
Abstract
Gastrointestinal stromal tumors (GISTs) and leiomyosarcomas are histologically similar primary neoplasms commonly occurring in the gastrointestinal tract of dogs and humans. Immunohistochemical staining (IHC) is needed to differentiate between these 2 entities and positive reactivity for KIT (cluster of differentiation [CD]117) is regarded as the gold standard for diagnosis of canine GIST. Studies estimate 5–10% of human GISTs stain negative or only weakly positive for KIT and have identified DOG1 (discovered on gastrointestinal stromal tumors protein 1) as a highly sensitive and specific marker for human GISTs. The purpose of this study was to evaluate immunoreactivity of a commercially available DOG1 antibody for use in diagnosis of canine GISTs. Fifty-five primary mesenchymal gastrointestinal tumors with histologic features consistent with GIST or leiomyosarcoma were evaluated via IHC for KIT, DOG1, and desmin. A subset of tumors was additionally evaluated for reactivity for smooth muscle actin (SMA). Thirty-three tumors (60%) were diagnosed as GIST based on positive immunoreactivity for KIT or DOG1 regardless of reactivity for desmin or SMA. Most GISTs (32/33, 97.0%) had similar staining for both KIT and DOG1. DOG1 expression was identified in 2 tumors (1 study tumor and 1 additional tumor) negative for KIT and desmin that had histologic features consistent with KIT-negative, platelet-derived growth factor receptor-alpha (PDGFRA)-mutant human GISTs. Our results suggest that DOG1 has improved specificity and sensitivity to that of KIT for differentiating between canine GISTs and leiomyosarcomas. Inclusion of both DOG1 and KIT IHC in diagnostic panels will improve the accuracy of canine GIST diagnosis.
Keywords
Introduction
Gastrointestinal stromal tumors (GISTs) and leiomyosarcomas are histologically similar neoplasms commonly occurring in the gastrointestinal tract of dogs and humans. Immunohistochemical stains are needed to differentiate between these 2 entities. Accurate diagnosis is important in part because GISTs and leiomyosarcomas are thought to arise from different cells and may have different biological behavior.22,43,53 More studies are needed to better understand the malignant tendencies of both tumors and to obtain accurate diagnoses. GISTs commonly have gain-of-function mutations in the c-KIT gene or less commonly the platelet-derived growth factor receptor-alpha (PDGFRA) gene. These tumors may respond to targeted molecular therapy with tyrosine kinase inhibitors.10,24,49
Canine GISTs are currently differentiated from other gastrointestinal sarcomas by positive immunoreactivity for KIT (cluster of differentiation [CD]117).4,16,35 This distinction followed findings that human GISTs arise from cells sharing features of interstitial cells of Cajal (ICC) including expression of KIT protein.27,41,44 In fact, some GISTs may be driven by activating mutations in the c-KIT gene, making them sensitive to treatment with receptor tyrosine kinase inhibitors that target KIT. 27 KIT-positive gastrointestinal sarcomas have also been reported in horses, a chimpanzee, a cat, and a F344 rat.9,17,38,45 In addition to KIT, GISTs demonstrate variable reactivity for CD34 and α-smooth muscle actin (SMA), and are negative for desmin and S100. Leiomyosarcomas more commonly stain positive for SMA and desmin.4,16,18,22,43 In humans, ~5–10% of GISTs are KIT negative or only weakly positive.8,14,33,36,52 These KIT-negative tumors are clinicopathologically indistinct from KIT-positive GISTs and have characteristic loss of chromosomes 1p, 14, and/or 22 via karyotype analysis.8,12,13,25,36 KIT-negative GISTs may still have activating mutations in c-KIT, or often have activating PDGFRA mutations and thus may remain responsive to treatment with kinase inhibitors. Markers such as PDGFRA, protein kinase Θ (PKΘ), and DOG1 (discovered on gastrointestinal stromal tumors protein 1) have been explored in human tumors in hopes of identifying GISTs in this subgroup.33,36 Several human studies have concluded that DOG1 should be included in diagnostic panels designed to identify GISTs.14,33,52
DOG1 (syn. ANO1, FLJ10261, and TMEM16A) is a plasma membrane protein involved in calcium-dependent chloride transport5,54 specifically expressed by GISTs and discovered during analysis of tissue microarray results comparing various sarcomas.39,52 In the intestine, DOG1 is specifically expressed in ICCs in humans and mice, and there is evidence DOG1 promotes proliferation of ICCs and is required for normal gut contractility.19,47,48 DOG1 is typically coexpressed with KIT in ICCs, though populations of KIT-negative, DOG1-positive ICC cells have been identified in the mouse colon.19,30,50 A number of studies in human GISTs have shown DOG1 to be a highly sensitive and specific marker for these tumors using in situ hybridization (ISH) and/or immunohistochemical staining (IHC).14,26,33,52 DOG1 stains a majority of c-KIT and PDGFRA mutant as well as wild type GISTs while only occasionally staining other tumor types.14,26
Specifically, KIT and DOG1 expression were compared in 935 non-GIST sarcomas with similar results: KIT was expressed in 3 of 331 (0.9%) leiomyosarcomas, 1 of 10 (10%) desmoplastic melanomas, and 1 of 87 (1.1%) undifferentiated sarcomas; DOG1 was expressed in 1 of 326 (0.3%) leiomyosarcomas, 1 of 10 (10%) desmoplastic melanomas, and 1 of 39 (2.5%) synovial sarcomas. 14 In addition, utilizing a tissue microarray of 432 non-GIST “nonsarcomas,” DOG1 expression was identified in 4 carcinomas and a desmoplastic melanoma compared with KIT expression in 87 tumors including a variety of carcinomas, melanomas (nondesmoplastic and desmoplastic), seminomas, brain tumors, and 1 diffuse large B-cell lymphoma. 14 In this report, staining intensity was required to be moderate to strong with no cutoff for percent of cells staining for a positive call. Thus, DOG1 demonstrated equal-to-superior specificity compared with KIT in non-GIST tumors. Further, in a study examining only DOG1 staining patterns, expression of DOG1 in 3 of 117 non-GIST sarcomas (1 each desmoplastic melanoma, malignant peripheral nerve sheath tumor, and schwannoma) and 11 of 50 carcinomas (3 colorectal, 4 endometrioid, and 4 acinic cell adenocarcinomas) was typically focal (<5–50% cells staining), of weak to moderate intensity, and, in the case of carcinomas, showed a distinct luminal pattern easily distinguished from the diffuse, moderate to strong staining typically seen in GISTs. 26 Expression in up to 50% of cells was reported in the desmoplastic melanoma, malignant peripheral nerve sheath tumor, and acinic cell adenocarcinomas. KIT IHC was not performed in these tumors. Given the observed sensitivity and specificity of staining for DOG1 in human GISTs, our laboratory recently evaluated a commercially available rabbit monoclonal anti-human DOG1 antibody for use in canine tissues.
The goals of this study were to document the immunoreactivity of a monoclonal antibody (mAb) to DOG1 and to compare the results of IHC for DOG1 to that of KIT for the differential diagnosis of canine GISTs and leiomyosarcomas. Fifty-five primary mesenchymal gastrointestinal tumors with histologic features consistent with GIST or leiomyosarcoma (Table 1) were evaluated via IHC for KIT, DOG1, and desmin. Our results indicate that DOG1 is a sensitive and specific marker for canine GISTs that may be an alternative to KIT or used in addition to KIT as part of a gastrointestinal diagnostic panel similar to current recommendations in the human literature.
Summary of signalment and diagnoses for 55 dogs with primary gastrointestinal mesenchymal tumors.*
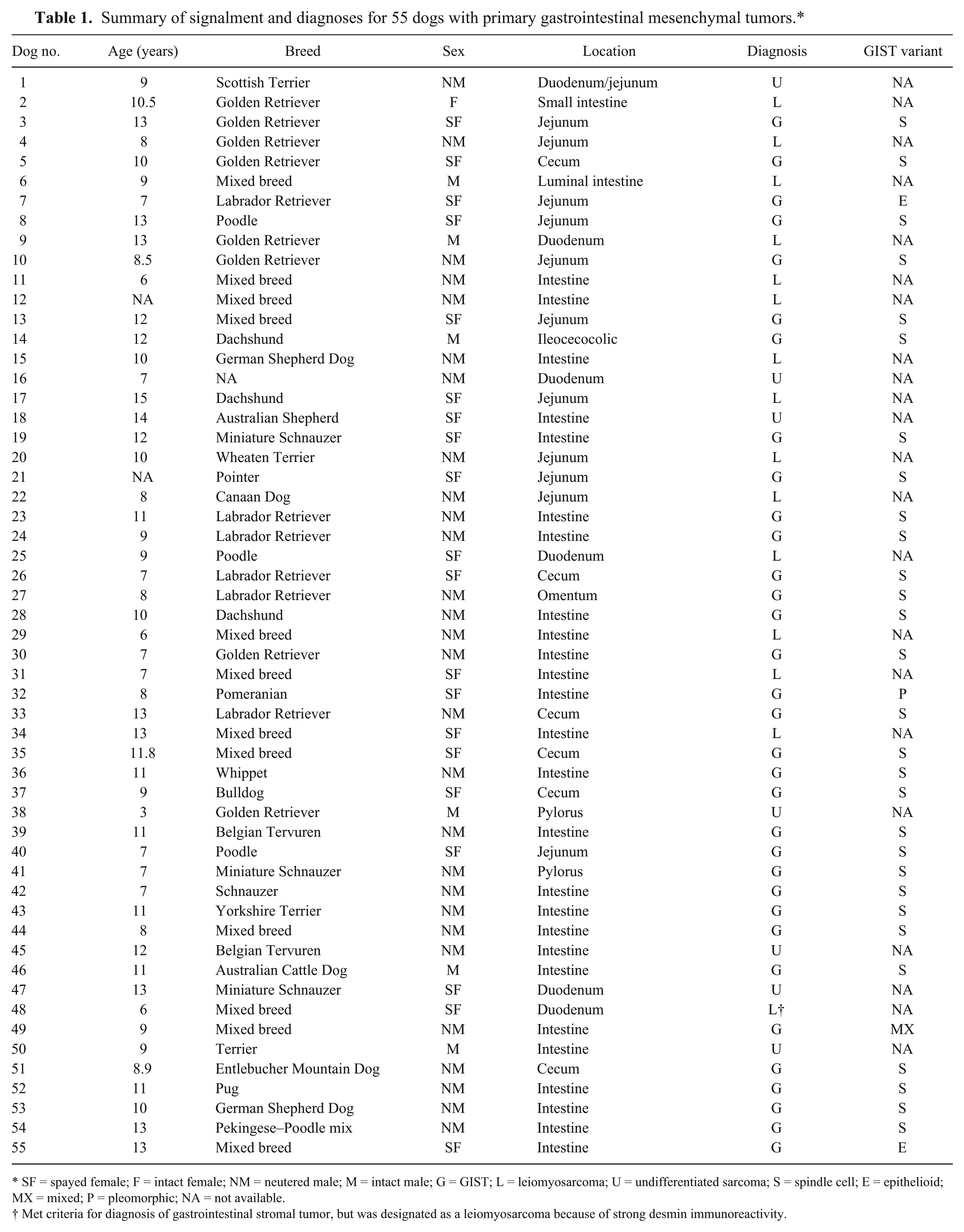
SF = spayed female; F = intact female; NM = neutered male; M = intact male; G = GIST; L = leiomyosarcoma; U = undifferentiated sarcoma; S = spindle cell; E = epithelioid; MX = mixed; P = pleomorphic; NA = not available.
Met criteria for diagnosis of gastrointestinal stromal tumor, but was designated as a leiomyosarcoma because of strong desmin immunoreactivity.
Materials and methods
Tumor selection
Fifty-five canine tumors submitted to the Colorado State University Veterinary Diagnostic Laboratory (CSU-VDL; Fort Collins, Colorado) for routine biopsy between 2005 and 2012 were utilized in this study (Table 1). All tissues had been fixed in 10% neutral buffered formalin for at least 24 hr and then embedded in paraffin following standard processing protocols. Thirty-two tumors were selected from a time period before the CSU-VDL offered IHC for KIT (2005 and 2008) and were therefore diagnosed as leiomyosarcomas based on histologic characteristics alone. The other 23 tumors were diagnosed as GIST or leiomyosarcoma and were submitted between 2011 and 2012. All tumors were primary mesenchymal tumors located within the gastrointestinal tract with histologic features consistent with GIST or leiomyosarcoma as previously described in both human and veterinary literature.4,15,32,37 Tumors with benign characteristics consistent with leiomyoma, or with histologic characteristics of other primary tumors (adenocarcinoma, lymphoma), were not included in this study.
Immunohistochemical staining
For each of the 55 tumors, IHC for DOG1, KIT, and desmin was performed with an indirect biotin-free detection kit designed for use with an automated immunostainer.a,b Primary antibodies used were a rabbit mAb for DOG1 c (1:600 dilution), a mouse mAb for desmin d (ready-to-use), and a rabbit polyclonal antibody for KIT e (1:150). Selected samples were also stained for anti-SMA utilizing a mouse mAb f (ready-to-use). Positive control tissues consisted of normal canine intestine (desmin, SMA) and normal intestinal ICC (KIT and DOG1). Canine mast cell tumors and KIT-positive GISTs identified as staining strongly for KIT and DOG1 were used for additional positive technical controls. Additionally, the majority of tumors examined contained varying amounts of normal intestinal tissue, including ICCs, providing internal positive controls. For negative controls, primary antibodies were replaced with nonspecifically targeted mAbs of the same isotype (immunoglobulin G) and same species as the primary antibodies (mouse or rabbit negative isotype control antibodies).
For all markers, average percentage of tumor cells staining, cellular staining pattern, and stain intensity were recorded by a single pathologist (DL Dailey). Cutoffs for IHC positivity were established based on the range of criteria recorded for each marker as well as previously published guidelines.4,14,22,32,52 Samples were considered KIT or DOG1 positive if ≥50% of tumor cells showed distinct cytoplasmic and/or membranous staining of any intensity. Nuclear KIT staining (reported as rarely occurring in human leiomyosarcoma 52 ) or faint, indistinct staining consistent with background was noted, but considered negative. For SMA and desmin, samples were considered positive if ≥25% of tumor cells stained positive with distinct cytoplasmic staining. This was consistent with reports in the veterinary literature where >70% of tumor cell staining was required for designation as KIT positive, while SMA and desmin reactivity was considered positive over a more variable range. 22 When immunostaining was equivocal, 3 pathologists (Dailey, Ehrhart, and Powers) reviewed the case and reached a consensus.
Western blot
Western blot analysis was performed using tissue lysates prepared from 2 additional canine tumors (not included in the 55 tumor study population) from the Flint Animal Cancer Center (Fort Collins, Colorado) tissue archive, which included both fresh frozen and corresponding formalin-fixed, paraffin-embedded (FFPE) samples. One was from a GIST liver metastasis verified as KIT and DOG1 positive via IHC and the other was from an undifferentiated intestinal sarcoma negative via IHC for KIT, DOG1, and desmin. Lysates were prepared in triethanolamine (TEA) lysis buffer (55 mM TEA [pH 7.5], 111 mM NaCl, 2.2 mM ethylenediamine tetra-acetic acid [EDTA], and 0.44% sodium dodecyl sulfate [SDS]) with 1× protease inhibitor. g Samples were separated using SDS–polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. The membrane was blocked with a commercial blocking agent h for 2 hr at room temperature and incubated with the same rabbit mAb used for IHC (1:1,000 in blocking reagent) at 4°C overnight. After 0.1% Tris buffered saline–Tween 20 (TBST) rinses and a 15-min block in hydrogen peroxide, the membrane was incubated with a secondary horseradish peroxidase–conjugated goat anti-rabbit antibody i (1:20,000 in TBST) for 2 hr at room temperature. A chemiluminescent horseradish peroxidase substrate j was used for detection of antibody binding.
Molecular evaluation for c-KIT and PDGFRA mutations
Approximately 2 mm3 of neoplastic tissue from 8 GISTs (6 KIT and DOG1 positive; 2 KIT negative and DOG1 positive) was excised from each FFPE block targeting sections corresponding to KIT and DOG1 (or in the case of the KIT-negative tumors, DOG1 only) positive areas. DNA was isolated as previously described.2,55 Briefly, FFPE tissue was heated for 10 min at 95°C in 400 μL of digestion buffer (50 mM Tris [pH 8.5], 1 mM EDTA, 0.5% Tween), vortexed, microwaved for 1 min (pausing to vortex every 30 sec), allowed to cool, then incubated overnight at 42°C after addition of 5 μL of proteinase K (15 mg/mL). Digested samples were heated to 95°C for 10 min, and then centrifuged for 10 min at 12,000 rpm. One-hundred fifty microliters was removed from this tube, avoiding remaining paraffin film, moved to a new tube, and carried forward into PCR.
Exons 12, 14, and 18 of PDGFRA and exons 8, 9, 11, 13, and 17 of c-KIT were amplified from tissue sections via PCR, using conditions and primers (for c-KIT) optimized in earlier studies.22,28,51 The expected amplicon sizes for the c-KIT primers ranged from 149 to 328 base pairs. Primers for PDGFRA exons were newly designed to amplify regions corresponding to the common mutations identified in human GIST 7 and are listed in Table 2. PCR was set up in 50-μL total reaction volume consisting of 50 or 100 ng of DNA template, 5 pmol of each primer, 1.25 U of Taq polymerase, k and final concentrations of 0.2 μM deoxynucleoside triphosphate and 2 mM MgCl2. Cycling conditions were 94°C for 4 min; 40 cycles of 95°C for 30 sec, annealing temperatures averaging 58°C for 30 sec, and 72°C for 20 sec; followed by a final elongation step at 72°C for 5 min.
Primers for PDGFRA polymerase chain reaction.*
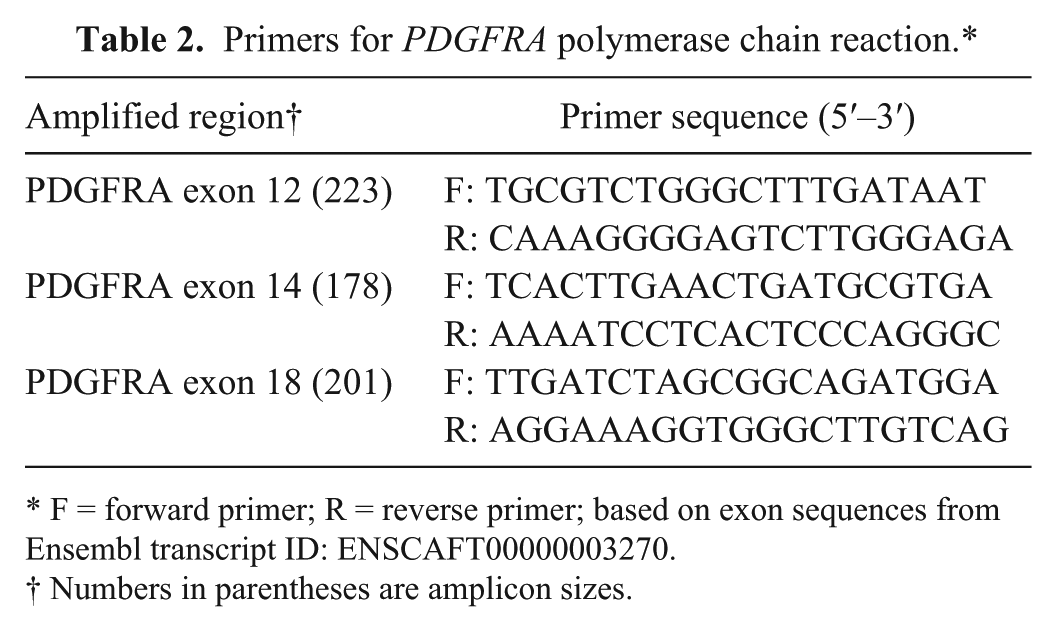
F = forward primer; R = reverse primer; based on exon sequences from Ensembl transcript ID: ENSCAFT00000003270.
Numbers in parentheses are amplicon sizes.
Polymerase chain reaction products were subjected to electrophoresis on 1% agarose gels, l visualized under ultraviolet light, and extracted from the gel. m DNA (20–30 ng) from each gel-isolated PCR product was submitted in 20-μL reactions with 10 pmol of the appropriate primer to the Colorado State University’s Proteomics and Metabolomics Facility for automated sequencing. n Sequences generated from both the forward and reverse primers were examined for each exon of interest for each sample. DNA previously extracted from cultured canine osteosarcoma cells and determined to be wild type for the c-KIT and PDGFRA exons of interest was amplified under similar conditions and used as a control.
Results
Fifty-five primary gastrointestinal sarcomas from the CSU-VDL archives were reviewed and selected for additional IHC and analysis. The average age of all dogs was 9.9 years (range: 3–15.5 years). There were 32 (58.1%) males and 23 (41.8%) females. Of the 54 dogs for which breed was specified, the top 5 breeds represented were 12 (22.2%) mixed breed, 8 (14.8%) Golden Retrievers, 6 (11.1%) Labrador Retrievers, and 3 (5.6%) each Dachshunds and Poodles. The remaining 22 (40.7%) dogs included a variety of breeds of which none was represented more than twice. Lesions were located throughout the gastrointestinal tract most commonly in the small intestine.
Tumors ranged from well-to-poorly demarcated, unencapsulated, infiltrative mass lesions comprised of predominantly spindle-shaped cells supported by a fibrovascular stroma. Individual cells had indistinct cell borders and moderate fibrillar to occasionally vacuolated brightly eosinophilic cytoplasm. Cells were typically arranged in streaming bundles frequently intersecting at variable angles, but also occasionally arranged as palisading cells and/or whorls (Fig. 1A). Most commonly, cell density was high, with cells arranged in close proximity to one another, but several tumors had areas of loosely arranged cells separated by small to moderate amounts of mucin. Nuclei were oval to oblong, with stippled chromatin and variably distinct nucleoli. Mitotic figures averaged 2 per high-powered field (HPF), ranging from less than 1/HPF to greater than 5/HPF. Degree of nuclear pleomorphism, hemorrhage, and necrosis varied between and within tumors. Rarely, cells were epithelioid with a round shape, a larger oval-to-round nucleus with prominent nucleoli, vacuolated cytoplasm, and arranged in sheets (Fig. 1E).
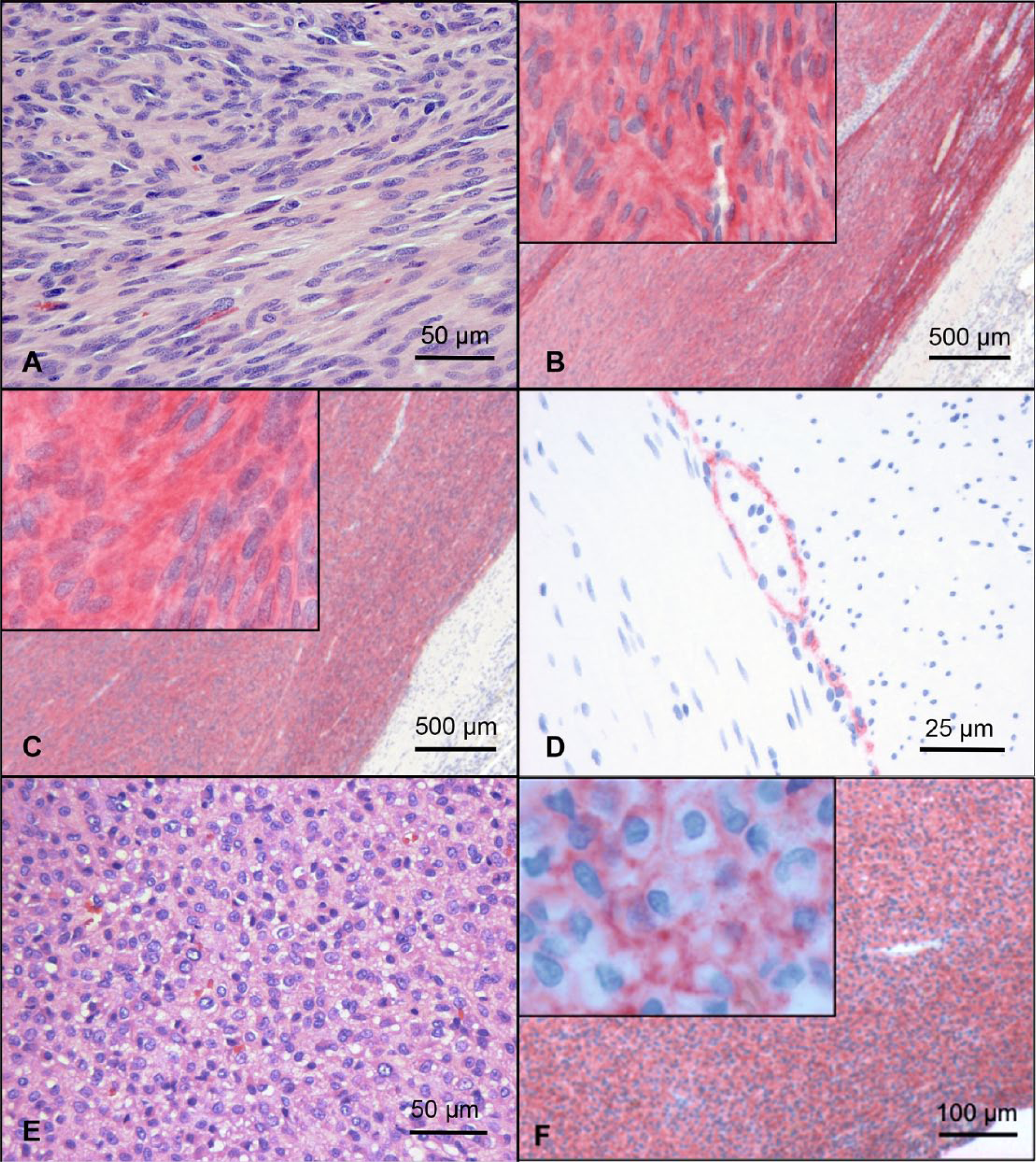
Gastrointestinal stromal tumor (GIST), dog, small intestine.
Gastrointestinal stromal tumor
A diagnosis of GIST was assigned for any tumor that was either KIT positive or DOG1 positive regardless of immunoreactivity for other markers. Thirty-three (60.0%) tumors were diagnosed as GIST based on these criteria. GISTs were diagnosed in 15 (45.5%) female and 18 (54.5%) male dogs with an average age of 10.2 years (range: 7–15.5 years). GISTs were most commonly located in the small intestine (n = 21, 63.6%) including 2 in the duodenum and 7 in the jejunum. For the remaining 13, a more specific location was not specified, but based on microscopic features such as lack of Bruner glands or Peyer patches and moderate to high numbers of goblet cells, location was considered consistent with aboral duodenum or jejunum. Nine (27.3%) were located at the ileocecocolic junction (2) or cecum (7), 2 (6.1%) in the colon, and 1 (3.0%) in the omentum. Consistent with other reports, none were located in the stomach.4,18,22 The majority of tumors diagnosed as GIST (29, 87.9%) were made up of spindle-shaped cells. The remaining tumors (4, 12.1%) were of epithelioid (2; dogs 7 and 55) or mixed morphology (1; dog 49) with 1 tumor (dog 32) containing areas of moderately pleomorphic cells.
Thirty-two (97.0%) GISTs stained positive for both KIT and DOG1. None of these tumors stained positive for desmin and, in all cases, the staining was remarkably similar in both intensity and percentage of cells staining (Fig. 1B, 1C). In all but 3 of these cases, 75% or more of tumor cells stained distinctly positive. Staining for both KIT and DOG1 was predominantly strong and diffusely cytoplasmic, with membranous staining less commonly evident (Fig. 1B and 1C, insets). In the normal intestine, both KIT and DOG1 antibodies stained a fine network of cells located between the muscular layers and frequently associated with myenteric plexi, consistent with ICCs (Fig. 1D). Western blot analysis using the same DOG1 antibody used for IHC showed binding of the DOG1 mAb to a 110–120 kDa protein confirming the specificity of the antibody in canine tissues (Fig. 2). Unlike KIT, DOG1 did not react with mast cells in the lamina propria, which is consistent with previous reports. 14 One KIT-negative case was positive for DOG1. Greater than or equal to 90% of tumor cells showed immunoreactivity for DOG1 with a predominant membranous pattern (with less cytoplasmic intensity), and intensity ranged from light to moderate (Fig. 1F). The predominant morphology of cells in this tumor was epithelioid (Fig. 1E). A second KIT-negative, DOG1-positive GIST submitted to our diagnostic laboratory after analysis of the original study population showed similar morphology and IHC characteristics.
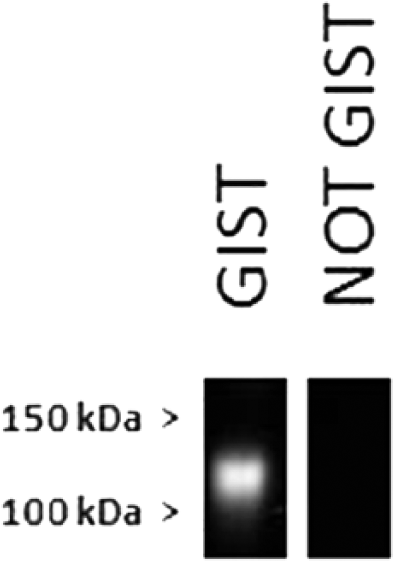
Western blot for DOG1 in 2 canine tumors pulled from archived tissue (not included in the 55 tumors of this study) showed a band in the range of 110–120 kDa. Lane 1 is protein from a liver metastasis from a canine gastrointestinal stromal tumor (GIST), which was KIT and DOG1 positive by immunohistochemical staining (IHC). Lane 2 is protein from a canine gastrointestinal sarcoma negative for KIT, DOG1, and desmin by IHC.
PDGFRA mutation status was evaluated in 6 KIT- and DOG1-positive tumors and 2 KIT-negative and DOG1-positive tumors. The 2 KIT-negative and DOG1-positive tumors were also evaluated for c-KIT mutations. All cases yielded amplification products for all exons evaluated, producing a single band when subjected to electrophoresis. All tumors evaluated were wild type for exons 12, 14, and 18 of PDGFRA. No nonsynonymous mutations were identified in exons 8, 9, 11, 13, or 17 of c-KIT. Two heterogeneous and synonymous single nucleotide polymorphisms (SNPs) were detected, 1 each in exons 8 and 11 of c-KIT. Both SNPs have been previously reported. The SNP in exon 8 that was present in both GISTs evaluated as well as in the canine control DNA has been recorded in public databases (rs22299980) and in the literature occurring in canine mast cell tumors, 55 and was also noted in normal canine tissues sequenced for another project (DL Duval, personal communication). The SNP in exon 11 was identified in a single GIST sample in this study and has been previously documented in canine mast cell tumors 55 and canine GISTs. 20
Leiomyosarcoma
A diagnosis of leiomyosarcoma was given for any tumor that was both KIT and DOG1 negative and positive for either desmin or SMA. One tumor that stained positive for KIT (low to moderate intensity; 60% of tumor cells) and negative for DOG1 (dog 48) was diagnosed as leiomyosarcoma based on strong reactivity for desmin (high intensity; 80% of tumor cells). Given the low specificity of SMA, only a few tumors that had equivocal staining for desmin and were negative for the other markers were selected for SMA IHC. Fifteen (27.3%) leiomyosarcomas were diagnosed in 6 (40%) female and 9 (60%) male dogs with an average age of 9.3 years (range: 6–15 years). All leiomyosarcomas were located in the small intestine: 4 in the jejunum, 3 in the duodenum, and the remaining 8 specified only as intestine. None were located in the stomach or the colon. Thirteen leiomyosarcomas were among the 32 tumors previously diagnosed as leiomyosarcoma based solely on anatomic location and histologic features. Eight (53.3%) leiomyosarcomas were desmin positive. Twelve tumors that were negative for KIT and DOG1, and had equivocal desmin staining (indistinct, in less than 25% of tumor cells) were stained for SMA. Of these, 8 (66.7%) were positive for SMA. Interestingly, 3 leiomyosarcomas showed some reactivity for KIT (dogs 17, 20, and 48) with low intensity in ~50–60% of tumor cells. KIT staining in tumors from dogs 17 and 20 was nuclear (and thus, by our criteria, considered KIT negative). All these tumors were DOG1 negative, which is consistent with a similar case described in the human literature. 52
Undifferentiated sarcomas
Tumors negative for all markers were classified as undifferentiated sarcomas. Seven (12.7%) tumors fell into this category from 2 (28.6%) female and 5 (71.4%) male dogs with an average age of 9.6 years (range: 3–14 years). Six (85.7%) tumors were located in the small intestine, with 3 in the duodenum, 1 at the junction of duodenum and jejunum, and the remainder unspecified. One (14.3%) tumor was located in the pylorus. Three of these tumors had previously been diagnosed as leiomyosarcoma based on histology and location alone (2 were grade 3 and 1 was grade 2 based on a commonly used soft tissue sarcoma grading scheme11,31).
Discussion
The accurate diagnosis of human GISTs has become increasingly important given the clinical benefit of the tyrosine kinase inhibitor, imatinib, in the treatment of GISTs with activating c-KIT or PDGFRA mutations.10,24,49 Molecular studies identified the novel protein, DOG1, that is distinct from, but often coexpressed with KIT at high levels in GISTs.39,52 Since then, numerous studies have demonstrated that DOG1 serves as a sensitive and specific marker of GISTs with increased sensitivity for diagnosing GISTs with PDGFRA mutations and other KIT-negative GISTs.14,29,33,40,42 Canine GISTs share similar histological features, immunohistochemical expression, and activating c-KIT mutation (exon 11) with their human counterparts.4,16,20 The canine DOG1 protein has high homology with the human protein (93% amino-acid sequence identity). In this study, we explored the expression of DOG1 in canine gastrointestinal sarcomas and normal intestine and found similar immunoreactivity to that reported in corresponding human tissues. Our results suggest that, for the purpose of differentiating between GIST and leiomyosarcoma, detection of DOG1 via IHC displayed improved sensitivity and specificity compared with IHC for KIT. Similar to reports in the human literature, DOG1 identified at least 2 KIT-negative, epithelioid GISTs.
When specific expression of DOG1 in human GISTs was discovered, little was known about the nature or function of this protein. Since that time, it has been determined that DOG1, a plasma membrane protein with 8 transmembrane domains, is the defining member of a new family of calcium-activated chloride channels (CaCCs).5,54 CaCCs, in response to intracellular calcium levels, control the outflux of chloride necessary for physiologic functions including transepithelial electrolyte secretion and smooth muscle contraction. 21 DOG1 expression is utilized as a specific marker for ICCs in human and rodent studies with evidence that DOG1 contributes to proliferation of ICCs and normal gut contractility.19,47,48 Studies have demonstrated DOG1 activity in GIST cells, with modest effects on cell viability and proliferation in vitro and oncogenic activity in GIST xenografts in vivo.3,46 Amplification of the genomic location of human DOG1 (chromosome band 11q13) has been noted in other cancers including squamous cell carcinomas of the head and neck (SCCHN) where DOG1 overexpression is associated with both a poor prognosis in patients and gain of metastatic characteristics in SCCHN cells. 1
The canine DOG1 protein is located on chromosome 18 and shares high homology with the human protein. In the current study, IHC utilizing a rabbit monoclonal anti-human DOG1 antibody demonstrated comparable immunoreactivity in normal canine intestine, localizing to ICCs between muscular layers, and associated with myenteric plexi, and in canine GISTs. Similar to reports in the human literature, DOG1 was most commonly coexpressed with KIT, with the majority of these tumors exhibiting near diffuse, intense, cytoplasmic to membranous immunoreactivity of both proteins. Additionally, DOG1 membranous staining was demonstrated in 2 KIT-negative tumors exhibiting morphologic characteristics consistent with epithelioid GISTs.
KIT-negative gastrointestinal tumors have been confirmed as GISTs in the human literature based on results of karyotype analysis demonstrating characteristic chromosomal loss.8,12,13,25,36 Despite the lack of a comparable metric for canine GISTs, we propose that the KIT-negative canine tumors represent GISTs based on the following evidence. The histomorphologic features of these tumors are indistinguishable from KIT-positive epithelioid GISTs present in this study population and previously described.16,22 IHC and Western blot results reported herein demonstrating specific immunoreactivity for canine DOG1 in ICCs in normal canine intestinal tissue are consistent with evidence in the human literature for DOG1 as a specific marker for ICCs.19,30,47,48 It is therefore reasonable to conclude that DOG1-positive tumors in the gastrointestinal tract represent tumors arising from cells with features of ICC differentiation, consistent with a diagnosis of GIST and inconsistent with leiomyosarcoma. Additional molecular analyses in DOG1-positive, KIT-negative GISTs may help determine if these tumors truly represent a distinct subtype of GIST arising from a subset of KIT-negative ICCs. Alternatively, these tumors may simply represent cases where the DOG1 antibody–protein interaction was less susceptible to technical factors leading to false-negative KIT immunostaining. In either case, the diagnosis of GIST remains appropriate.
In the human literature, KIT-negative GISTs are more likely to have an epithelioid morphology, and 35–72% harbor PDGFRA mutations (with c-KIT and PDGFRA mutations apparently mutually exclusive).23,29,36 We examined the 3 exons of PDGFRA most commonly mutated in human GISTs in 8 of our 55 canine tumors including 6 KIT-positive GISTs and the 2 DOG1-positive, KIT-negative tumors and failed to identify any mutations. We also examined 5 exons of c-KIT in the 2 KIT-negative canine GISTs and failed to find any amino acid–changing c-KIT mutations. It is not clear whether the apparent absence of PDGFRA mutations in KIT-negative canine GISTs is because of our small sample size or indicates PDGFRA mutations do not occur in canine GISTs. The only other published study looking for PDGFRA mutations in canine GISTs published primer sequences that targeted exons 13, 15, and 19 (discovered during our molecular analysis). 20 Additionally, the GISTs examined in that report were all KIT positive via IHC and so unlikely to harbor PDGFRA mutations, as c-KIT and PDGFRA mutations are mutually exclusive in human GISTs.6,29 One advantage of including DOG1 in IHC panels for diagnosis of GISTs is identification of more cases of DOG1-positive, KIT-negative GISTs. Identification and additional analyses of these tumors are necessary to determine potential clinical significance of reportedly rare KIT-negative and/or epithelioid canine GISTS.
One DOG1-negative tumor was KIT positive using the criteria described in the Material and Methods section. This case was only 1 of 4 tumors weakly positive for KIT (percent cells staining 50–75% and intensity ranging from low to moderate) that did not show positivity for DOG1 and it showed significant immunoreactivity for desmin. We interpreted this case as a leiomyosarcoma with equivocal KIT staining. With this interpretation, DOG1 is more sensitive than KIT for diagnosis of canine GIST consistent with reports in the human literature.14,33,52 Supporting this theory is the finding in our study that, overall, DOG1 staining was more consistent than KIT with all DOG1-positive cases showing immunoreactivity in >75% of cells, including 3 of the 4 previously mentioned weakly KIT-positive cases. Thus, a second advantage of including DOG1 in diagnostic panels for GISTs is the potential to accurately diagnose more GISTs, especially those with equivocal IHC results for KIT.
In human GISTs, DOG1 immunoreactivity has been demonstrated in tumors negative for KIT, but harboring PDGFRA or even rarely c-KIT mutations, thus establishing DOG1 as a valuable marker for identifying KIT-negative GISTs that may benefit from treatment with targeted kinase inhibitors. While the clinical relevance of targeted therapy for canine GISTs has yet to be determined, effectiveness of kinase inhibitors for the treatment of canine mast cell tumors, which are driven by similar c-KIT mutations, has been documented. 34 Additionally, studies have suggested that the prognosis for canine GIST and leiomyosarcoma may be significantly different.22,43 Accurate diagnosis of canine GIST is critical to clarify the clinical behavior of these tumors, identify tumors that may be responsive to novel targeted therapies, and to improve the characterization of canine GIST as a model of human GIST. Based on our results, we suggest the inclusion of DOG1 in IHC panels for the most accurate diagnosis of canine GIST.
Footnotes
Acknowledgements
We would like to thank Brad Charles of the Molecular Pathology laboratory in the Flint Animal Cancer Center at Colorado State University for technical assistance performing Western blot analysis and obtaining photomicrographs, and Marjorie Owen for collection of signalment and history information from archived cases.
a.
ultraView universal alkaline phosphatase red detection kit with the NexES IHC module, Ventana Medical Systems, Tucson, AZ.
b.
Novocastra Bond polymer refine red detection kit with the Novocastra Bond research detection system, Leica Biosystems, Buffalo Grove, IL.
c.
Monoclonal rabbit anti-human DOG1 (SP31), Cell Marque, Rocklin, CA.
d.
Monoclonal mouse anti-human desmin (DE-R-11), Leica Biosystems, Buffalo Grove, IL.
e.
Polyclonal rabbit anti-human CD117, c-kit; Dako North America Inc., Carpinteria, CA.
f.
Monoclonal mouse anti-human smooth muscle actin (asm-1), Leica Biosystems, Buffalo Grove, IL.
g.
SuperBlock T20, Thermo Scientific, Rockford, IL.
h.
Complete protease inhibitor cocktail, Roche Diagnostics, Indianapolis, IN.
i.
Goat anti-rabbit IgG secondary antibody HRP conjugate, Thermo Scientific, Rockford, IL.
j.
SuperSignal West Pico substrate, Thermo Scientific, Rockford, IL.
k.
GoTaq Flexi DNA polymerase, Promega Corp., Madison, WI.
l.
GelGreen, Biotium Inc., Hayward, CA.
m.
QIAquick gel extraction kit, Qiagen Inc., Valencia, CA.
n.
ABI 3130xL genetic analyzer, Life Technologies, Grand Island, NY.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.