
Abstract
The primary objective of our study was to optimize detection of serum antibodies to Borrelia burgdorferi using a new commercial automated fluorescence system (Accuplex4 BioCD system, Antech Diagnostics, Lake Success, New York). The system used multiple natural and artificial peptides—outer surface proteins (OspA, OspC, OspF), an outer membrane protein (P39), and a proprietary synthetic peptide (small Lyme peptide [SLP])—and the results were compared with a commercially available enzyme-linked immunosorbent assay that uses a proprietary peptide (C6). Sera from 4 groups were evaluated: dogs vaccinated with 1 of 3 commercially available vaccines (n = 18); dogs infested with adult Ixodes scapularis (black-legged tick; n = 18); dogs previously vaccinated and then infested with I. scapularis (n = 18); and dogs with B. burgdorferi infection that were then vaccinated (n = 14). All of the vaccines evaluated induced OspA responses. However, antibodies against OspF or C6 were not induced in any of the vaccinated dogs. Additionally, the OspF antibodies had 100% sensitivity and specificity when compared to antibodies against C6 peptide. In B. burgdorferi–infected dogs, antibodies against OspC and SLP were detected in serum sooner than antibodies against the other targets. Low levels of antibodies against OspA developed in 6 of 14 B. burgdorferi–infected, unvaccinated dogs and had the shortest duration compared to the other antibodies. Detection of antibody responses to multiple B. burgdorferi targets with this system can be used to help differentiate vaccinated dogs from exposed dogs as well as acute infection from chronic infection.
Introduction
Borrelia burgdorferi infection in dogs in the United State can be extremely common in areas endemic for Ixodes spp.3–5,12 For example, in one study, the seroprevalence rates for dogs residing in Connecticut and Wisconsin were 18.1% (1,846 positive tests from 10,209 samples tested) and 10.2% (6,018 positive tests from 59,070 samples tested), respectively. 4 As B. burgdorferi infection is an important cause of clinical illnesses, such as polyarthritis and nephritis, the ability of veterinarians to make an accurate diagnosis is important.7,12
Multiple serological test methods including indirect fluorescent antibody test, enzyme-linked immunosorbent assay (ELISA), and Western blot immunoassay (WB) have been assessed using a number of different B. burgdorferi target antigen preparations. 8 In an article published in 2011, a bead-based multiplex assay for detection of antibodies to outer surface proteins A, C, and F (OspA, OspC, and OspF, respectively) was described, indicating the use of OspC as an early marker of natural infection and OspF as a later stage marker of natural infection. 17 Detection of antibodies to OspA in the WB and bead assays has previously been associated with response to commercially available vaccines.10,17 However, it was then documented that antibodies to OspA may occur in infected nonvaccinated dogs but the response is transient and lower than noted in vaccinated dogs. 18 Additionally, a commercially available vaccine a induces antibodies to both OspA and OspC, complicating differentiation of natural exposure and vaccinal response in the first few weeks of natural infection or a few weeks postvaccination. 10 Many antigen preparations used in B. burgdorferi serological assays cannot be used to discriminate antibodies against natural exposure and postvaccinal antibodies. 12 However, some peptides, including the C6 peptide used in one commercially available kit (SNAP), b rarely detect antibodies against vaccinal antigens. 12
Humoral immune responses against B. burgdorferi infection or vaccination are robust, with differing responses to different targets over time following exposure or vaccination. The B. burgdorferi component of the SNAP4DX b consists of only the C6 peptide. Automated antibody detection systems have progressed over the last several years with systems now available that can measure responses against multiple antigens using small volumes of serum. 17 The commercial automated fluorescent system, the Accuplex4 BioCD system, c used in our study can be used to assess antibody responses to multiple antigens using 70 µL of serum.14,15 The system is a novel, silicon disc substrate multiplex platform for detection of peptides and antibodies, using a single serum or plasma sample. The use of the assay for detection of antibodies against peptides of Anaplasma phagocytophilum and Ehrlichia canis was previously reported.14,15
The primary objective of the current study was to optimize detection of serum antibodies against multiple natural and artificial peptides of B. burgdorferi for use on the Accuplex4 system platform. The discs also contain peptides for detection of antibodies against natural and artificial peptides of A. phagocytophilum and E. canis, as well as an antigen of Dirofilaria immitis. After optimization, the secondary objective was to apply the assays to sera collected from dogs exposed to B. burgdorferi–infected Ixodes scapularis (black-legged ticks) before or after immunization with 3 of the market-leading commercial vaccines against B. burgdorferi. The primary hypothesis was that the combination of multiple antibody responses can be used to differentiate natural exposure to B. burgdorferi from vaccination.
Materials and methods
Animals
This study was approved by the Institutional Animal Care and Use Committee at Colorado State University. The Beagles (n = 18 male and 18 female) used in our study were from a vector-free laboratory animal facility and ranged in age from 12 to 13 months at the beginning of the experiments. On arrival at the Colorado State University research facility (Fort Collins, Colorado), the males were neutered using the standard operating procedures in the Small Animal Anesthesia and Soft Tissue Surgery Services. The dogs were housed in elevated runs and fed ad libitum. Daily animal care was provided by the staff of Laboratory Animal Resources. Except during times of tick placement, university student volunteers walked the dogs and provided socialization in preparation for adoption to private homes or transfer to other studies at the end of the experiments described herein.
Experimental design
Experiment 1: Serological responses of adult dogs after administration of B. burgdorferi–containing vaccines
A total of 18 Beagles were randomly selected to be administered 1 of 3 market-leading vaccines (3 male and 3 female per vaccine).a,d,e The vaccines were administered on weeks 0 and 4, and the dogs were observed daily for clinical signs of disease in a vector-free facility with blood samples collected twice after arrival at the research facility prior to vaccination and then weekly for 20 weeks.
Experiment 2: Serological responses of nonvaccinated adult dogs after exposure to I. scapularis
A total of 18 Beagles (n = 9 male and 9 female) were randomly selected to be infested with I. scapularis prior to vaccination. Adult I. scapularis f were placed on all 18 dogs in chambers f for 1 week as described below. The ticks were removed, an acaricide g applied, and the dogs were observed daily for clinical signs of disease, with blood samples collected twice after arrival at the research facility prior to tick infestation and then weekly for 12 weeks. Two skin biopsies (6 mm) were collected from the I. scapularis infestation sites 60 days after infestation and evaluated at a service laboratory f for B. burgdorferi DNA by a polymerase chain reaction (PCR) assay. 13 The laboratory utilized primers A2 [5′-GTTTTGTAATTTCAACTGCTGACC-3′] and A4 [5′-CTGCAGCTTGGAATTCAGGCACTTC-3′].
Experiment 3: Serological responses of B. burgdorferi–vaccinated adult dogs after exposure to I. scapularis
Blood samples were collected from each of the dogs from experiment 1, 22 weeks after the first immunization, and then adult I. scapularis f were placed on all 18 dogs in chambers for 1 week as described below. The ticks were then removed, an acaricide g applied, and the dogs observed daily for clinical signs of disease with blood samples collected weekly for 16 weeks. Two skin biopsies (6 mm) were collected from the I. scapularis infestation sites 60 days after infestation and evaluated for B. burgdorferi DNA by PCR assay. f
Experiment 4: Serological responses of B. burgdorferi–infected adult dogs after administration of B. burgdorferi–containing vaccines
Dogs infected with B. burgdorferi in experiment 2 were administered 1 of 3 market-leading vaccines (3 male and 3 female per vaccine).a,d,e The vaccines were administered on weeks 0 and 4 of this experiment, and the dogs were observed daily for clinical signs of disease with blood samples collected weekly for 7 weeks.
Tick infestation
Adult I. scapularis wild-caught in Rhode Island in March 2010 were purchased. f The prevalence rate of B. burgdorferi DNA in a representative aliquot of adult ticks from the capture area was ~50% as assessed by PCR 13 on tick extracts performed by a service laboratory. f The adult ticks were maintained at room temperature in humidified chambers until used. To aid in manipulation of the tick chambers, the dogs were placed under heavy sedation during placement. When placed on the dogs, 13 female and 12 male ticks were allowed to attach under a tick chamber made of adhesive bandage materials. A collar h that blocked the dogs from removing the tick chamber was placed until day 7 when the ticks were removed.
Borrelia burgdorferi targets
The B. burgdorferi peptides used in our study included OspA, OspC, OspF and an outer membrane protein (P39).16–18 The peptides OspA, OspC, OspF, and P39 were sourced from different laboratories.i–l In addition, a proprietary synthetic peptide derived from components of a Borrelia flagellar protein and an outer surface protein, termed small Lyme peptide (SLP), was used. The expression of these proteins was conducted via commercial suppliers.m,n In brief, the genes coding for these peptides were cloned into LIC-pEV-L8 o and pET-46 Ek/LIC p expression vectors according to the manufacturer’s protocol. The fusion proteins contain an N-terminal cleavable His-Tag sequence. The resulting expression constructs were transformed into Escherichia coli BL21 (DE3) pLysS according to the manufacturer’s protocol. The PCR products and plasmids were sequenced at a core facility. p
To express the recombinant protein, the transformed E. coli strain was grown in lysogeny broth medium in the presence of the appropriate selective markers at 37°C until the optical density (at 600 nm) reached 0.8. The cell culture was induced with isopropyl-β-
Purified recombinant protein fractions were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis. s Gels were stained following the manufacturer’s instructions. t Western blot analyses were performed using a 1:1,000 dilution of monoclonal mouse antibodies against histidine that were conjugated to horseradish peroxidase, u and a chromogenic detection system v was used according to the manufacturer’s recommendations. The purified recombinant proteins were then used for ELISA and Accuplex4 system applications.
The Accuplex4 system
The system was optimized for the B. burgdorferi peptides OspA, OspC, OspF, P39, and SLP as described for E. canis. 15 Positive and negative control sera used in assay titrations were obtained from the dogs in experiment 2 before (negative control) and after (positive control) infestation with I. scapularis. The optimal peptide concentrations were determined by assessing optimal signal-to-noise ratio, varying printed target concentrations, and buffer compositions. The cut-point for a positive test result for each antibody assay was determined by assay of serum from dogs with known infection status. Change in signal over time and comparison with results of existing assays (Western blot immunoassay and the SNAP kit b ) were used to establish static cutoff points.
The intra-assay variation of the assay was calculated by determining the mean and standard deviation for the fluorescent counts for 20 positive control samples that were just above the positive cutoff value and then calculating the coefficient of variation. This experiment was performed with the same positive control samples on separate discs on 3 consecutive days. The interassay variation was determined by comparing the coefficient of variations between the 3 discs.
Other assay
In this study, the sera from the vaccinated and I. scapularis–infested dogs were assessed in both the optimized Accuplex4 system as described and the SNAP kit following the manufacturer’s instructions.
Statistical evaluation
Borrelia burgdorferi infection was diagnosed in dogs that had specific DNA amplified from skin biopsies or in dogs that had antibodies detected in >3 serum samples collected after I. scapularis infestation by the SNAP kit. Sensitivity and specificity for some of the target antibodies were determined as indicated by comparing with results for antibodies against the C6 peptide, the basis of the SNAP kit. The time to first positive test result was compared for some target antibodies using the Wilcoxon rank sum test. Significance was defined as P < 0.05.
Results
Preinfestation or vaccination serological assay results
All dogs were shown to be negative for antibodies against B. burgdorferi (C6 peptide), A. phagocytophilum, E. canis, and antigens of D. immitis by the SNAP kit in sera collected prior to shipping to Colorado State University and in sera collected 2 weeks apart prior to being vaccinated with B. burgdorferi vaccines or infested with I. scapularis.
Assay titration experiments
The interassay variations for antibody responses against OspA, OspF, P39, SLP, and OspC targets were 6.1%, 6.6%, 8.1%, 5.3%, and 6.5%, respectively. The intra-assay variations for antibody responses against OspA, OspF, P39, SLP, and OspC targets were 10.7%, 9.2%, 9.4%, 10.4%, and 12%, respectively.
Clinical findings
Greater than 90% of the adult I. scapularis attached and fed on all dogs during the 7-day feeding period. The Beagles were evaluated daily for signs of depression, inappetence, or lameness. Clinical signs of disease that could be related to vaccination were not noted in any of the dogs. Clinical signs of disease consistent with fever or polyarthritis were not noted in any of the dogs over the duration of the study.
Borrelia burgdorferi and Anaplasma phagocytophilum infections
No dogs had antibodies against B. burgdorferi or A. phagocytophilum antigens prior to exposure to I. scapularis. After I. scapularis infestation in experiment 2 (I. scapularis infestation prior to vaccination), 14 dogs (77.8%) developed evidence of B. burgdorferi infection and 10 dogs (55.6%) developed evidence of A. phagocytophilum infection. A total of 13 out of 14 infected dogs had B. burgdorferi DNA amplified from skin, and the remaining dog was positive for antibodies against the C6 peptide and other specific antigens in the Accuplex4 system on >3 sample dates after I. scapularis infestation. After I. scapularis infestation in experiment 3 (vaccination prior to I. scapularis infestation), 6 dogs (33.3%) developed evidence of B. burgdorferi infection based on the presence of specific DNA in skin biopsies indicating vaccine failure. Additionally, 12 dogs (66.7%) in experiment 3 developed evidence of A. phagocytophilum infection. Overall, 20 of 36 dogs (55.6%) developed evidence of B. burgdorferi infection, and 22 of 36 (61.1%) dogs developed evidence of A. phagocytophilum infection.
Experiment 1: Serological responses of adult dogs after administration of B. burgdorferi–containing vaccines
None of the vaccinated dogs developed antibodies against OspF or C6. In contrast, all 18 of the dogs administered 1 of 3 vaccines prior to infestation with I. scapularis developed antibodies against OspA (Fig. 1). The first positive test result occurred at week 1 (1 dog), week 2 (14 dogs), week 3 (1 dog), and week 5 (2 dogs) with a median of 2 weeks (Table 1). After the first positive result, 16 dogs were positive for OspA antibodies for 20 weeks; OspA titers waned in 1 dog after week 11 and another dog after week 12. Antibodies against P39 (7 dogs), SLP (9 dogs), and OspC (9 dogs) were detected less frequently than those against OspA and were generally of short duration (Fig. 1; Table 1). Antibodies against OspC were first detected at week 1 in all 6 dogs vaccinated with the vaccine containing OspC a and at week 5 for 3 dogs vaccinated with the other bacterin. d The OspC antibodies were no longer detected after week 5 (1 dog), week 7 (1 dog), week 8 (1 dog), week 11 (1 dog), week 12 (2 dogs), week 13 (1 dog), or week 14 (1 dog).
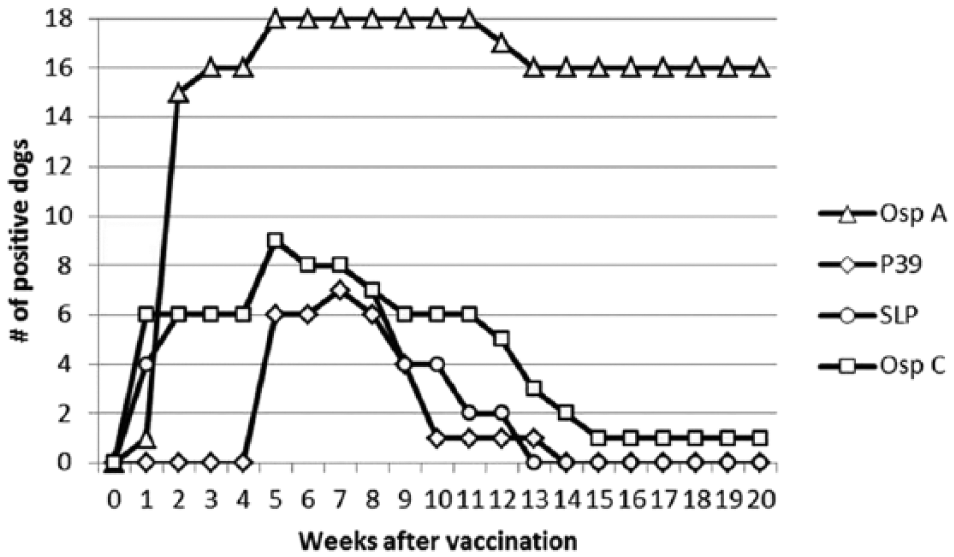
Serum antibody responses to Borrelia burgdorferi antigens after vaccination with 1 of 3 commercially available vaccines (6 dogs per vaccine).a,d,e Vaccines were administered at weeks 0 and 4. None of the dogs developed antibodies against outer surface protein F (OspF) or C6. See Table 1 for further results.
Distribution of serum antibody results against 6 Borrelia burgdorferi targets in serum of 18 vector-free dogs vaccinated with 1 of 3 vaccines that are commercially available in the United States.*
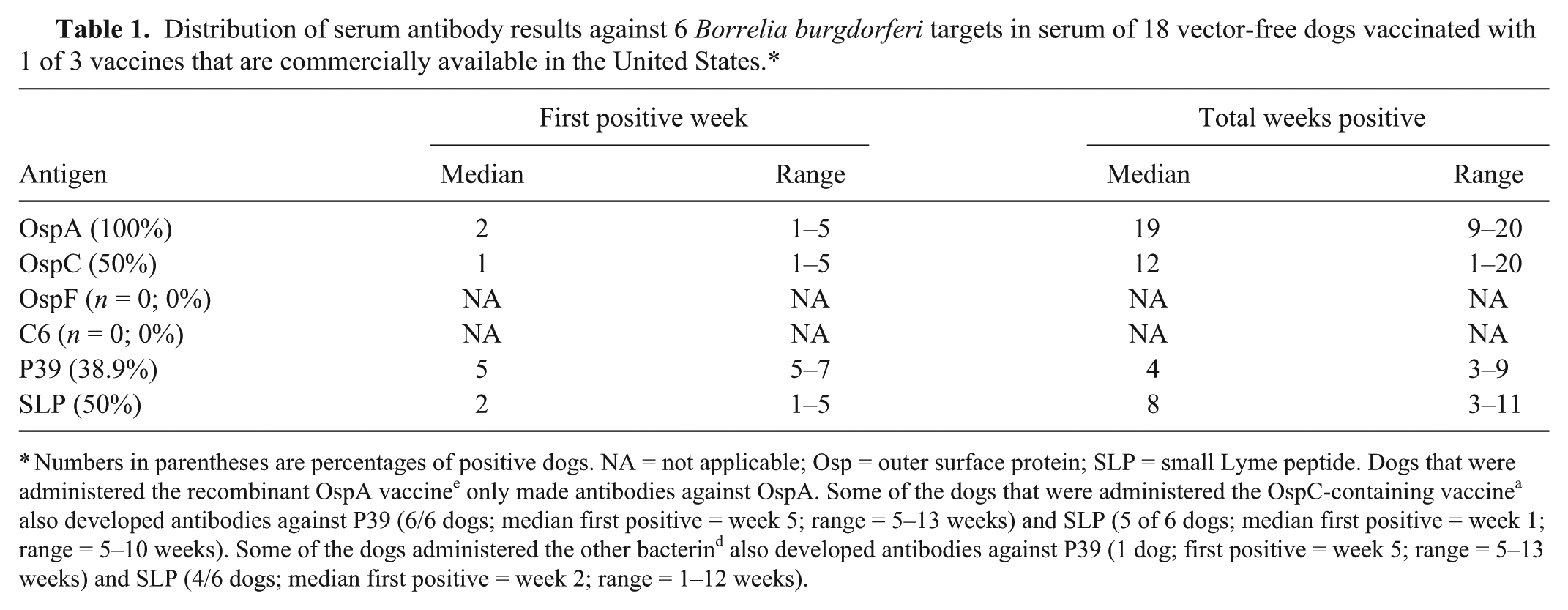
Numbers in parentheses are percentages of positive dogs. NA = not applicable; Osp = outer surface protein; SLP = small Lyme peptide. Dogs that were administered the recombinant OspA vaccine e only made antibodies against OspA. Some of the dogs that were administered the OspC-containing vaccine a also developed antibodies against P39 (6/6 dogs; median first positive = week 5; range = 5–13 weeks) and SLP (5 of 6 dogs; median first positive = week 1; range = 5–10 weeks). Some of the dogs administered the other bacterin d also developed antibodies against P39 (1 dog; first positive = week 5; range = 5–13 weeks) and SLP (4/6 dogs; median first positive = week 2; range = 1–12 weeks).
Experiment 2: Serological responses of nonvaccinated adult dogs after exposure to I. scapularis
Borrelia burgdorferi DNA that was amplified from skin biopsies or antibodies against C6 peptide were detected in 14 of 18 dogs confirming B. burgdorferi infection. The other 4 dogs did not develop antibodies to any of the 6 B. burgdorferi targets and did not have B. burgdorferi DNA in skin biopsies and so were not analyzed further. Of the 14 unvaccinated dogs that developed B. burgdorferi infection after I. scapularis infestation, 14 developed antibodies against OspF and P39, 13 developed antibodies against SLP and OspC, and 6 developed low levels of antibodies against OspA (Fig. 2; Table 2). While not compared statistically, fluorescent counts for OspA antibodies induced by B. burgdorferi infection were generally lower than those of dogs administered vaccines. The time to first positive test result was fastest for OspC and SLP compared with each of the other targets (P < 0.05) with a median of 3 weeks for both targets. In addition, antibodies against the C6 peptide were detected sooner than antibodies against OspF. The shortest duration of antibodies was for those against OspA. However, there was overlap for time to first positive results as well as the number of positive weeks for all antigens in some dogs (Table 2).
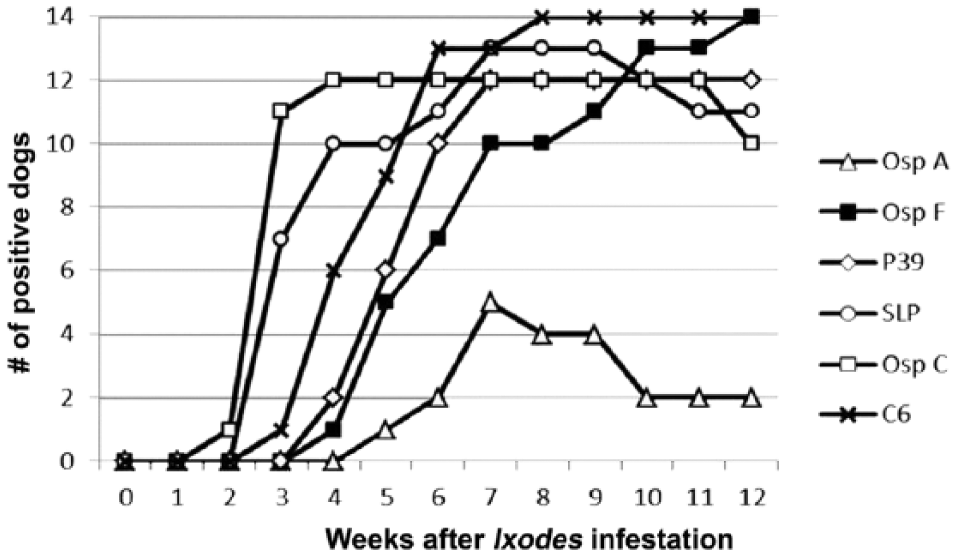
Serum antibody responses to 6 different Borrelia burgdorferi antigens in 14 dogs infected by exposure to wild-caught Ixodes scapularis. Fluorescent counts associated with outer surface protein A (OspA) were numerically lower than those associated with vaccination (Fig. 1). See Table 2 for further detailed results. Antibodies against an antigen were not always detected in all dogs on a given day.
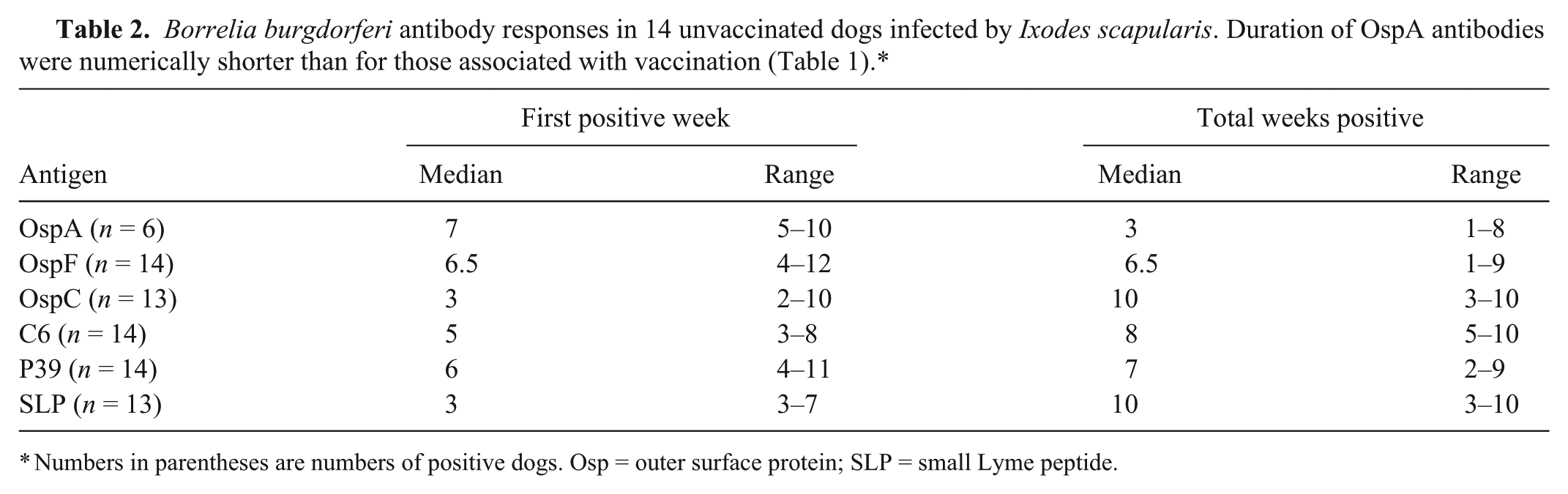
Numbers in parentheses are numbers of positive dogs. Osp = outer surface protein; SLP = small Lyme peptide.
Experiment 3: Serological responses of B. burgdorferi–vaccinated adult dogs after exposure to I. scapularis
At the time of I. scapularis infestation (22 weeks after the first vaccine administration), 16 of 18 dogs had, in experiment 1, antibodies to OspA, and 1 of 18 dogs had antibodies to OspC; all other antibody test results were negative. After tick infestation, 6 of 18 dogs had B. burgdorferi DNA amplified from skin biopsies, documenting B. burgdorferi infection. These 6 dogs also developed antibodies against OspF, P39, SLP, and OspC (Table 3). A total of 5 dogs had been administered a bacterin, and 1 dog had been administered the recombinant vaccine. Three of the 6 dogs had OspA antibodies alone, and 1 dog had both OspA and OspC antibodies at the time of I. scapularis infestation and still became infected with B. burgdorferi.
Borrelia burgdorferi antibody test results in 6 dogs that developed infection after being previously vaccinated and then exposed to Ixodes scapularis.*
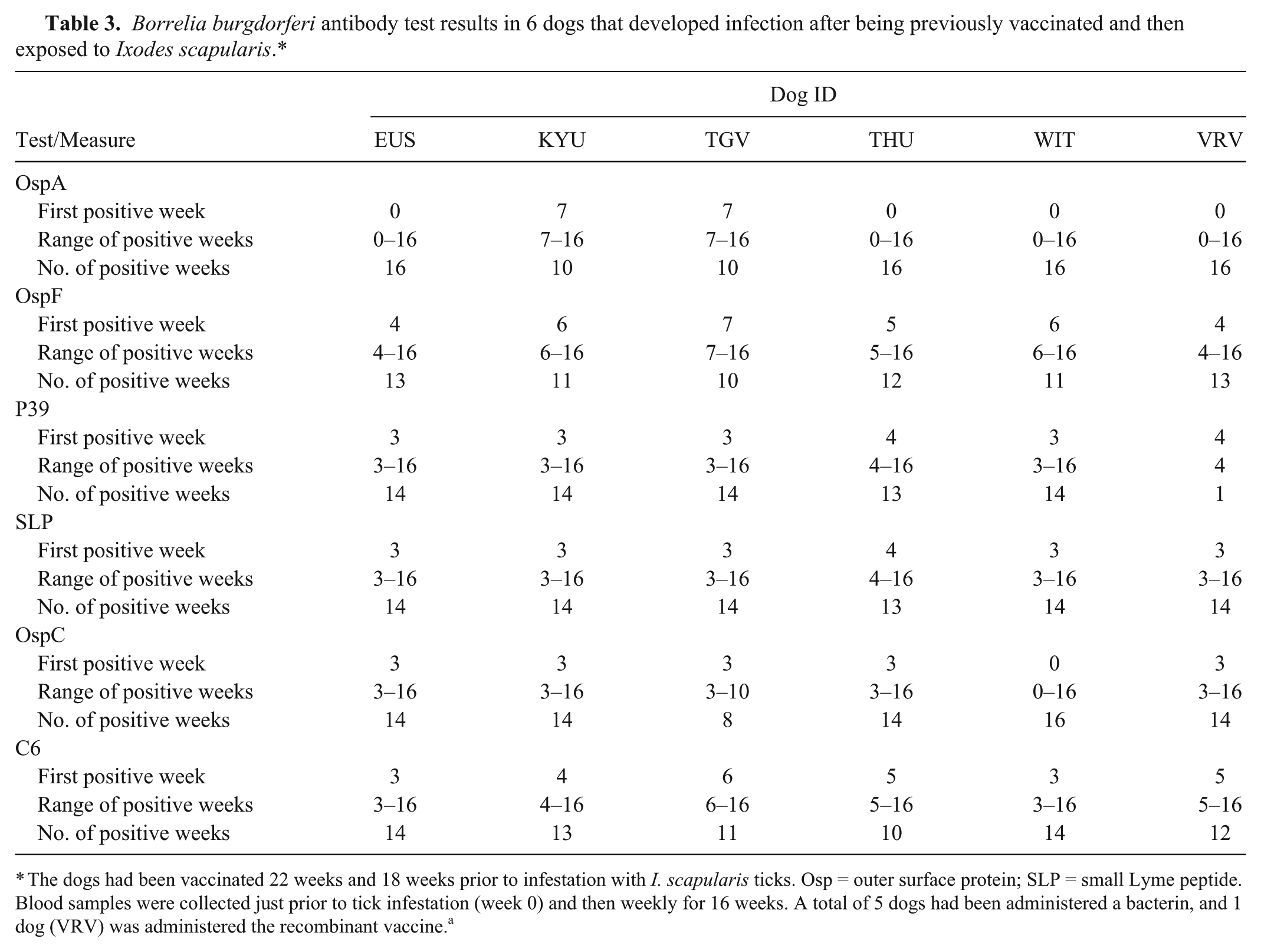
The dogs had been vaccinated 22 weeks and 18 weeks prior to infestation with I. scapularis ticks. Osp = outer surface protein; SLP = small Lyme peptide. Blood samples were collected just prior to tick infestation (week 0) and then weekly for 16 weeks. A total of 5 dogs had been administered a bacterin, and 1 dog (VRV) was administered the recombinant vaccine. a
Experiment 4: Serological responses of B. burgdorferi–infected adult dogs after administration of B. burgdorferi–containing vaccines
The 14 B. burgdorferi–infected dogs from experiment 2 were vaccinated with 1 of 3 commercially available vaccines after blood sample collection at week 12 after I. scapularis infestation. At the time of vaccination, antibodies against OspA (2 dogs), OspF (14 dogs), P39 (12 dogs), SLP (10 dogs), OspC (7 dogs), and C6 (14 dogs) were still detectable in some dogs. After 1 dose of vaccine, all 12 OspA-negative dogs, both P39-negative dogs, 3 of 4 SLP-negative dogs, and 3 of 7 OspC-negative dogs developed detectable antibodies against the respective targets. At the end of the 7-week test period, antibodies against OspA (14 dogs), OspF (13 dogs), P39 (14 dogs), SLP (12 dogs), OspC (7 dogs), and C6 (14 dogs) were still detectable in serum in some dogs.
Discussion
The Accuplex4 system for the detection of B. burgdorferi peptides OspA, OspC, OspF, P39, and SLP had good interassay and intra-assay variation (all ≤12%). All of the vaccines evaluated induced OspA responses with a duration of 20 weeks in the majority of the 18 dogs vaccinated prior to I. scapularis infection in experiment 1. Antibodies against OspF or C6 were not induced in any of the vaccinated dogs and so antibodies against these 2 targets are likely to indicate B. burgdorferi infection as previously discussed.8,18 Antibodies against P39 and SLP were induced by the vaccines studied herein but were of shorter duration than those against OspA. The role that humoral responses to these antigens play in protection against borreliosis in dogs has not been studied. All the dogs vaccinated with the vaccine containing OspC became positive as previously described with this vaccine.1,9,10 As a total of 3 dogs administered the other bacterin d developed detectable OspC antibodies, this indicated the presence of this antigen within the vaccine. Antibodies against OspC induced by vaccination had a median duration of 12 weeks with only 1 dog having a positive test result at 20 weeks, showing that antibody responses against this antigen can wane over time.
When the 18 vaccinated dogs were infested with I. scapularis 22 weeks after the first dose of vaccine was administered, 6 dogs administered 2 different vaccines developed B. burgdorferi infection. These results show that a serologic response to OspA, while confirming vaccination, does not predict resistance to infection in an individual dog and that B. burgdorferi vaccines do not have 100% efficacy. Vaccine failures were noted both for a bacterin and for the recombinant vaccine. While B. burgdorferi vaccines use in the dog can be effective,6,9–11 protection may not be 100%, which emphasizes that vaccines against B. burgdorferi should not be used as the sole prevention strategy; it is imperative to also use acaricides as well. 12 The B. burgdorferi serological responses in the vaccinated dogs that developed infection were similar to those for dogs in experiment 2 that developed infection without being previously vaccinated.
When the 18 previously unvaccinated dogs in experiment 2 were infested with I. scapularis, 14 dogs developed infection as indicated by the presence of B. burgdorferi DNA in skin biopsies (13 dogs) or antibodies against the C6 peptide on at least 3 sample dates (1 dog). The dog that was C6 antibody positive but negative for B. burgdorferi DNA in skin was interpreted as being infected by B. burgdorferi with falsely negative skin biopsy results, which has been previously described. 2 Each of 14 B. burgdorferi–infected dogs also developed antibodies against OspF and P39, and 13 of the 14 dogs developed antibodies against OspC and SLP. Fewer dogs in this experiment developed low levels of short-lasting antibodies against OspA, which was similar to a previous study. 18 While OspA is expressed in the mid gut of infected ticks and becomes downregulated during transmission, the immune system may be briefly exposed to immunogenic OspA, resulting in a transient serological response. 18 In the current study, antibodies against SLP and OspC were detected more quickly in infected dogs than antibodies against C6, OspF, OspA, or P39. Antibodies against OspC have been proposed as markers of early B. burgdorferi exposure in other studies.1,18 While the OspC and OspF responses using the Accuplex4 system and another published multiplex system were similar, the previously reported multiplex system detected OspA antibodies earlier than the Accuplex4 system described herein. 18 However, care must be taken in attempting indirect comparisons of serological response in different studies, as different derivations of peptides can behave differently.1,2,16
When the OspF antibody responses in all 36 dogs were compared with those of the C6 peptide, sensitivity and specificity were shown to both be 100%. Because antibodies against the other markers could be induced by either vaccination or infection, the sensitivities and specificities were <100%. Dogs that are positive for OspF or C6 antibodies are likely to have been previously exposed to B. burgdorferi as previously described.8,18
Antibodies against OspA that were below levels induced by vaccination developed in 6 of the 14 dogs infected with B. burgdorferi in experiment 2 and were only detected in serum for a short period of time. However, after the vaccination of these dogs in experiment 4, all of the B. burgdorferi–infected dogs that were negative for OspA developed detectable antibodies again, and similar results were seen for SLP, P39, and OspC antibodies. These results suggest that vaccines can induce anamnestic responses against these antigens in dogs with previous B. burgdorferi infection. Clinical manifestations of polyarthritis were not noted in these dogs, suggesting that, in our short term study, administering vaccines to previously infected dogs did not exacerbate clinical borreliosis from hypersensitization, which was mentioned as a concern in a B. burgdorferi consensus statement. 12 Urinalyses were not performed on dogs in these experiments and so the effect of B. burgdorferi vaccines on the kidneys of dogs with prior infection could not be assessed.
Overall, the results of our study show that evaluating antibody responses to OspA, OspF, P39, SLP, and OspC simultaneously can provide more information concerning the timing of B. burgdorferi exposure in an individual dog than assessing antibodies against C6 peptide alone, which confirms the findings of others. 18 In addition, it is possible that use of the assay results quantitatively could further refine the stage of exposure and vaccination influence and should be explored in future experiments. However, no B. burgdorferi antibody test results correlate to the presence of clinical illness from B. burgdorferi infection.8,12 In addition, the results described herein should also be interpreted carefully when attempting to compare to cases in the field. In these experimentally infected dogs, only 1 tick exposure was provided. In nature, dogs could be exposed to B. burgdorferi many times, which may induce different antibody responses from those described herein.
Footnotes
Acknowledgements
We thank Michael Ramos, Jennifer Hawley, Melissa Brewer, Amber Caress, Kris Obssuth, Michael Kim, Melissa Arbaugh, and Daniel Fredholm for aid in animal management and sample collection.
a.
Nobivac Lyme, Intervet/Schering-Plough Animal Health, Summit, NJ.
b.
SNAP 4DX, IDEXX Laboratories, Portland, ME.
c.
BioCD, Antech Laboratories, Lake Success, NY.
d.
Lyme Vax, Fort Dodge Animal Health, Fort Dodge, IA.
e.
Recombitek Lyme, Merial Ltd., Duluth, GA.
f.
Dr. Thomas Mathers, Tick Encounter, University of Rhode Island, City, RI.
g.
Frontline, Merial Ltd., Duluth, GA.
h.
Bite Not Collar, Bite Not Products Inc., San Francisco, CA.
i.
OspA, Meridian Laboratories, Charlotte, NC.
j.
OspC, licensed from Mikrogen GmbH, Neuried, Germany.
k.
OspF, licensed from Yale University, New Haven, CT.
l.
P39, licensed from National Institutes of Health, Bethesda, MD.
m.
Biopeptide Co. Inc., East Setauket, NY.
n.
GenScript USA Inc., Piscataway, NJ.
o.
Novagen, Merck KGaA, Darmstadt, Germany.
p.
Genomics Core Facility, Purdue University, West Lafayette, IN.
q.
Ni Sepharose 6 Fast Flow column, Ni-NTA matrix applying 6M-1M linear urea gradient using AKTA Explorer 100 purification system; General Electric, Schenectady, NY.
r.
HiLoad 26/60 Superdex 200 pg column, General Electric, Schenectady, NY.
s.
Bio-Rad Laboratories, Hercules, CA.
t.
SimplyBlue SafeStain, Life Technologies, Grand Island, NY.
u.
Miltenyi Biotech GmbH, Bergisch Gladbach, Germany.
v.
Thermo Scientific Pierce TMB-Blotting 1-step solution, Life Technologies, Grand Island, NY.
Authors’ contributions
S Moroff and MR Lappin contributed to conception and design of the study. S Moroff, I Sokolchik, and MR Lappin drafted the manuscript. All authors contributed to acquisition, analysis, and interpretation of data; critically revised the manuscript; gave final approval; and agree to be accountable for all aspect of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: All the authors except Michael R. Lappin are employees of Antech Diagnostics. However, none of the authors directly benefited financially from completion of this work.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The funding for the study was provided by Antech Diagnostics.