% to asinine herpesvirus 5 (GenBank accession FJ98319.1), 89% to EHV-5 (GenBank accession JX125459.1), and 87% to EHV-2 (GenBank accession HQ247790.1). EHV-1 and -4 were not detected via PCR, using specific EHV-1 and EHV-4 primers on infected RK-13B cell culture supernatant. An identical sequence was obtained when the same degenerate herpesvirus primers were used for PCR on the formalin-fixed, paraffin-embedded placenta. DNA was extracted from the formalin-fixed, paraffin-embedded placenta using a commercial kit b as previously described. 16 The sequence obtained for the WADDL 2014-5403 herpesvirus was deposited into GenBank as accession KP245904.
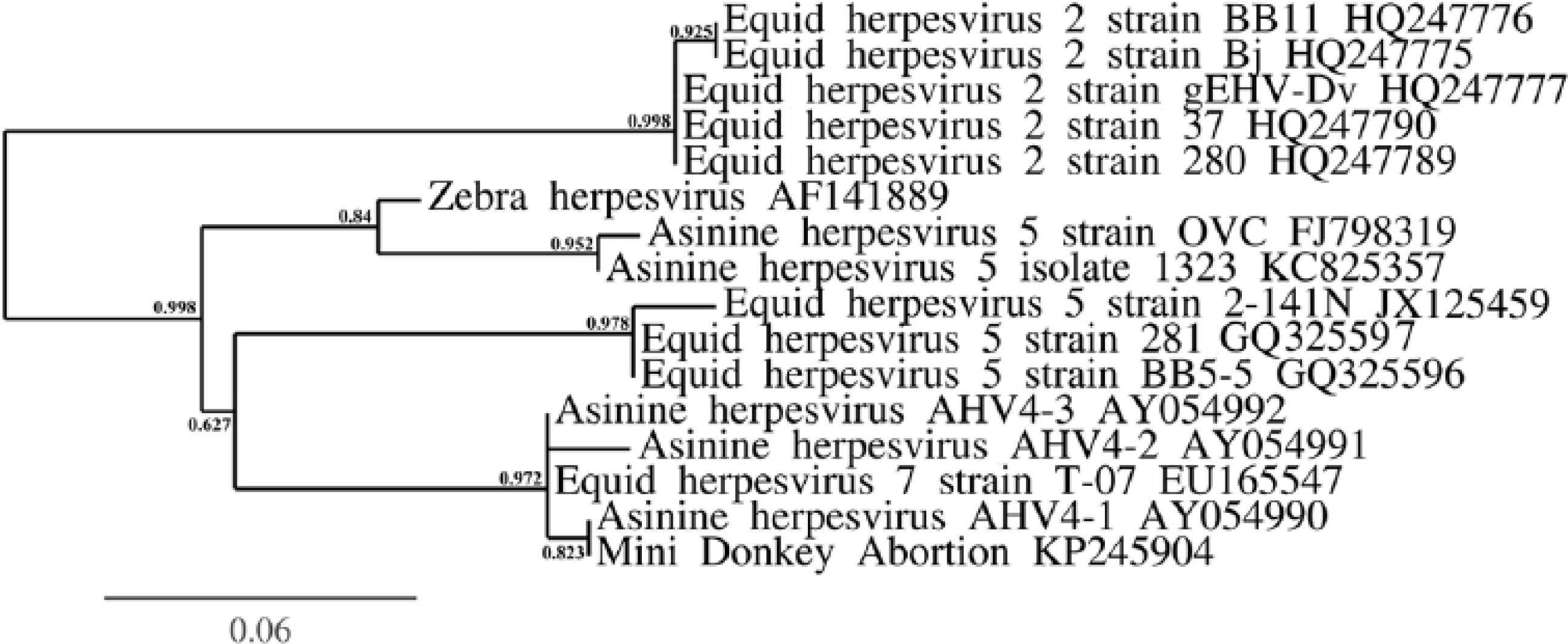
Phylogenetic relationship, using the approximate likelihood ratio test for branches, 4 between the abortion-associated WADDL 2014-5403 herpesvirus (listed as “mini donkey abortion”) and the other viruses to which it had greatest nucleotide identity (all equine gammaherpesviruses).
Because the sequence similarity level between the virus reported herein and EHV-7 was only 96.7%, this suggested that the virus isolated from the placenta of this miniature donkey abortion may represent a new species of equid herpesvirus, the sequence of which has not been previously reported. However, it is also known that there is sequence heterogeneity within many of the gammaherpesviruses, so the virus isolated in our study may simply be a variant of EHV-7. 13 While there was a high degree of similarity to sequences from the DNA polymerase gene of asinine herpesvirus 4, the banked sequences of AHV-4 are too short (only 166 bp) to imply that the virus described is AHV-4. In fact, in the report of EHV-7 isolated from a healthy mule, it was also noted that a portion of the DNA polymerase gene had 100% sequence identity to a short portion of AHV-4. 1
Approximately 4 months after the abortion occurred, sera were collected from the dam of the aborted fetus along with 7 herd mates. All the miniature donkeys and horses on the property were sampled (miniature donkey–zebra hybrids could not be sampled), and microtiter virus neutralization assays were performed in order to detect virus-neutralizing antibody against EHV-1 and -4, EHV-3, Equine arteritis virus (EVA), and the virus isolated from the placenta of the aborted miniature donkey fetus (WADDL 2014-5403 herpesvirus). Tests for known viruses were performed as described in the OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 17 EHV-1 and -4 and EVA assays were performed using RK-13B cells, and the EHV-3 assay was performed using primary equine kidney cells. Positive controls and virus stocks were obtained from the National Veterinary Services Laboratory. c The microtiter virus neutralization assay for WADDL 2014-5403 herpesvirus was performed on RK-13B cells using a general protocol for herpesvirus virus neutralization assays. 17 The 50% tissue culture infective dose (TCID50) of WADDL 2014-5403 herpesvirus was determined by incubating serial dilutions of virus on pre-seeded RK-13B cells and observing for CPE. 9 The Spearman–Kärber method 9 was used to calculate TCID50, which was 104.125/50 µL. All animals were negative for neutralizing antibody against EHV-3 at 1:4 or 1:8 (serum cytotoxicity observed at 1:4 in 2 animals) and negative at 1:4 for EVA virus-neutralizing antibodies. All animals showed evidence of neutralizing antibody against EHV-1 and -4, but titers did not correlate with neutralizing antibody titers against WADDL 2014-5403 herpesvirus (Table 1). Four miniature donkeys had evidence of virus-neutralizing antibody against WADDL 2014-5403 herpesvirus, and all infected animals were genetically related to the dam of the aborted fetus, with the exception of the sire of the aborted fetus. The horses and other non-related miniature donkeys on the property did not have evidence of infection with WADDL 2014-5403 herpesvirus based on absence of detectable virus-neutralizing antibody. The infection pattern in this herd is consistent with a familial pattern of virus infection even though there is ample opportunity for nose-to-nose contact between all animals on the property.
Familial distribution of virus-neutralizing antibody against the abortion-associated WADDL 2014-5403 virus and unrelated Equid herpesvirus 1 and 4 (EHV-1 and -4, respectively; TCID50 = 102.625/50 µL). Two independent virus neutralization assays were performed using WADDL 2014-5403 herpesvirus, one at a lower concentration of virus (100 TCID50) and one at a higher concentration (1,000 TCID50).*
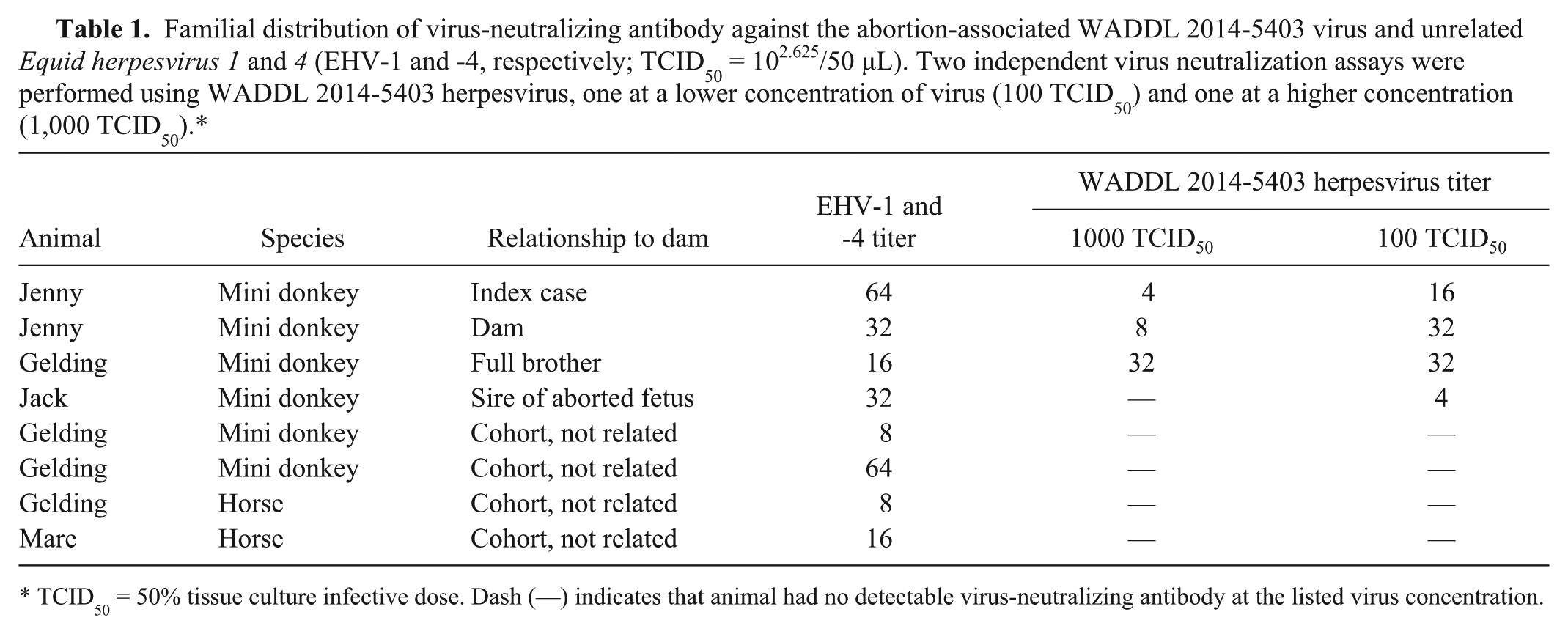
TCID50 = 50% tissue culture infective dose. Dash (—) indicates that animal had no detectable virus-neutralizing antibody at the listed virus concentration.
The microscopic lesions reported in our study are novel for herpesvirus-associated abortion in an equid. Alphaherpesviruses, such as EHV-1, are well-known abortifacient viruses in horses. Histologically, the placenta is not affected, and typical microscopic lesions include multifocal necrosis of the fetal lung, liver, spleen, and kidney, often accompanied by viral intranuclear inclusion bodies and surrounded by mononuclear inflammation. This is in sharp contrast to the placentitis observed in this abortion without any accompanying lesions in the fetal internal organs. In fact, examination of placenta was required in order to make this diagnosis, which is important to note when examining tissues from other donkey abortions submitted to veterinary diagnostic laboratories. Although the gammaherpesviruses EHV-2 and EHV-5 have been infrequently isolated from aborted equine fetal tissues (0.75% and 0.25% incidence, respectively), the role of these viruses in the pathogenesis of equid abortion remains unclear. 12
Evidence suggests that the etiologic agent associated with the miniature donkey abortion described herein is a novel gammaherpesvirus similar to EHV-7, as supported by isolation of the virus from the placenta, and histologic observation of intranuclear viral inclusion bodies, which are compatible with herpesvirus inclusions, that accompanied placental necrosis. Additional work will be required to further compare this potentially novel virus to EHV-7, and to better understand the association between this virus and fetal loss in donkeys and other equids.
Footnotes
Acknowledgements
We thank Lorraine Tanaka, Jessica Schiflett, Jennifer Baisley, and Dr. Christine Davitt for technical assistance.
Authors’ contributions
A Rink contributed to conception of the study and acquisition of data. JF Evermann and GJ Haldorson contributed to conception and design of the study. TE LeCuyer, DS Bradway, and T Baszler contributed to design of the study. TE LeCuyer, DS Bradway, JF Evermann, and GJ Haldorson contributed to acquisition, analysis, and interpretation of data. AV Nicola and T Baszler contributed to interpretation of data. TE LeCuyer drafted the manuscript. A Rink, DS Bradway, JF Evermann, AV Nicola, T Baszler, GJ Haldorson critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
a.
Ambion MagMax AM1836TM-96 viral isolation kit, Thermo Fisher Scientific Inc., Waltham, MA.
b.
QIAamp DNA minikit, Qiagen Inc., Valencia, CA.
c.
070-EDV equine viral arteritis virus, 370-EDV EVA antiserum, 040-EDV EHV-1 virus, 340-EDV EHV-1 antiserum, 042-EDV EHV-3 virus, 342-EDV EHV-3 antiserum; National Veterinary Services Laboratories, Ames, IA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research was funded by teaching funds provided by the Department of Veterinary Microbiology and Pathology and by the Washington Animal Disease Diagnostic Laboratory, Washington State University. Dr. Tessa LeCuyer is supported by an Achievement Rewards for College Scientists Fellowship.