
Abstract
Enzyme-linked immunosorbent assays (ELISAs) based on the SnSAG surface antigens of Sarcocystis neurona provide reliable detection of infection by the parasite. Moreover, accurate serodiagnosis of equine protozoal myeloencephalitis (EPM) is achieved with the SnSAG ELISAs by measuring antibodies in serum and cerebrospinal fluid (CSF) to reveal active infection in the central nervous system. Two independent ELISAs based on recombinant (r)SnSAG2 or a chimeric fusion of SnSAG3 and SnSAG4 (rSnSAG4/3) are currently used together for EPM serodiagnosis to overcome varied antibody responses in different horses. To achieve reliable antibody detection with a single ELISA instead of 2 separate ELISAs, rSnSAG2 was fused with rSnSAG4/3 into a single trivalent protein, designated rSnSAG2/4/3. Paired serum and CSF from 163 horses were tested with all 3 ELISAs. When the consensus antibody titers obtained with the rSnSAG2 and rSnSAG4/3 ELISAs were compared to the single SAG2/4/3 ELISA titers, Spearman rank correlation coefficients of ρ = 0.74 and ρ = 0.90 were obtained for serum and CSF, respectively, indicating strong agreement between the tests. When the rSnSAG2 and rSnSAG4/3 consensus serum-to-CSF titer ratio was compared to the rSnSAG2/4/3 serum-to-CSF titer ratio, the Spearman correlation coefficient was ρ = 0.87, again signifying strong agreement. Importantly, comparing the diagnostic interpretation of the serum-to-CSF titer ratios yielded a Cohen kappa value of 0.77. These findings suggest that the single ELISA based on the trivalent rSnSAG2/4/3 will provide serologic and diagnostic results that are highly comparable to the consensus of the 2 independent ELISAs based on rSnSAG2 and rSnSAG4/3.
The protozoan parasite Sarcocystis neurona is the primary cause of equine protozoal myeloencephalitis (EPM), a debilitating neurologic disease of horses in North, Central, and South America. 1 Horses come into contact with S. neurona by ingesting parasite sporocysts in food or water that has been contaminated with feces from an infected opossum. In most parts of the Americas, horses are commonly exposed to S. neurona and produce antibodies to this parasite. However, only a small proportion of infected horses develop EPM (National Animal Health Monitoring System. Equine protozoal myeloencephalitis (EPM) in the U.S. Fort Collins, CO: USDA, APHIS, VS. Available from: http://purl.umn.edu/32788), which confounds serologic diagnosis of this disease.
The outer membrane of S. neurona merozoites is covered by multiple immunogenic surface antigens designated SnSAGs.2,6 The SnSAGs are homologous to the TgSAG surface antigens originally described in Toxoplasma gondii, 8 and most of these parasite surface proteins are comprised of either a single SAG1-related sequence (SRS)-like domain (e.g., SnSAG2) or 2 SRS-like domains (e.g., SnSAG1, SnSAG3, and SnSAG4). Consistent with their immunogenicity, an earlier study demonstrated the utility of recombinant (r)SnSAGs in enzyme-linked immunosorbent assays (ELISAs) for detecting equine antibodies against S. neurona. 4 In particular, ELISAs based on SnSAG2, SnSAG3, and SnSAG4 exhibited superior sensitivity and specificity when testing samples from seropositive and seronegative horses.
Importantly, it has been observed that some equine samples show significant variation in reactivity to the different rSnSAGs (Morrow JK, et al., unpublished results), 4 which reduces the diagnostic accuracy that can be achieved using a single SnSAG in a serologic assay. A 2011 study described a chimeric fusion protein containing the amino-terminal domain of SnSAG4 fused to the carboxyl-terminal domain of SnSAG3 (rSnSAG4/3) that was constructed to serve as a polyvalent antigen in an ELISA. 11 The new rSnSAG4/3 ELISA showed high sensitivity and specificity for detecting antibodies against S. neurona in equine serum. 11 The rSnSAG2 and rSnSAG4/3 ELISAs are now being used together to assess the EPM status of horses (i.e., Equine Diagnostic Solutions LLC, Lexington, Kentucky), with both assays run in parallel to enhance diagnostic sensitivity.
In an effort to reduce the time, materials, and cost associated with running 2 separate ELISAs, a triple-domain chimeric fusion protein containing epitopes from SnSAG2, SnSAG3, and SnSAG4 was prepared and evaluated in the present study. The new triple-domain expression plasmid rSnSAG2/4D1/3D2 was constructed with cloning strategies similar to those used to make the rSnSAG4/3 chimera (see Fig. 1, Supplementary Table 1; Supplementary Tables, available at http://vdi.sagepub.com/content/by/supplemental-data). 11 Briefly, the complementary DNA clone of SnSAG2 6 was used as polymerase chain reaction (PCR) template with primers designed to amplify a DNA fragment that would serve as the amino-terminal portion (reverse primer with 3′ overhangs complimentary to 5′-end of SnSAG4D1) of a triple-domain recombinant protein. Similarly, the previously constructed plasmid pET4D1/3D2 11 for expression of rSnSAG4/3 was used as PCR template with primers designed to amplify a DNA fragment that would serve as the carboxyl-terminal portion of the triple-domain protein. The SnSAG2 and SnSAG4/3 DNA fragments were purified a before being used in a fusion PCR 10 to splice the SnSAG2 and SnSAG4/3 PCR products together into the SnSAG2/4D1/3D2 DNA fragment. The unified PCR product was digested with the appropriate enzymes and ligated into the pET22b expression vector, b which incorporates a carboxyl-terminal His-tag into the recombinant protein. The new expression plasmid was transformed into Escherichia coli DH5α, and recombinant clones were screened to identify plasmids containing the appropriately sized inserts. Selected plasmids were transformed into E. coli BL21 expression cells, and the new rSnSAG chimera was expressed and purified by nickel-column chromatography. The purified rSnSAG2/4/3 protein was incorporated into an indirect ELISA and run essentially as described previously.4,11
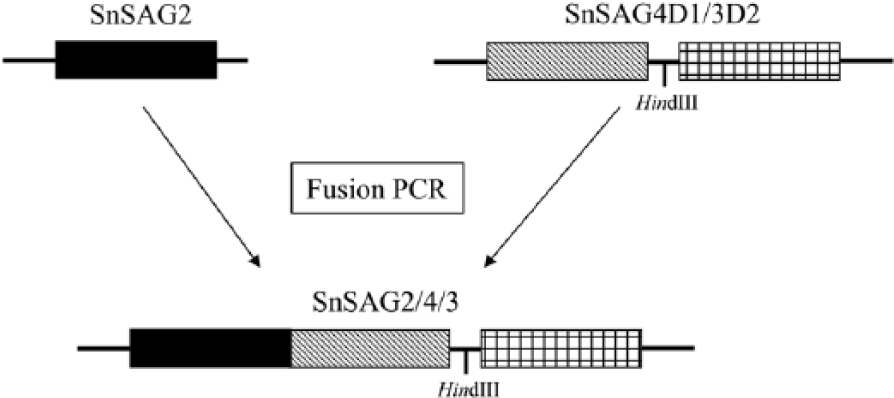
Simple graphic depiction of the rSnSAG2/4/3 trivalent recombinant antigen. The single domain of SnSAG2 and the 2 domains of the previously constructed SnSAG4/3 chimera were amplified independently, and then combined into a single DNA fragment by fusion polymerase chain reaction (PCR).
To get a preliminary assessment of the new trivalent antigen ELISA, a collection of 34 equine serum samples were tested using the rSnSAG2, rSnSAG4/3, and rSnSAG2/4/3 ELISAs. ELISA results were obtained by different operators in 2 independent laboratory facilities; the rSnSAG2 and rSnSAG4/3 ELISAs were performed at Equine Diagnostic Solutions LLC, and the rSnSAG2/4/3 ELISA was performed at the University of Kentucky Gluck Equine Research Center (Lexington, Kentucky). Several of the serum samples in the collection were selected specifically because the rSnSAG2 and rSnSAG4/3 ELISAs yielded discrepant endpoint titers for these samples. The consensus (i.e., higher) endpoint antibody titer from the rSnSAG2 and rSnSAG4/3 ELISAs (designated rSnSAG2,4/3) was compared to the endpoint titer obtained with the rSnSAG2/4/3 ELISA (Table 1) and analyzed by Spearman rank correlation coefficient. This analysis yielded a Spearman coefficient of ρ = 0.72, indicating very good agreement between the rSnSAG2,4/3 consensus result and the rSnSAG2/4/3 ELISA result. Identical endpoint titers were observed for 10 of the 34 serum samples, while a difference of only a single dilution factor was observed for another 18 of the samples (Table 1). A large difference was observed for 1 sample (no. 28), which had a rSnSAG2,4/3 reciprocal consensus titer of 4,000 but was negative by rSnSAG2/4/3. While there may be a biological basis for this, it is conceivable that the observed difference was due to a technical error (e.g., a mislabeled sample) because the assays were run in independent laboratory facilities using split samples. Interestingly, the rSnSAG2/4/3 ELISA titer was often intermediate when the rSnSAG2 and rSnSAG4/3 independent titers were discrepant (e.g., sample 1 in Table 1). Collectively, the results obtained with this sample set of 34 sera demonstrated that the new rSnSAG2/4/3 trivalent antigen will provide serologic results in a single ELISA that are very comparable to the consensus results obtained with the 2 independent assays based on rSnSAG2 and rSnSAG4/3.
Reciprocal endpoint serum titers obtained from 34 equine serum samples using the rSnSAG2, rSnSAG4/3, and rSnSAG2/4/3 enzyme-linked immunosorbent assays.
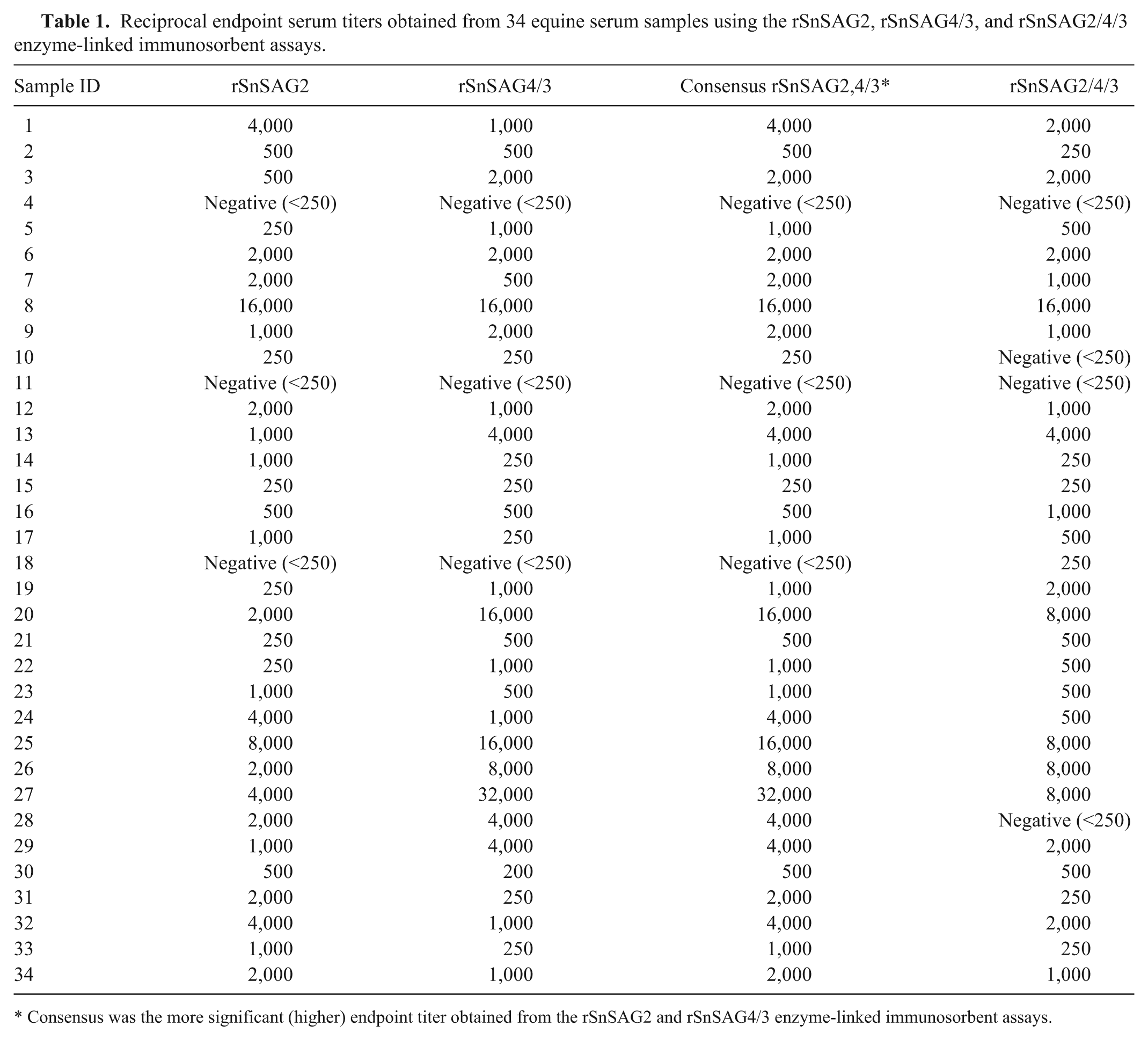
Consensus was the more significant (higher) endpoint titer obtained from the rSnSAG2 and rSnSAG4/3 enzyme-linked immunosorbent assays.
As the initial evaluation of the rSnSAG2/4/3 ELISA gave promising results, a more extensive comparison of the ELISAs was made using a collection of paired serum and cerebrospinal fluid (CSF) samples from 163 horses. When the rSnSAG2,4/3 consensus endpoint titers for the serum samples were compared with the rSnSAG2/4/3 serum endpoint titers, the titers were concordant or within 1 dilution difference for ~64% of the samples (Fig. 2A; Supplementary Table 2). The Spearman coefficient was ρ = 0.74, indicating good agreement between the test results. Comparison of the endpoint titers obtained for the 163 CSF samples yielded an even higher Spearman coefficient of ρ = 0.90, with ~75% of the samples showing concordant titers or 1 dilution difference (Fig. 2B; Supplementary Table 2). These results further confirmed that the single ELISA based on rSnSAG2/4/3 will produce serologic data that agrees strongly with the consensus data obtained with the 2 independent ELISAs.
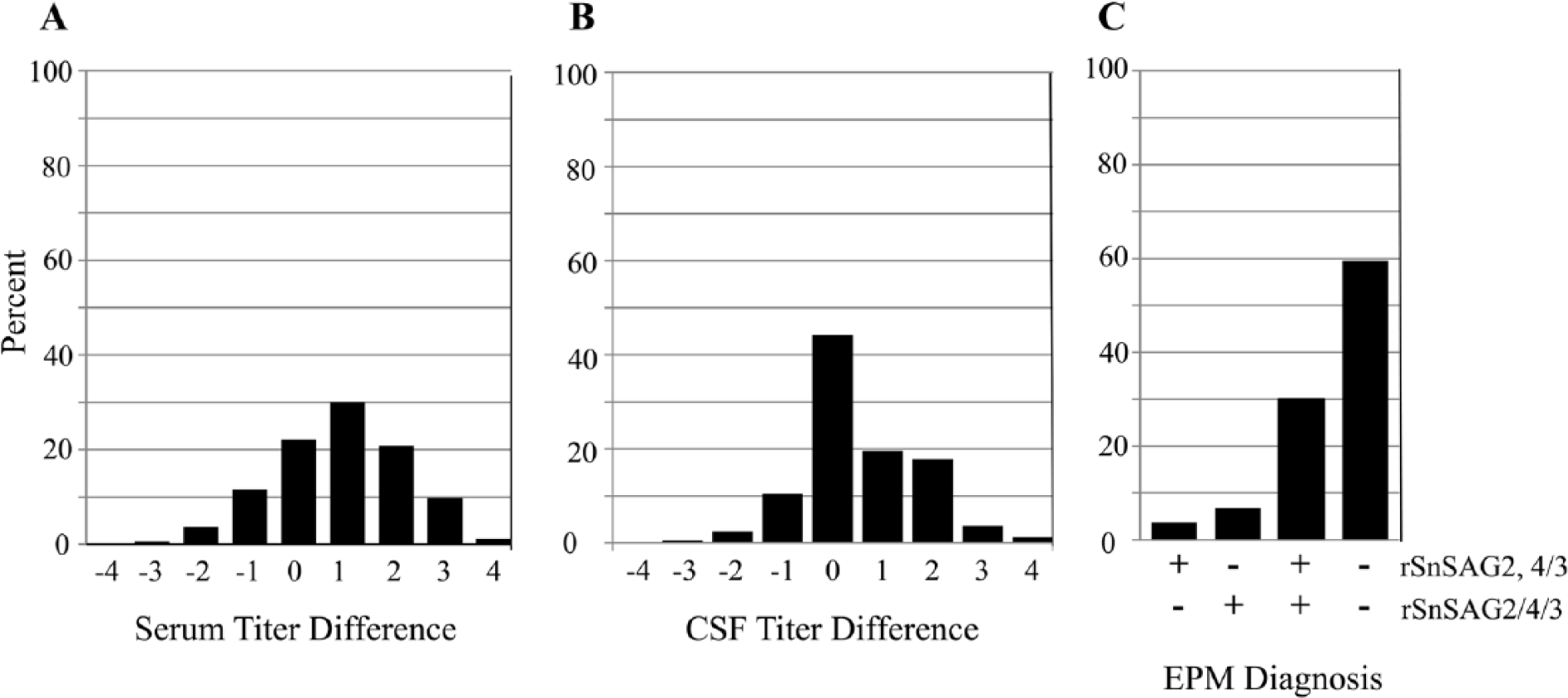
Agreement between the rSnSAG2,4/3 consensus result and the rSnSAG2/4/3 result for serum and cerebrospinal fluid (CSF) samples from 163 horses.
Horses in many regions of the Americas are commonly exposed to S. neurona, so simple detection of serum antibodies against the parasite has minimal diagnostic value. Detection of antibodies in CSF is more informative, but diagnosis of EPM based on CSF results is still confounded by normal passive transfer of antibodies across the blood-brain barrier. Therefore, diagnosis of EPM using serologic assays is best accomplished by measuring parasite-specific antibodies in both serum and CSF to assess intrathecal antibody production, which indicates infection in the central nervous system (CNS).3,7,9 The current diagnostic protocol involves using the rSnSAG2,4/3 ELISAs to determine a consensus serum-to-CSF titer ratio, with ratios ≤100 suggesting that antibodies against S. neurona are being produced in the CNS.7,9 To assess whether the single rSnSAG2/4/3 ELISA would yield serum-to-CSF ratios similar to the rSnSAG2,4/3 ELISAs, the ratios for the 163 paired serum and CSF samples were calculated and compared (see Supplementary Table 2). This gave a Spearman rank correlation coefficient of ρ = 0.87, which implies very strong agreement between the rSnSAG2,4/3 consensus and the rSnSAG2/4/3 single test. The diagnostic interpretation of the serum-to-CSF titer ratios (i.e., ≤100 implies EPM; >100 implies not EPM) obtained with the SnSAG2,4/3 test and the SnSAG2/4/3 test was also compared (Fig. 2C). Of the 163 paired samples, the rSnSAG2,4/3 titer ratio suggested that 56 cases were EPM horses while the rSnSAG2/4/3 titer ratio suggested that 61 cases were EPM. Overall, the 2 tests yielded the same diagnosis for ~90% of the cases (146/163) for a Cohen kappa coefficient of 0.77, thus signifying substantial agreement. Because the kappa statistic is considered an overly conservative measure of agreement, it was anticipated that the kappa value would be lower than the nonparametric analysis (i.e., Spearman coefficient).
In conclusion, the immunogenic SnSAG surface antigens have proven to be very useful for detection of antibodies against S. neurona and the diagnosis of EPM.3,4,7,9 However, antigenic difference exists between different isolates of S. neurona, 5 and it is apparent that horses vary in the array of parasite antigens against which they produce antibodies (Morrow JK, et al., unpublished results). 4 Consequently, serologic testing to aid EPM diagnosis is best achieved by incorporating multiple S. neurona antigens into the serologic assay. For commercial testing, the rSnSAG2 ELISA and the rSnSAG4/3 ELISA are presently run separately but the results from the 2 assays are considered collectively. This approach has been shown to be very effective for identifying horses suffering from EPM caused by S. neurona.7,9 The findings from the present study indicate that the single ELISA based on the trivalent fusion proteins rSnSAG2/4/3 will provide serologic and diagnostic results that agree well with the collective results obtained from the 2 ELISAs based on rSnSAG2 and rSnSAG4/3. Therefore, it is reasonable to consider replacing the 2-ELISA approach with the single rSnSAG2/4/3 ELISA as this will reduce the time, labor, and costs associated with serologic testing for the diagnosis of EPM.
Footnotes
Acknowledgements
We thank Hernani Kerber for assisting with ELISA analysis of samples.
Authors’ note
The article is published as Kentucky Agricultural Experiment Station article no. 15-14-002.
a.
Wizard SV gel, PCR Clean-up system; Promega Corp., Madison, WI.
b.
Novagen, EMD Millipore, Billerica, MA.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: The University of Kentucky Research Foundation has obtained patent protection for the technologies described in this article, and the assays have been licensed for commercial use by Equine Diagnostic Solutions LLC.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research was made possible by funds from the Amerman Family Equine Research Endowment. Izabela de Assis Rocha was supported by a scholarship from CAPES, Brazil’s National Council for the Improvement of Higher Education.