
Abstract
Besnoitiosis is an emerging infectious disease of donkeys (Equus asinus) in the United States for which there are currently no serologic methods of diagnosis. A study was performed to evaluate physical examination findings and 3 serologic assays for the detection of Besnoitia bennetti infection in donkeys. A prospective study of 416 donkeys from 6 privately owned herds across 5 U.S. states (New York, Pennsylvania, Vermont, Oregon, and Washington) was performed. Donkeys were examined for clinical lesions suggestive of besnoitiosis and evaluated for antibodies against B. bennetti using a fluorescent antibody test (FAT) and 2 immunoblot assays specific for bradyzoite and tachyzoite antigens, respectively. Donkeys were confirmed to be infected with B. bennetti by histology (cases; n = 32) and were compared to those with no clinical signs of besnoitiosis (controls; n = 384). Identifying clinical lesions in 2 or more locations correctly identified infected donkeys 83% of the time. Donkeys with besnoitiosis had significantly higher FAT titers (P < 0.001) and numbers of bradyzoite (P < 0.001) and tachyzoite (P < 0.001) immunoblot bands than control donkeys. The sensitivity and specificity of the serologic assays for detecting besnoitiosis was 88% and 96% for FAT, 81% and 91% for bradyzoite immunoblot, and 91% and 92% for tachyzoite immunoblot, respectively. Fluorescent antibody and immunoblot assays are effective at identifying donkeys with besnoitiosis and provide a more efficient and less invasive diagnostic alternative to histology.
Besnoitiosis is caused by infection with Besnoitia spp. protozoa, which are host-specific, cyst-forming coccidians that affect multiple host species worldwide.1–3,7,10 Clinical disease is characterized by pinpoint parasitic nodules in the skin over the face and body, within the nares, on the internal and external pinnae, and on the legs and perineum (Fig. 1).1,4,6,9 Infected animals may also develop nodules along the limbal margin of the sclera, termed scleral pearls (Fig. 2).1,4,6,9 Bovine besnoitiosis, caused by Besnoitia besnoiti, has emerged as a disease of economic importance in European cattle, and is currently considered endemic in several countries, including France, Spain, Portugal, and Italy, with sporadic cases reported in Germany and Switzerland. It is characterized by generalized skin lesions, decreased milk production, transient or permanent male infertility, and increased mortality.5,6,10,11 Cervine besnoitiosis, caused by Besnoitia tarandi, has been described in reindeer (Rangifer tarandus tarandus) and caribou (Rangifer tarandus caribou) in Canada, Alaska, Finland, and Sweden. 2 Equine besnoitiosis, caused by Besnoitia bennetti, was historically limited to donkeys and horses in Africa; however, reports have suggested that besnoitiosis may be an emerging disease of donkeys (Equus asinus) in the United States.1,4,9,10
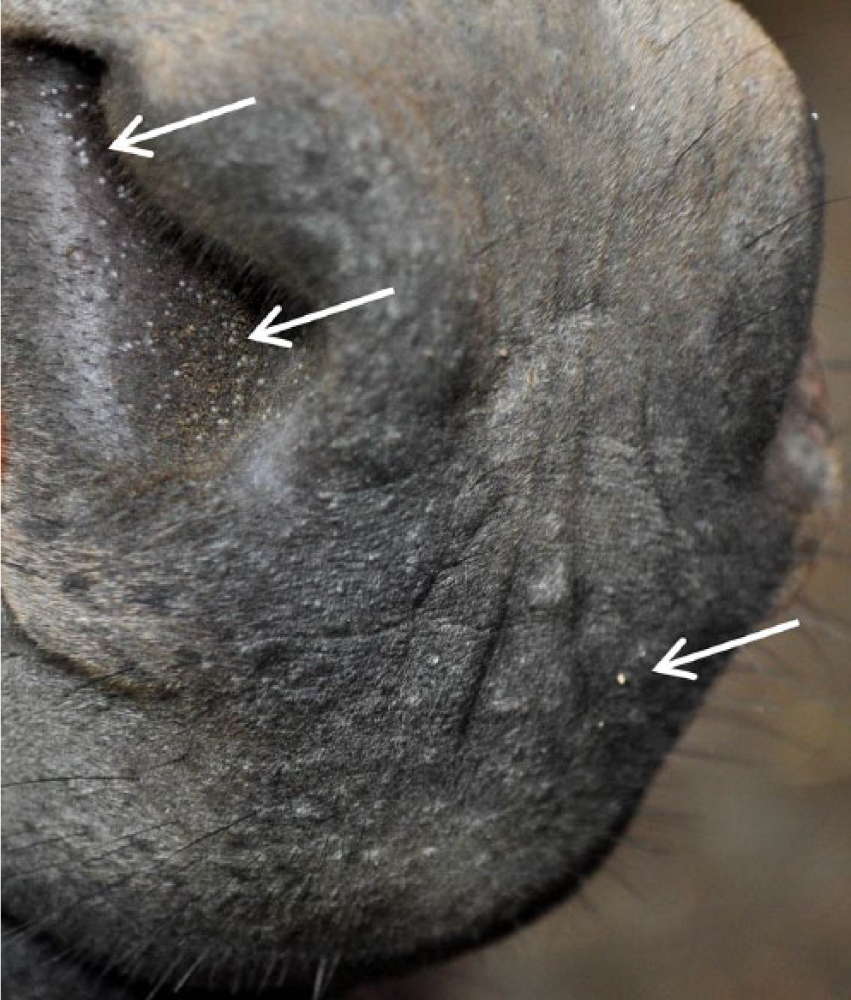
Nares and muzzle lesions (arrows) in a donkey (Equus asinus) with besnoitiosis.
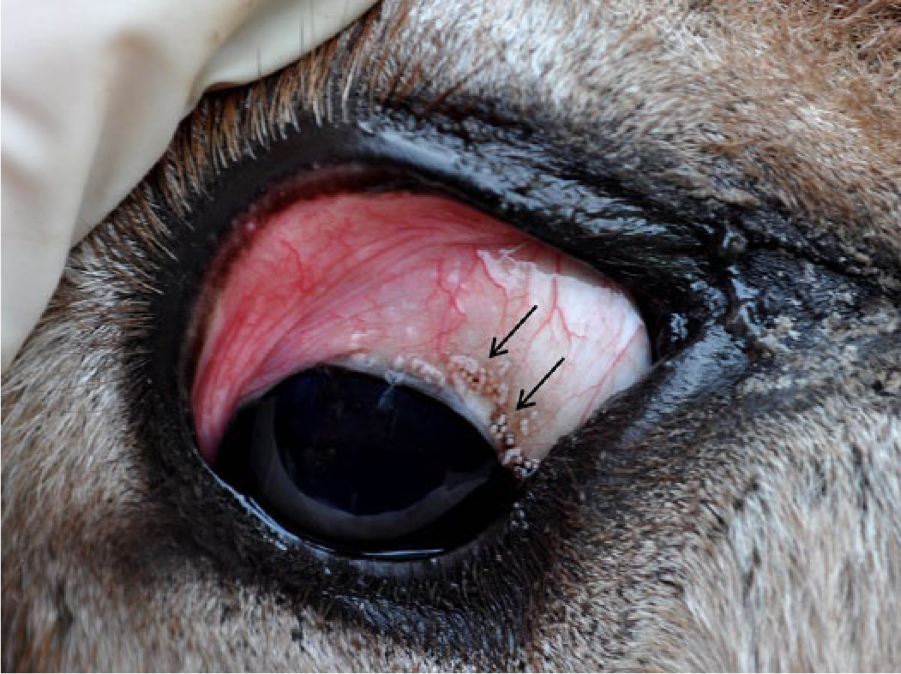
Scleral pearls (arrows) in the right eye of a donkey (Equus asinus) with besnoitiosis. Note the concurrent conjunctivitis and crusting ocular discharge that is frequently observed in affected animals.
The life cycle of Besnoitia spp. involves both a definitive (predator) and intermediate (prey) host. A feline definitive host has been identified for Besnoitia oryctofelisi, Besnoitia darlingi, and Besnoitia neotomofelis, which affect the rabbit (Oryctolagus cuniculus), Virginia opossum (Didelphis virginia), and southern plains woodrat (Neotoma micropus), respectively. 10 Attempts to demonstrate the cat, or any other animal, as the definitive host in equine besnoitiosis have been unsuccessful, and the definitive host, life cycle, and mode of transmission of Besnoitia spp. in equids and ruminants remain unknown.1,10
The emergence of bovine besnoitiosis in Europe has prompted the development of several serologic and molecular techniques for the diagnosis of besnoitiosis in cattle.5,12,13 Phylogenetically, B. besnoiti, B. tarandi, and B. bennetti are similar and differ primarily by biological traits such as intermediate host specificity. Although microsatellite markers have been identified showing differences between field isolates from B. besnoiti, B. bennetti, and B. tarandi, 8 there are only minor differences in the ribosomal (r)RNA genes of these 3 Besnoitia spp., and rRNA provides no molecular markers to distinguish them on a genetic level.1–3,11 The current gold standard for diagnosing besnoitiosis in donkeys is identification, via skin biopsy, of characteristic Besnoitia spp. within the dermis of individuals with clinical lesions (Fig. 3).1,4,9 Validation of efficient and minimally invasive methods for identifying infected donkeys may strengthen efforts to prevent the spread of besnoitiosis within donkey populations, as infected animals may serve as reservoirs for the parasite, and the introduction of infected animals into naive herds likely plays a role in transmission of infection within herds.6,10
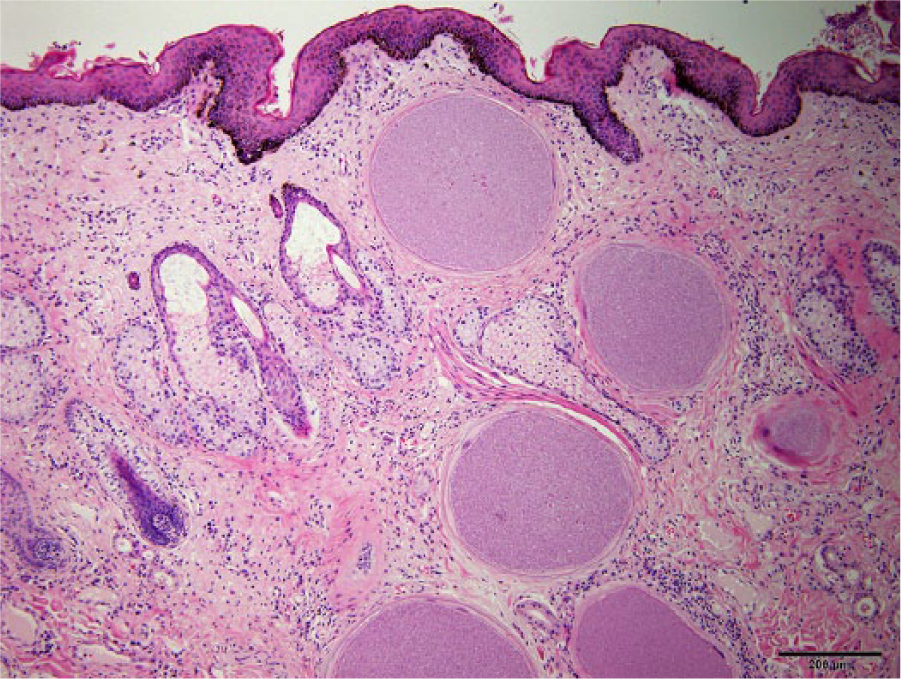
Besnoitia spp. tissue cysts within the dermis of a miniature donkey (Equus asinus). Note 6 lesions, each containing bradyzoite stage parasites. Some lesions are surrounded by mild lymphoplasmacytic infiltrates. Magnification 40×. Bar = 200 µm.
The aim of the present study was to evaluate the utility of physical examination and 3 serologic assays validated for use in cattle 12 for the detection of B. bennetti infection in donkeys. It was hypothesized that, due to phylogenetic similarities between B. besnoiti and B. bennetti, serologic assays developed for the detection of antibodies against B. besnoiti in cattle would cross-react and correctly identify antibodies against B. bennetti in donkeys.
The current study prospectively examined 416 donkeys from 6 privately owned herds across 5 U.S. states (New York, Pennsylvania, Vermont, Oregon, and Washington). Premises were visited by 1 or more of the authors (SL Ness, LD Mittel) between October 2010 and October 2012. Owner consent was obtained for all examinations and procedures. The nares and perineum of all donkeys on each property were examined for pinpoint, white, nodular lesions suggestive of besnoitiosis. Both eyes were inspected for scleral pearls. The sites were chosen based on previous work demonstrating the nares and sclera as the most common locations for Besnoitia spp. lesions in donkeys. 9 The presence and location of lesions were recorded when observed. Skin biopsies were obtained from all animals identified with lesions suggestive of besnoitiosis and preserved in 10% neutral buffered formalin solution for histologic examination performed by one of the authors (J Peters-Kennedy). Biopsy sites were selected to maximize the number of lesions obtained within the sampled tissue. Blood was collected from every donkey and evaluated for antibodies against B. bennetti using a fluorescent antibody test (FAT) and 2 immunoblot assays specific for bradyzoite and tachyzoite antigens, respectively, as previously described. 12
Briefly, suspensions of B. besnoiti Bb1Evora03 tachyzoites (5 × 106 tachyzoites/ml) in phosphate buffered saline (PBS) solution (pH 7.2) were air-dried on glass slides and frozen at −20°C until used. After incubation in serum, the slides were rinsed with fluorescent antibody buffer (25 mM Na2CO3, 100 mM NaHCO3, and 35 mM NaCl; pH 9.0) and PBS solution prior to conjugate incubation. Fluorescein isothiocyanate–conjugate anti-horse IgG (Heavy + Light [H + L] chains), a diluted 1:50 in PBS solution with 0.05% Evans blue was added, and the slides were examined with a fluorescence microscope. b Only peripheral but not apical fluorescence was considered specific. To prepare antigen-coated membranes by Western blotting, samples containing 4 × 107 B. besnoiti zoites (either cell culture–derived Bb1Evora03 tachyzoites or bradyzoites released from the skin of infected cattle 12 ) were treated for 10 min at 94°C with nonreducing sample buffer (2% [weight/volume] sodium dodecyl sulfate [SDS], 10% [v/v] glycerol, and 62 mM Tris–HCl; pH 6.8) and electrophoresed in a SDS–polyacrylamide minigel. Separated parasite antigens and marker proteins were electrophoretically transferred to polyvinylidene fluoride membranes c in a semidry transfer system. d The antigen-coated membrane was blocked with PBS-TG e containing PBS with 0.05% (v/v) Tween 20 and 2% (v/v) liquid fish gelatin, air-dried overnight, cut into strips, and stored frozen at −20°C until used. Prior to immunoblot analysis, serum samples were diluted 1:200 in PBS-TG, and the strips were blocked again with PBS-TG. After washing in PBS solution with 0.05% (v/v) Tween 20, the strips were incubated with peroxidase conjugate solution (affinity purified goat anti-horse IgG [H+L], a diluted 1:500 in PBS-TG). After washing in PBS-TG and distilled water, antibody reactions were detected by adding substrate solution (40 µl of H2O2 [30% {v/v}] and 30 mg of 4-chloro-1-naphthol f in 40 ml of PBS solution and 20% [v/v] methanol). Preinfection and postinfection sera from a heifer experimentally infected with Neospora caninum 14 were used as negative controls, and sera of 3 cattle naturally infected with Besnoitia 13 were used as positive controls. To detect antibodies to Besnoitia, affinity-purified goat anti-horse IgG (H + L) a diluted 1:500 in PBS-TG was used. In both the Besnoitia tachyzoite and bradyzoite immunoblot assays, diagnosis was made on the basis of 10 selected specific bands, as previously described. 12 Patterns of band positivity within the 2 immunoblot assays were not analyzed, but the number of positive bands in each of the assays was recorded.
Clinical and serologic findings in donkeys confirmed to be infected with B. bennetti by histology (cases) were compared to those with no clinical signs of besnoitiosis (controls). Data was evaluated for normality using the Shapiro–Wilk test and summarized as median and range. Proportions were compared by Fisher exact test. The Wilcoxon rank sum test was used to compare results of the FAT, bradyzoite immunoblot, and tachyzoite immunoblot between cases and controls. Sensitivity, specificity, and positive and negative predictive values (PPVs and NPVs, respectively) with 95% confidence intervals were calculated for each assay. Receiver operating characteristic (ROC) curves were derived for each assay, and the area under the curve (AUC) was calculated. The ROC analysis was used to assess the ability of the FAT, bradyzoite immunoblot, and tachyzoite immunoblot to identify animals with besnoitiosis, and to help estimate the optimal diagnostic cutoffs that would best classify cases correctly. All analyses were performed with a commercially available software program, g and P ≤ 0.05 was considered significant.
A total of 416 donkeys were enrolled in the present study. The study population comprised 311 (75%) females, 85 (20%) sexually intact males, and 20 (5%) castrated males, reflecting a primarily breeding-operation population. The following breeds were included: 391 Miniature (94%), 9 Standard (2%), 8 Poitou (2%), 6 Mule (1.5%), and 2 Mammoth (0.5%). Median overall age was 48 months (range: 1–336 months). Within the study population, 32 donkeys were identified as infected with B. bennetti by histology (cases), of which there were 26 (81%) females, 5 (16%) sexually intact males, and 1 (3%) castrated male. The median age of donkeys with besnoitiosis (24 months; range: 9–156 months), was significantly lower than the median age of control donkeys (60 months; range: 1–336 months; P = 0.002). Neither sex nor breed was significantly associated with besnoitiosis.
Lesions in the nares and perineum were identified in 30 out of 32 (94%) and 22 out of 32 (69%) infected donkeys, respectively. Scleral pearls were present in 26 out of 32 (81%) infected donkeys. Identifying clinical lesions in either the nares or the perineum correctly identified an infected donkey 53% of the time, while identifying scleral pearls identified an infected donkey 70% of the time. Identifying lesions in 2 or more locations (e.g., nares and sclera) correctly identified infected donkeys 83% of the time, regardless of lesion location.
Donkeys with besnoitiosis had significantly higher FAT titers (median: 800; range: 0–3,200) than control donkeys (median: 0; range: 0–800; P < 0.001). A FAT-positive cutoff value of ≥400 provided an 88% sensitivity, 96% specificity, 65% PPV, and 99% NPV (Table 1). Donkeys with besnoitiosis were positive for more bradyzoite-specific bands (median: 6; range: 1–10) than control donkeys (median: 0; range: 0–7; P < 0.001). When a positive cutoff was set at the detection of 4 or more bradyzoite-specific antigen bands, the bradyzoite-based immunoblot performed with 81% sensitivity, 91% specificity, 42% PPV, and 98% NPV (Table 1). Donkeys with besnoitiosis were positive for more tachyzoite-specific bands (median: 5; range: 0–9) than control donkeys (median: 0; range: 0–7; P < 0.001). When a positive cutoff was set at the detection of 4 or more tachyzoite-specific antigen bands, the tachyzoite-based immunoblot performed with 91% sensitivity, 92% specificity, 50% PPV, and 99% NPV (Table 1). The ROC analysis revealed that the FAT (AUC: 0.96), bradyzoite immunoblot (AUC: 0.94), and tachyzoite immunoblot (AUC: 0.95) were all useful for correctly identifying donkeys with besnoitiosis (Fig. 4). Pairwise comparisons of ROC curves revealed no significant differences in diagnostic performance between the 3 assays.
Receiver operating characteristic curve analysis with area under the curve (AUC), sensitivity, specificity, positive and negative predictive values (PPVs and NPVs, respectively), and 95% confidence interval (CI) for fluorescent antibody test (FAT) and bradyzoite-specific and tachyzoite-specific immunoblot assays for identifying donkeys with besnoitiosis.
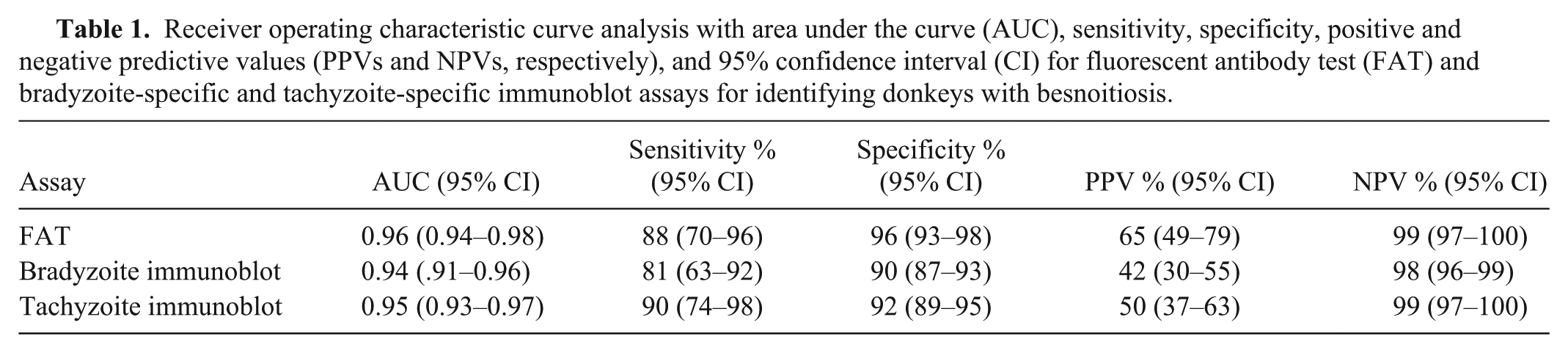
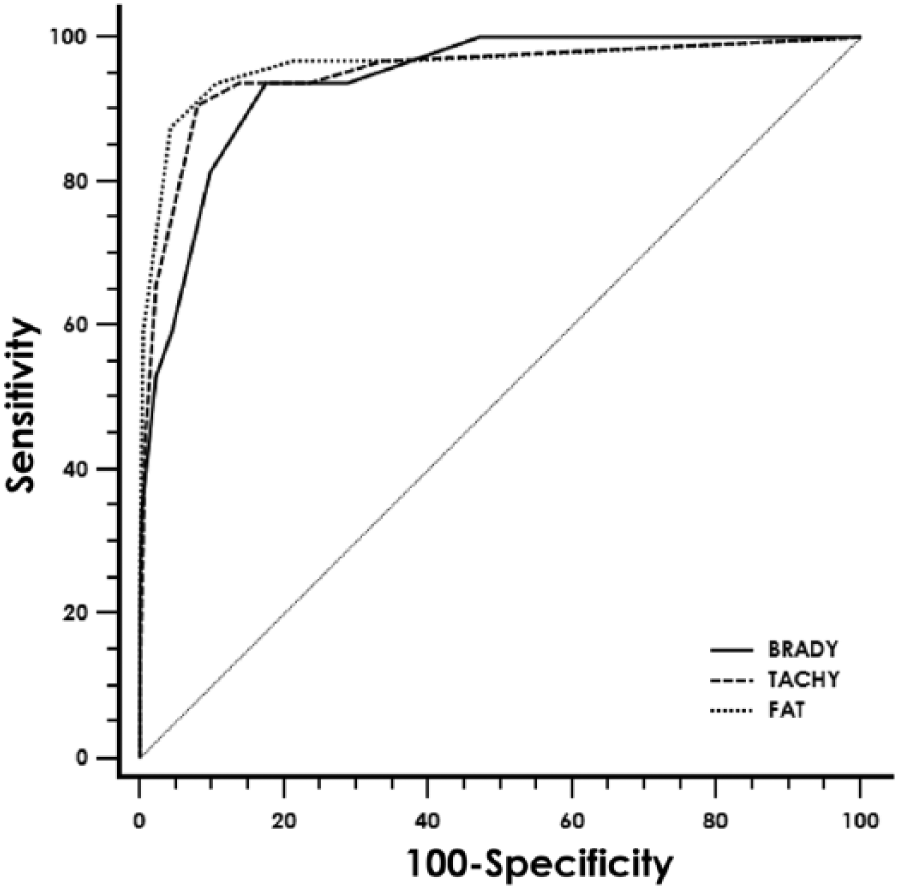
Receiver operating characteristic (ROC) curve illustrating the sensitivity and specificity of fluorescent antibody test (FAT) and bradyzoite-specific and tachyzoite-specific immunoblot assays to identify donkeys (Equus asinus) with besnoitiosis. The area under the ROC curves are as follows: FAT, 0.96; bradyzoite-specific immunoblot, 0.94; tachyzoite-specific immunoblot, 0.95. Pairwise comparisons of ROC curves revealed no significant differences in diagnostic performance between the 3 assays.
In the present study, FAT and immunoblot assays were able to identify donkeys with besnoitiosis and performed with similar sensitivity and specificity to that which has been previously reported for cattle. 12 The assays provide an alternative to the current method of physical examination and histology, and have the advantage of being less labor-intensive and less invasive. Moreover, accurate identification of Besnoitia spp. lesions on physical examination requires that the examiner has a working knowledge of the clinical presentation of besnoitiosis, and was only effective at identifying infected donkeys 83% of the time in the present study.
In the current study population, young animals were at increased risk of infection with Besnoitia spp. This finding has been previously reported in donkeys.4,9 The potential for recovery from besnoitiosis in infected donkeys is currently unknown, but it is possible that the development of acquired immunity after exposure or infection is a relevant factor in the observed age discrepancy. In contrast to donkeys, seropositivity has been reported to increase with age in cattle. 6 The reason for the difference in these 2 host species is not currently known. Neither sex nor breed was significantly associated with besnoitiosis in the present study. Male donkeys have been previously reported to be at increased risk for developing besnoitiosis. 9 Sex predisposition in cattle is controversial, with some studies reporting that females are at higher risk for infection, whereas others report similar findings for males. 6 It is important to note that the disproportionately high number of female Miniature donkeys in the study population presented herein may have prevented objective analysis.
The results of the current study also highlight the variability in host serologic response to infection and the limitations of using serology alone to identify donkeys with besnoitiosis. Within this study population, 2 of the 32 confirmed cases displayed FAT and immunoblot profiles less than the positive cutoff values despite having Besnoitia spp. tissue cysts confirmed on histology. Notably, one of these animals demonstrated almost no serologic response at all (FAT titer: 0, tachyzoite immunoblot: 0, bradyzoite immunoblot: 1) despite severe clinical infection characterized by widespread tissue cysts in the skin, mucous membranes, and laryngeal, pharyngeal, and tracheal mucosa, as well as generalized cachexia, malaise, and weakness. Marked variability in clinical presentation and serologic response has been reported in cattle with besnoitiosis, including a report 13 in which 2 clinically positive animals with FAT titers <50 tested positive for besnoitiosis by polymerase chain reaction assay. The reason for this variability in host response is not currently understood.
Fifty-three of the 384 control donkeys demonstrated serologic values greater than the positive cutoff on 1 or more of the serologic assays, resulting in the relatively low PPVs noted in this study. The relationship between seroconversion and the development of clinical lesions following infection in donkeys is not currently understood; however, at least 1 of the seropositive but clinically negative (i.e., lacking clinical lesions) donkeys went on to develop Besnoitia spp. lesions in the months following initial evaluation. Thus, continued monitoring of any donkey found to be serologically positive in the absence of clinical lesions is recommended.
In conclusion, FAT and immunoblot assays are effective at identifying donkeys with besnoitiosis and provide a more efficient and less invasive diagnostic alternative to histology. The findings of the current study will aid in the diagnosis and surveillance of besnoitiosis in donkeys and will further the understanding of the epidemiology and transmission of this disease in the United States
Footnotes
Acknowledgements
The authors thank the owners of the donkey herds enrolled in this study, and Jan Liotta, Andrea Bärwald, and Lieselotte Minke for technical assistance.
a.
Jackson ImmunoResearch Laboratories Inc., West Grove, PA.
b.
Vanox AHBT3, Olympus Deutschland GmbH, Hamburg, Germany.
c.
Millipore Immobilon-P, Merck Chemicals GmbH, Hessen, Germany.
d.
Pharmacia Biotech, Freiburg, Germany.
e.
SERVA Electrophoresis GmbH, Heidelberg, Germany.
f.
Sigma-Aldrich Chemie GmbH, Munich, Germany.
g.
Statistix version 9.0, Analytical Software Inc., Tallahassee, FL.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant from National Miniature Donkey Association.