
Abstract
During a 6-year period, 62 out of 2,980 calves examined (2.08%) between 1–30 days of age had lesions consistent with Bovine herpesvirus 1 infection. The mean and median age of affected calves was 14 days of age. Most cases of infection were in calves 12 and 15 days of age, with 34 calves (54.8%) affected; 14 calves (22.6%) were between 16–21 days of age; 11 calves (17.8%) were between 7–11 days of age; and 3 calves (4.8%) were between 22–30 days of age. In 29 of the calves, the adrenal gland was the only organ affected. In 25 calves, the adrenal gland and at least 1 other tissue had lesions consistent with herpes viral infection. Five animals had no adrenal lesions but had 1 or more additional tissue with lesions consistent with herpesvirus. Three animals had no adrenal gland present for examination but had lesions in other tissues. Organs and number of animals affected by Bovine herpesvirus 1 infection were adrenal gland (54), liver (18), lungs (15), kidney (8), small intestines (7), large intestines (7), rumen/omasum (7), trachea/larynx (3), abomasum (2), heart (1), esophagus (1), thymus (1), and lymph node (1).
Bovine herpesvirus 1 (BHV-1; order Herpesvirales, family Herpesviridae, subfamily Alphaherpesvirinae, genus Varicellovirus) is the cause of infectious bovine rhinotracheitis, which is widespread in cattle and often associated with subclinical infections. The incidence of clinical disease has decreased due to routine vaccination of both dairy and beef animals. Respiratory infections are usually seen in unvaccinated stressed animals involving the upper respiratory tract and lungs and can vary from subclinical to severe infections. Young animals may also develop keratoconjunctivitis with or without upper respiratory disease. Disseminated infections involving multiple organs have also been identified in neonatal calves and fetuses infected with BHV-1.1-3,8-10 Bovine herpesvirus 1 can also cause pustular vulvovaginitis and balanoposthitis in young adult animals.6,7
Systemic infections in neonatal calves have been reported, with lesions seen primarily in the upper respiratory tract, lungs, oral cavity, esophagus, and rumen, as well as systemic spread to the liver, kidney, and spleen.1-3,8-10 Enteritis and encephalitis have also been reported with this viral agent.1,3 The purpose of the current study is to describe the prevalence and lesions in systemic herpesvirus disease seen in neonatal calves over a 6-year period (2004–2009), involving multiple calf ranches and large dairies served by the California Animal Health and Food Safety Laboratory in Tulare, California.
During the 6-year period, 62 BHV-1–infected neonatal calves 1–30 days of age were submitted for examination. The animals came from multiple calf-raising operations (dairies and calf ranch facilities) located primarily in the central California dairy region and Idaho. Calves were submitted to the laboratory for disease evaluation, and BHV-1 was not suspected. The majority of animals submitted for examination were calves submitted directly to the Tulare laboratory for necropsy but some submitted cases involved only tissue submissions. In both calf and tissue submission cases, the following tissues when available were placed in 10% neural buffered formalin for routine histopathology: heart, lung, liver, kidney, spleen, adrenal gland, thymus, lymph nodes, thymus, rumen, abomasum, multiple sections of small intestine including ileum, large intestine (spiral colon), and brain (brain examined histologically only if meningitis or encephalitis was suspected) . The lungs and liver were routinely cultured for common aerobic bacterial pathogens. Salmonella cultures of pooled bile and colonic content were performed on all calves. Sections of the colon were tested for the presence of coronavirus and Bovine viral diarrhea virus (BVDV) by fluorescent antibody techniques. Feces were tested for cryptosporidia and rotavirus. Fixed tissues were trimmed, processed, embedded in paraffin, sectioned at 5 µm, and stained with hematoxylin and eosin for routine histopathological examination.
All animals were examined histopathologically for pathological lesions. Sixty-two of the 2,980 calves (2.08%) had histopathological lesions consistent with BHV-1 involving 1 or more tissues. Affected calves ranged in age from 7 to 30 days of age. The mean and median age of affected calves was 14 days of age. Most cases of BHV-1 were in calves 12–15 days of age with 34 of the 62 calves (54.8%) occurring in this age group. Eleven calves (17.8%) were between 7 and 11 days of age; 14 calves (22.6%) were between 16 and 21 days of age; and 3 calves (4.8%) were between 22 and 30 days of age.
Lesions noted during histopathological examination that were suggestive of herpesvirus were confirmed as positive for BHV-1 by immunohistochemistry using a polyclonal antibody a that was specific for BHV-1. Only areas with lesions stained positive for the virus. Adrenal glands, lungs, liver, lymph node, thymus, heart, trachea, larynx, small intestines, spiral colon, esophagus, rumen, omasum, and abomasum had BHV-1 detected in 1 or more animals (Table 1).
Organs affected by Bovine herpesvirus 1 and the number of animals with lesions.

Of the 62 animals examined, 54 animals had adrenal gland lesions consistent with BHV-1. In 29 of these animals, the adrenal gland was the only organ identified as having lesions. In 25 of the animals, the adrenal gland plus 1 or more additional tissues had lesions in which BHV-1 was detected. Five animals had no adrenal gland lesions but had 1 or more additional tissues with lesions consistent with herpesvirus. In 3 animals, no adrenal gland was presented for examination.
Histologically, the lesions seen were typical of herpesvirus infection. The number of animals with involvement of each site is listed in Table 1. Lesions in the adrenal gland, liver, kidney, lymph node, and thymus were characterized by multifocal random foci of necrosis disrupting the normal parenchymal architecture. Scattered neutrophils infiltrated the associated debris and necrotic cells (Fig. 1A). Occasional lightly eosinophilic intranuclear inclusion bodies were present in the nuclei of the adrenal gland (most prevalent), liver, and kidney (occasionally seen in hepatocytes or renal epithelium). In the adrenal gland, the foci of necrosis were variable in size and ranged from discrete multifocal areas of necrosis to larger coalescing foci of necrosis in the cortex. Necrotic foci in the liver and kidney were multiple, variably sized, discrete foci of necrosis in the parenchyma. Lung lesions (Fig. 2A, 2B) consisted of necrosis of bronchial and bronchiolar epithelium with sloughing of epithelium into the lumina. Multifocal random areas of necrosis of the alveolar and bronchiolar parenchyma were evident with variable numbers of neutrophils and macrophages present in the debris. Occasional cells (primarily bronchiolar epithelium) with eosinophilic intranuclear inclusions and syncytial cells were present in the debris. A few pulmonary vessels in some sections of lungs examined demonstrated necrosis of the tunica media with variable amounts of debris present with a few inflammatory cells. Ruminal and esophageal lesions often were characterized by proliferation of the epithelium with necrosis of the surface epithelium; scattered multinucleated syncytial cells were often associated with the proliferative epithelium. Some eosinophilic intranuclear inclusions were present in associated epithelial cells and syncytial cells. Variable numbers of neutrophils were associated with the necrotic epithelial debris. In the rumen, bacteria and or yeast were often present in the hyperplastic epithelium and epithelial debris. The abomasal lesions were characterized by multifocal necrosis of the mucosa with disruption and loss of both glands and stromal connective tissue of the lamina propria. The associated muscularis mucosa was often necrotic, and the submucosa was often thickened with fibrin and edema present. Variable numbers of viable and degenerate neutrophils were present in the mucosa and submucosa. The necrotizing enteritis or enterocolitis was characterized by segmental attenuation and ulceration of the mucosa with necrosis, sloughing, and loss of epithelial cells in the intestinal gland. Sloughed necrotic epithelial cells and variable numbers of neutrophils and lymphocytes mixed with debris were present in the affected mucosa and adjacent submucosa (Fig. 3A). Necrosis (both coagulative and lytic) of the smooth muscle with variable amounts of debris and inflammation was noted in some sections of affected intestines (Fig. 4). Ganglia in the submucosa and tunica muscularis often had undergone necrosis and degeneration with lymphocytic infiltration of the ganglia and the adjacent connective tissue (Fig. 5A). Occasional intranuclear inclusions were noted in the neurons present in the ganglia. Tracheal and laryngeal lesions were characterized by necrosis of the surface epithelium with variable numbers of neutrophils in the associated cellular debris. Viable epithelium adjacent to the necrotic foci was often flattened over the affected mucosa with occasional multinucleated syncytial cells present in both viable and necrotic areas. Occasional intranuclear inclusion bodies were present in both viable epithelium and syncytial cells. The heart had several small foci of myocardial necrosis characterized by intense eosinophilia and condensation of myocardial fibers, mild edema of the interstitial connective tissue, and minimal neutrophilic infiltrates in the necrotic foci. No intranuclear inclusions were noted.
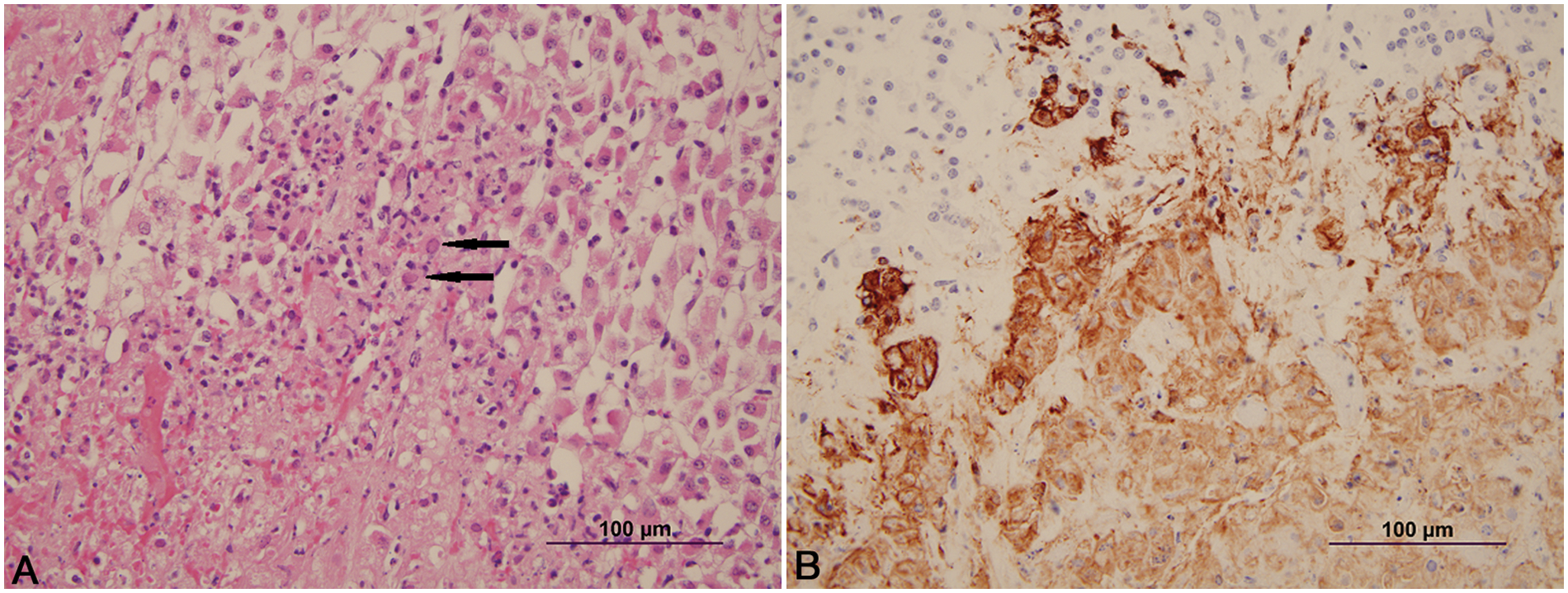
Calf; adrenal gland.
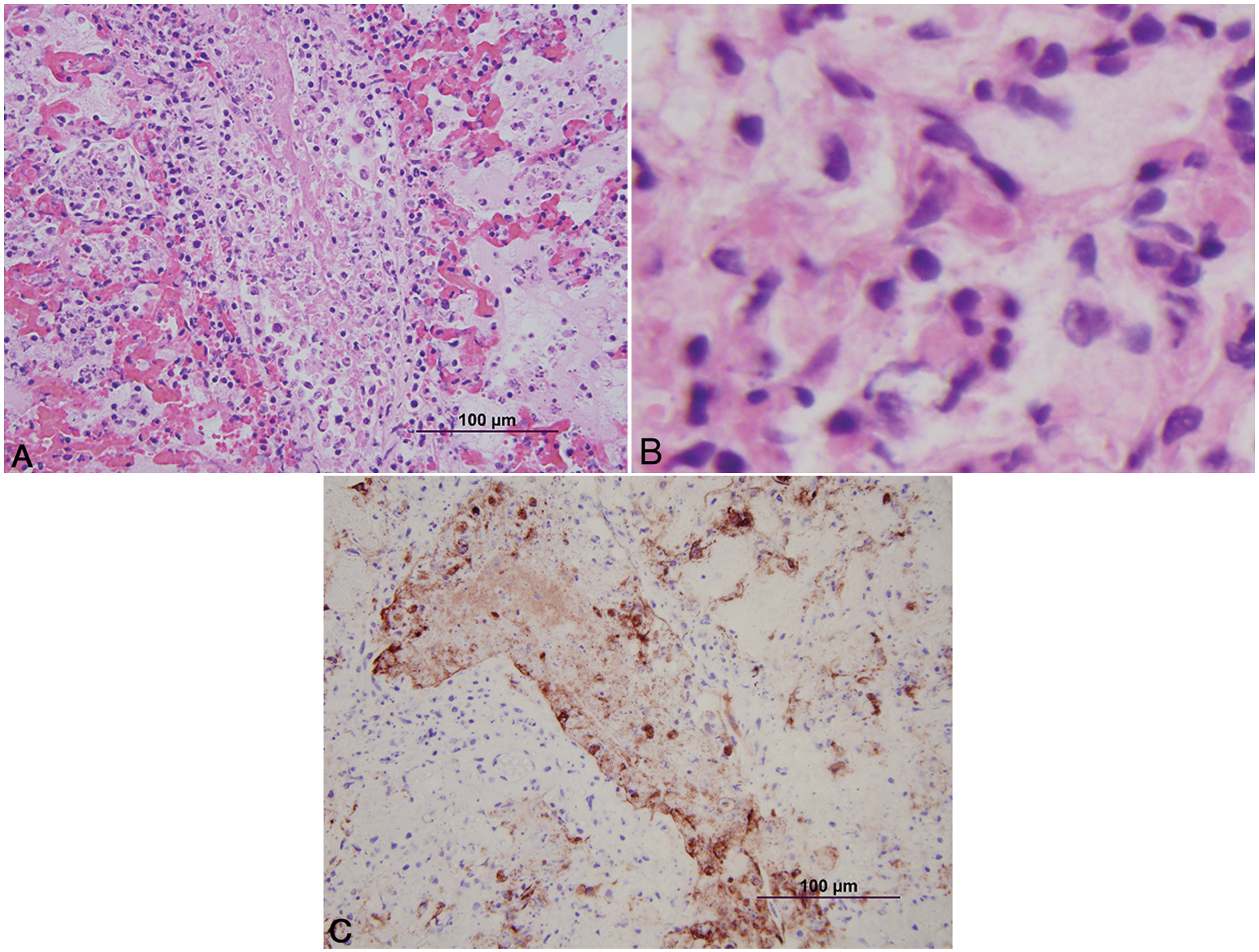
Calf; lung.
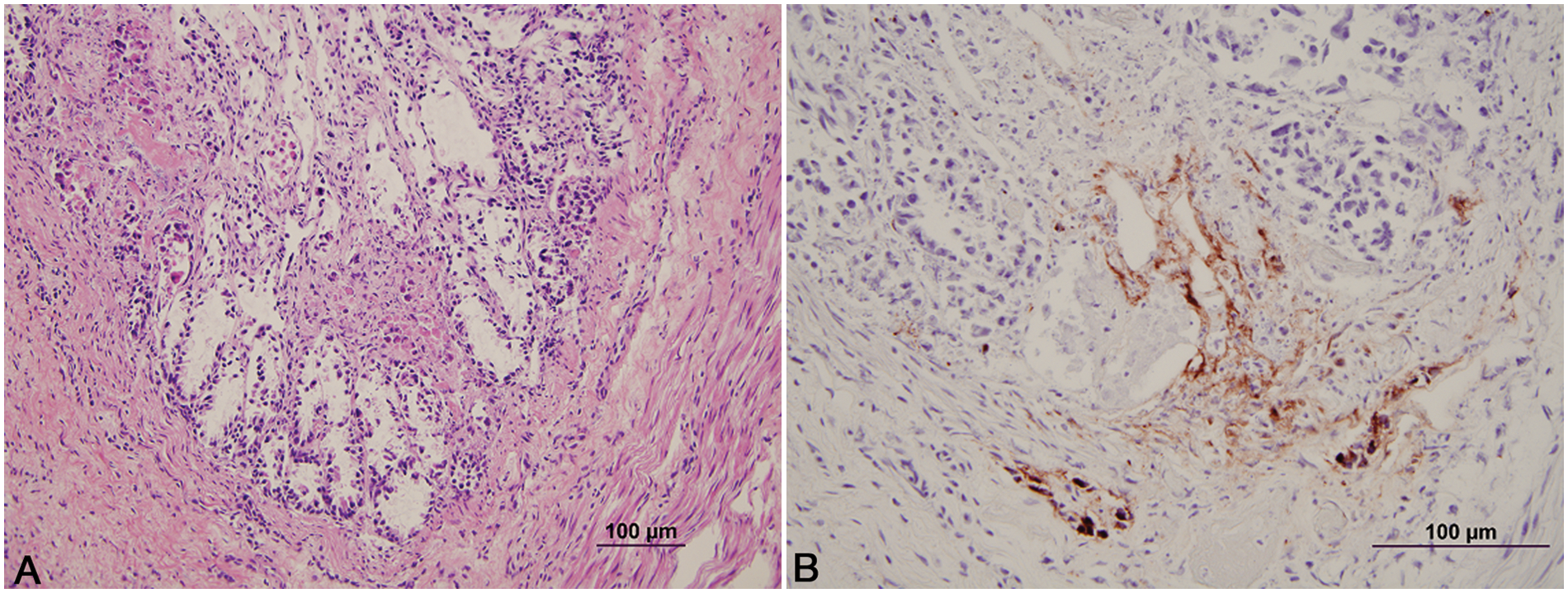
Calf; colon.
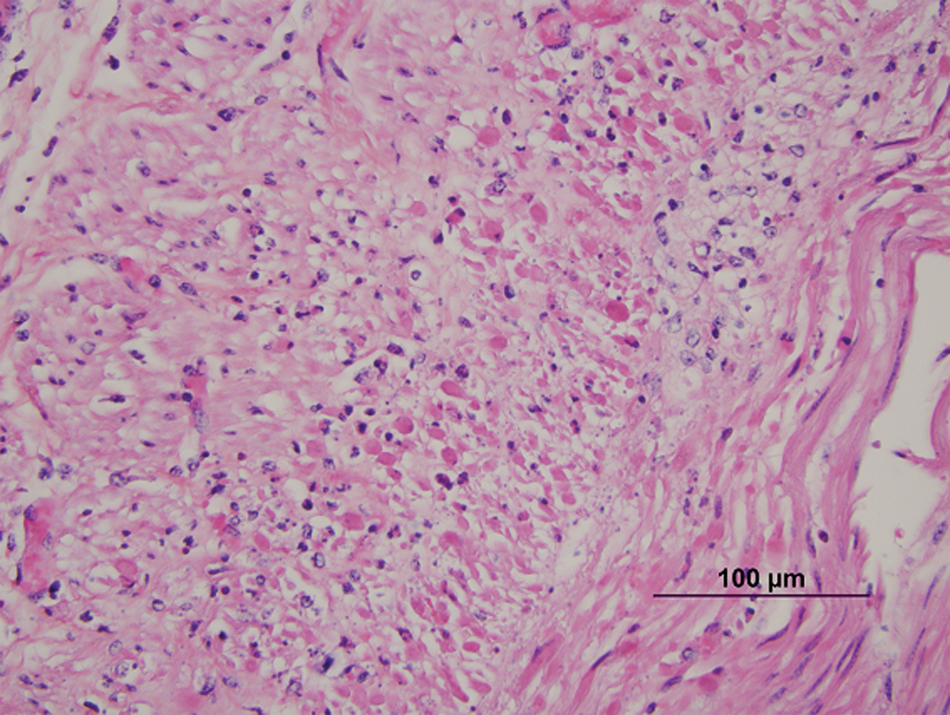
Calf; colon. Necrosis of the smooth muscle of the muscularis externa. Hematoxylin and eosin. Bar = 100 µm.
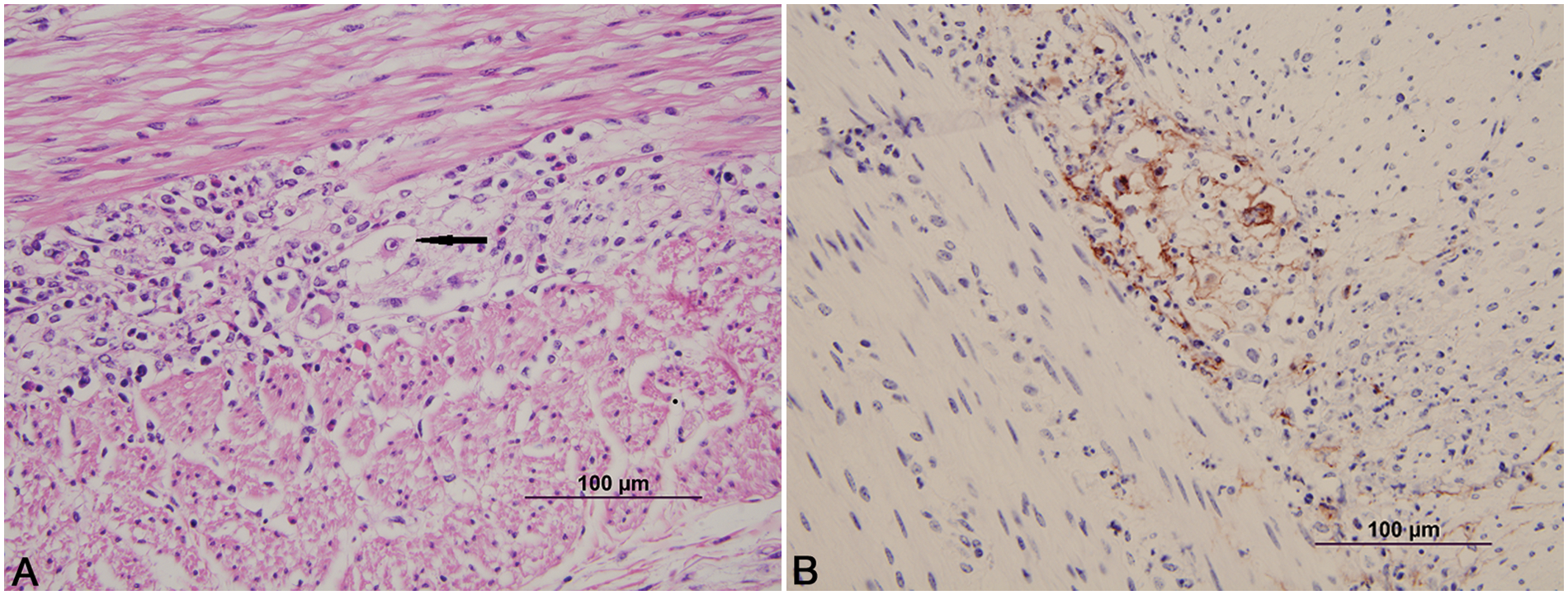
Calf; colon.
Immunohistochemistry in all cases identified viral antigen present only in the affected areas. In the adrenal gland (Fig. 1B), liver, and kidney, positive staining for BHV-1 involved the degenerate and necrotic parenchymal cells and the associated stromal connective tissue. In the lungs (Fig. 2B), positive staining involved the bronchi and bronchiolar epithelium, stromal connective tissue, and occasional vessel walls in affected regions. In the intestines, positive staining was noted involving neurons in the parasympathetic ganglia of the myenteric plexus and associated necrotic stroma of the ganglia, and affected necrotic regions of the mucosa and submucosa (Fig. 5B). Occasional vessels in the submucosa and lamina propria also demonstrated positive staining involving the vessel wall and endothelium (Fig. 3B). Intestinal epithelium and necrotic smooth muscle often failed to have positive staining for BHV-1. In the rumen and esophagus, positive staining involved primarily the affected epithelium and to a lesser extent the associated connective tissue of the adjacent submucosa.
In the 62 cases, additional agents identified in these animals could have significantly contributed to the death of the animals. In the 33 animals with BHV-1 present in multiple organs, 23 calves had 1 or more enteric pathogens (rotavirus, coronavirus, cryptosporidia, attaching and effacing E. coli, Salmonella Typhimurium, and Salmonella Anatum) in the intestines. Ten of these 23 animals had lesions of BHV-1 in the large intestines (spiral colon) and/or small intestines.
Twenty-five calves had pulmonary lesions with bacterial pathogens (Bibersteinia trehalosi, Mannheimia haemolytica, Mannheimia sp. indol +, Streptococcus suis, Mycoplasma bovis, Mycoplasma bovirhinis, Arcanobacterium pyogenes, and E. coli) often in mixed cultures isolated from the lungs. Fifteen of these 25 the calves had lesions consistent with BHV-1 in the lungs while the other 10 calves with bronchopneumonia did not have lesions in the lungs consistent with BHV-1. Seven animals had a bacterial septicemia (6 animals with E. coli and 1 animal with Klebsiella pneumoniae).
Twenty-nine of the animals had histopathological lesions compatible with BHV-1 only in the adrenal gland. It is unclear as to the significance of this lesion because all the animals had other disease problems that caused the animal’s death. Twenty-three of the 29 animals had an enteritis and/or colitis with severe dehydration. Enteric pathogens (coronavirus, cryptosporidia, rotavirus, and attaching and effacing E. coli) were seen as significant in causing the death of these animals. A bacterial septicemia (E. coli) was also present in 4 of the 29 animals. A bacterial bronchopneumonia (no evidence of BHV-1 present in the lungs by immunohistochemistry) was present in 8 of the animals and was considered to be a major contributing factor to the death of the calves. Mannheimia haemolytica, B. trehalosi, and S. suis in mixed cultures were the major pathogens present in the lungs of these animals. If the adrenal gland had not been examined on these animals, the associated herpesvirus infection would not have been identified.
It is interesting to note that in reported cases of systemic BHV-1 infections in neonatal calves, BHV-1 respiratory disease was the primary contributing factor to the calves’ illness.2,3,8,10 In the neonatal cases presented in this review, respiratory infections associated with BHV-1 infections were only noted in 15 of the 62 animals (24%). In addition to lung lesions seen in these 15 animals, lesions suggestive of multi-organ involvement were seen in the adrenal gland (13), liver (7), kidneys (2), colon (3), rumen (2), abomasum (2), esophagus (1), and larynx (1). The 10 other animals with a respiratory infection had no lesions consistent with BHV-1 in the lungs, but had other organs with BHV-1 lesions. All 10 animals had lesions in the adrenal gland while 2 animals had lesions present in additional organs (1 animal with liver, rumen, and colon lesions; 1 animal with liver and kidney lesions). This would suggest that most herpesvirus infections in neonatal calves are not associated with respiratory lesions.
Systemic infections have also been described as affecting primarily the upper digestive tract, which includes the oral mucosa, esophagus, and rumen, with lesser involvement of intestines. While similar lesions in the upper digestive tract were noted, 10 animals had lesions involving the small and large intestines. Five of these animals had lesions in both the small and large intestines. Two animals had only the small intestines involved, and 3 animals had only the large intestine lesions. In these cases, necrosis of the glandular epithelium and stromal elements of the mucosa may be noted. However, the most consistent finding of the ganglioneuritis involving the myenteric plexuses and necrosis of the smooth muscle in the tunica muscularis are findings that are different from lesions seen due to other enteric pathogens common in neonatal calves.
The source of the BHV-1 infection in the current study is unknown. Maternal infections at parturition (possibly from vulvar or vaginal infections) or late gestational systemic fetal herpesvirus infections may have been the source of infection resulting in latent infections in these young animals. Examination and follow-up of dams from affected calves for post-partum vaginal lesions was not performed. Activation of latent infections related to stress and immunosuppression are a possibility because many of the animals had multiple viral, bacterial, and protozoal infections. Stress factors have been shown to contribute to expression of herpesvirus infections in cattle.4-6 Exposure at the calf-raising facility to a BHV-1 strain from contact with infected asymptomatic calves or virally infected fomites (milk bottles or contact with virally contaminated workers or hutches) cannot be ruled out.
In the present study, 87% (54/62 animals) of the calves with BHV-1 infections were male calves. Evaluation of the data on calf submissions to the laboratory from the facilities affected identified 57% of the neonatal animals submitted for examination as male calves. With such a large number of male calves affected, this could suggest that male calves are at a greater risk of developing a BHV-1 infection. Colostrum deprivation has been shown to increase the risk of calves developing immunosuppression and systemic infections.4,5 Because many male calves have low levels of colostrum antibodies, it is probable that the male calves are predisposed to BHV-1 infections.
Vaccine-induced herpesvirus infection was also a possibility. Discussing vaccine protocols with herd veterinarians and herdsmen identified the routine use, in several of the facilities, of temperature-sensitive intranasal herpesvirus vaccines to calves upon arrival at the facility (at 1–3 days of age). These facilities later vaccinated calves intramuscularly with vaccines containing modified live virus BHV-1, Bovine viral diarrhea virus (BVDV), Bovine parainfluenza virus 3 (BPIV-3), and Bovine respiratory syncytial virus (BRSV) at 3 weeks of age. Forty-nine of the 62 animals (79% of the calves) with BHV-1 infection had been vaccinated under this protocol. One facility gave calves a single dose of a 4-way modified live vaccine with BHV-1, BVDV, BPIV-3, and BRSV at 1–3 days of age; 5 of 62 animals (8%) received this protocol. Four other facilities did not give any immunizations until the calves were 21–30 days of age when they received intramuscular immunization with a modified live vaccine containing BHV-1, BVDV, BPIV-3, and BRSV. Seven of 62 infected animals (11%) had been immunized under this protocol. One facility did not give any type of immunization to the calves at this age (1 calf affected). While it is possible that the herpesvirus present in these vaccines may have caused the lesions, the herpesvirus in the intranasal vaccines is temperature sensitive and cannot replicate at body temperature in the animal.6,7 It is possible that the vaccination with modified live BVDV may have caused the calves to become immunosuppressed, allowing latent herpesvirus expression. However, the majority of cases presented were observed before vaccination with multiple viral agents containing BVDV.
In 29 animals with BHV-1, associated lesions were observed only in the adrenal gland. The cause of death of calves that only had BHV-1 lesions in the adrenal gland was often associated with severe enteric disease (23 animals). It is possible that the adrenal gland damage may have contributed to the animal’s ability to respond to illness (corticosteroids production) and dehydration (mineralocorticoid production) during the enteric disease caused by other viral, protozoal, and bacterial pathogens. Examination of the adrenal gland on routine necropsies is critical in neonatal calves to ensure that adrenal lesions caused by BHV-1 are not missed. Further investigation on the nature of this virus is encouraged in order to understand the actions of this herpesvirus in neonatal calves.
Footnotes
a.
VMRD Inc., Pullman, WA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.