
Abstract
Respiratory infections are a major burden in the performance horse industry. Equine rhinitis B virus (ERBV) has been isolated from horses displaying clinical respiratory disease, and ERBV-neutralizing antibodies have been detected in 50–80% of horses in reported surveys. Current ERBV isolation and detection methods may underestimate the number of ERBV-positive animals and do not identify multiple serotype infections. The aim of the current study was to develop a serotyping ERBV antibody-detection enzyme-linked immunosorbent assay (ELISA) and examine the seroprevalence of ERBV in a group of Australian weanling horses. ELISAs with high sensitivity and specificity were developed. The seroprevalence of ERBV in the weanling horses was high (74–86%); ERBV-3 antibodies were most prevalent (58–62%) and ERBV-2 antibodies were least prevalent (10–16%). Many horses were seropositive to 2 or more serotypes. All 3 serotypes of ERBV were detected, and concurrent positivity to multiple serotypes was common.
Keywords
Respiratory disease in horses is of major economic importance, particularly in the performance horse industry. One of the lesser known equine respiratory viruses is Equine rhinitis B virus (ERBV; order Picornavirales; family Picornaviridae; genus Erbovirus). Equine rhinitis B virus has been isolated from horses with clinical signs, including fever to 41°C for 1–3 days, nasal discharge, edema of the legs, lethargy, and swelling of lymph nodes (Hofer B, Steck F, Gerber H, et al.: 1972, An investigation of the aetiology of viral respiratory disease in a remount depot. In: Proceedings of the third International Conference of Equine Infectious Diseases, pp. 527–545. Paris, France; Mumford JA, Thomson GR: 1978, Studies on picornaviruses isolated from the respiratory tract of horses. In: Proceedings of the fourth International Conference on Equine Infectious Diseases, ed. Bryans JT, Gerber H, pp. 418–429. Veterinary Publications, Princeton, NJ), 6 and has been shown to establish persistent infection, with virus isolated from horses over an 18–24-month period (Burrows R, Goodridge D: 1978, Observations of picornavirus, adenovirus, and equine herpesvirus infections in the Pirbright pony herd. In: Proceedings of the fourth International Conference on equine Infectious Diseases, ed. Bryans JT, Gerber H, pp. 155–164. Veterinary Publications, Princeton, NJ; Mumford JA, Thomson GR: 1978).
There are 3 ERBV serotypes (ERBV-1, -2, and -3), which are differentiated on the basis of acid lability or stability, genetic sequences, and neutralization by type-specific antisera. The ERBV capsid comprises 4 virion proteins—VP1, VP2, VP3, and VP4. Previously, it has been shown that the VP1 proteins, and more specifically the C-termini of VP1 (VP1-Ct), of each ERBV serotype are the location of serotype-specific B-cell epitopes. 9 Isolates of the same serotype have a high level of amino acid sequence identity in this region (81–100%).
The 3 ERBV serotypes are present in horse populations worldwide, reflected by both virus isolation (McCollum WH, Timoney PJ: 1992, Studies on the seroprevalence and frequency of equine rhinovirus 1 and 2 infection in normal horse urine. In: Proceedings of the sixth International Conference on Equine Infectious Diseases, ed. Plowright W, Rossdale PD, Wade JF, pp. 83–87. R and W Publications, Newmarket, UK; Steck F, Hofer B, Schaeren B, et al.: 1978, Equine rhinoviruses: new serotypes. In: Proceedings of the fourth International Conference on Equine Infectious Diseases, ed. Bryans JT, Gerber H, pp. 321–328. Veterinary Publications, Princeton, NJ)1,2,4-6,15 and a high seroprevalence of ERBV antibodies (Holmes DF, Kemen MJ, Coggins L: 1978, Equine rhinovirus infection—serologic evidence of infection in selected United State horse populations. In: Proceedings of the fourth International Conference on Equine Infectious Diseases, ed. Bryans JT, Gerber H, pp. 315–319. Veterinary Publications, Princeton, NJ; McCollum WH, Timoney PJ: 1992; Wernery U, Wernery R, Zachariah R, Hayden-Evans J: 1998, Serological survey of some equine infectious diseases in the United Arab Emirates. In: Proceedings of the eighth International Conference on Equine Infectious Diseases, ed. Wernery U, Wade JF, Mumford JA, Kaaden OR, pp. 367–370. R and W Publications, Newmarket, UK).2-4,15 Virus neutralization (VN) assays have detected antibodies to ERBV-1 and ERBV-2 in 83% and 66% of horses tested in Australia, respectively. 1 At present, no data is available on the seroprevalence of ERBV-3, as this serotype was isolated for the first time in Australia only recently (2011). 8 To the authors’ knowledge, there is no rapid method for determining the presence of ERBV antibodies in horse serum. An ERBV enzyme-linked immunosorbent assay (ELISA) has been reported elsewhere that uses the denatured VP1 protein as an antigen; however, this assay was developed only for the detection of ERBV-1. 11
In the current study, a serotyping ERBV antibody-detection ELISA was developed and used to examine the seroprevalence of ERBV antibodies in a group of 50 weanling horses from 2 farms in a key horse breeding region in New South Wales, Australia. The methodology employed in the ELISAs was similar to that described previously. 12 The ERBV fusion proteins used as antigen in each serotype ELISA were prepared as described previously. 9 The sequences for these proteins were derived from the prototype ERBV-1 strain 1436/71 (GenBank accession no. NC_003983), the prototype ERBV-2 strain 313/75 (NC_003077), and the Australian ERBV-3 isolate 2225AS (GU799328). For ERBV-1 and ERBV-2, the VP1-Ct (correlating to nucleotides 748-972 in ERBV-1 VP1 and nucleotides 745-957 in ERBV-2 VP1) expressed with a GST tag was used and, for ERBV-3, the full-length VP1 expressed with a hexahistidine tag was used. These proteins were selected based on their reactivity to serotype-specific rat and horse sera using Western blot and ELISA, 9 and the correlation of ELISA absorbance with VN titer. For ERBV-1 and -2, results were similar when using either the full-length VP1 or VP1-Ct as antigen; however, as full-length VP1 is more difficult to express than VP1-Ct and requires denaturation and refolding, VP1-Ct was chosen as the antigen for the ERBV-1 and -2 ELISAs. For ERBV-3, VP1-Ct showed poorer correlation with VN results than full-length VP1 and consequently full-length VP1 was chosen as the antigen for this ELISA.
To validate the ELISAs, 60 sera from naturally infected horses were used. The sera had varying serotype specificities as defined by VN. Sera were grouped according to their VN titers to ERBV-1, -2, or -3 for analysis of ERBV-1, -2, or -3 antigens, respectively. A serum was considered as negative if the neutralizing titer was less than 10. This cutoff was determined based on the neutralizing titer of serum from specific pathogen free horses and the cytotoxicity observed with higher concentrations of sera. Experimentally prepared reference sera were unavailable. Each serum was tested in triplicate. An unrelated His-tagged protein (His-neg: for full-length VP1) or GST alone (for VP1-Ct) were used as control proteins for background absorbance, which was subtracted from the absorbance value of the antiserum against the ERBV proteins.
The optimal coating antigen concentration and serum dilution were 0.5 µg/ml and 1 in 100, respectively, based on cross-titration using pooled sera from 2 horses with high, monospecific VN titers to ERBV-1, -2, or -3 (data not shown). Inter- and intra-assay variability was calculated using sera representing a high (VN titer <80), mid-range (VN titer 40–80), and low positive (VN titer 10–40), and negative (VN titer <10) for each serotype. Absorbances varied in the range of 1.6–16%, indicating a high level of reproducibility in each assay. Analysis of reactivity after preabsorption with the homologous or heterologous proteins indicated that the assays were detecting serotype-specific antibody and that there are no significant cross-reactive epitopes between the ELISA antigens of different ERBV serotypes (data not shown).
The absorbance values of the 60 sera and VN titer to homologous virus were compared for each serotype, and the cutoff absorbance values for positivity were determined by receiver operating characteristic analysis (Table 1). Using these parameters, contingency tables were prepared, and the level of agreement in each assay was assessed based on the calculation of kappa values. The kappa coefficient is a statistical measure of interrater agreement for qualitative items. For all ELISAs, the sensitivity and specificity were high, and the agreement between VN and ELISA was “good” (Table 1).
Analysis of the enzyme-linked immunosorbent assays (ELISAs) for each Equine rhinitis B virus (ERBV) serotype and agreement with virus neutralization results.*
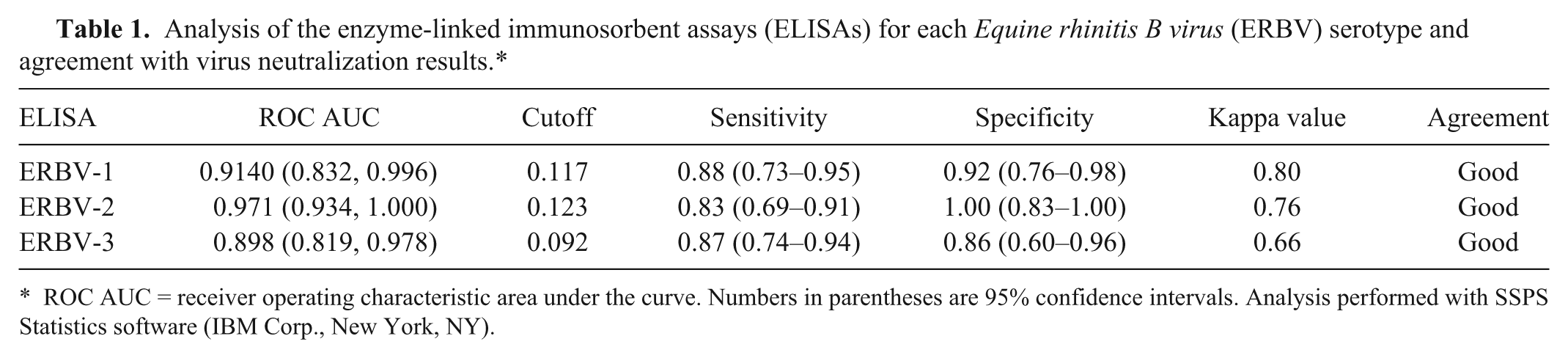
ROC AUC = receiver operating characteristic area under the curve. Numbers in parentheses are 95% confidence intervals. Analysis performed with SSPS Statistics software (IBM Corp., New York, NY).
The seroprevalence of antibodies to ERBV-1, -2, and -3 was examined in archived sera from 50 weanling Thoroughbred horses located on 2 adjacent farms in the Hunter Valley in New South Wales. 7 Three serum samples from each horse were used, collected when the animals were 4–6, 7–9, and 11–13 months of age. A sample was determined as seropositive when the ELISA absorbance value was greater than the cutoff absorbance value. The proportion of weanlings seropositive to each ERBV serotype in each age-group is summarized in Table 2. At each time point, the levels of positivity of each of the 3 serotypes were diverse, including both strong and weak positives (see Supplementary figure).
Equine rhinitis B virus (ERBV) seropositivity in weanlings.*
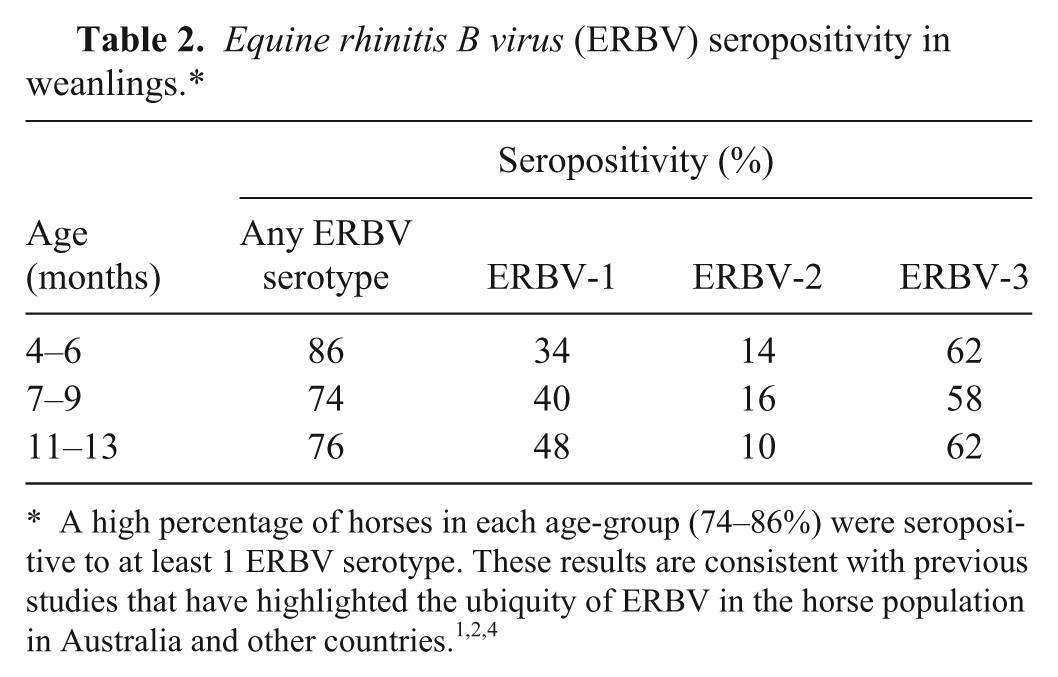
Despite some horses showing a decrease in ERBV antibody and others showing an increase over the study period, the number of horses positive to each serotype at any 1 time was consistent. ERBV-2 had the lowest prevalence (10–16%); ERBV-1 was found in 34–48% of horses; and interestingly, ERBV-3 was the most prevalent, with antibodies detected in 58–62% of animals in each age group. Over the 9-month period, 56% of the weanlings were seropositive to 2 ERBV serotypes at the same time, and 12% were seropositive to all 3 ERBV serotypes. This suggests that all serotypes are widespread and were actively circulating at the time of sampling. This may also be an indication of coinfection with different serotypes, as described previously. 8
When examining the prevalence of ERBV in the weanlings, a number of patterns of infection emerged. Positivity in all bleeds occurred most frequently for ERBV-3, whereas the majority of the weanlings (n = 36) were negative in the ERBV-2 ELISA in all bleeds. An increase in antibody levels indicating seroconversion was observed in 22 horses. Seroconversion to all ERBV serotypes was observed in individual horses and, in 5 animals, appeared to occur to 2 serotypes simultaneously, providing further evidence for coinfection with different serotypes.
In many cases, antibody was present in the first bleed, decreasing in consecutive bleeds. This may be the slow waning of maternal antibody, though previous studies have reported that the decline of maternal antibody in foals occurs prior to 4 months of age.10,16 In addition, the levels of antibody in the first bleeds (4–6 months) were as high as those seen after seroconversion and were significantly higher than would be expected for any remaining maternal antibody at this time point. Taken together, it is likely that active infection of these animals occurs at a very young age (<4 months).
Some animals were positive at 4–6 months and, after the dissipation of this antibody by 7–9 months, showed a return to positivity at 11–13 months. This is consistent withre-infection with the same serotype and may imply poor maintenance of protective antibodies or the existence of diverse subtypes of each serotype in the population. Persistence leading to recrudescence, as well as re-infection with ERBV, particularly in young horses, have been suggested as significant features in the epidemiology of these viruses (Burrows R, Goodridge D: 1978, Observations of picornavirus, adenovirus, and equine herpesvirus infections; Steck F, et al.: 1978, Equine rhinoviruses). No correlation was seen between conversion to ERBV positivity and month of the year, dismissing seasonal factors as contributors to increased infection rates in the present study.
The serotype-specific ERBV antibody detection ELISAs developed in the current study revealed the prevalence of all 3 ERBV serotypes in Australian horses. Previous studies have been limited to ERBV-1 and -2.1,5 In contrast to the high seroprevalence of ERBV, isolation and detection in nasal or oral swabs is infrequent. Studies using reverse transcription quantitative polymerase chain reaction have detected ERBV in only 1.5% 14 and 16% 13 of nasal swabs tested, suggesting the levels of virus in nasal swabs may be too low for regular detection and using these methods alone may underestimate the number of ERBV-positive animals. These methods also do not identify multiple serotype infections. A serotyping ELISA for ERBV is a particularly important and useful tool, and larger screens of horse sera will provide greater insight into the prevalence of ERBV in Australia.
Footnotes
Acknowledgements
The authors thank Nino Ficorilli and Cynthia Brown for technical assistance.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Jacquelyn Horsington was the recipient of a Melbourne Research Scholarship. Other funding support was from the Special Virology Fund, Centre for Equine Virology, The University of Melbourne.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.