
Abstract
Astrovirus has been reported to be associated with diarrhea in pigs. The current study was conducted for the detection and molecular characterization of astroviruses in diarrheic pigs submitted to the Veterinary Diagnostic Laboratory, University of Minnesota. Intestinal contents from 269 pigs were examined by reverse transcription polymerase chain reaction (RT-PCR), and 62% were found positive for astroviruses. Of the positive samples, 20% were positive for astrovirus alone while astrovirus with rotavirus was detected in 58% of the samples. The remaining 22% revealed the presence of astrovirus along with Porcine hemagglutinating encephalomyelitis virus, Transmissible gastroenteritis virus, or Porcine circovirus-2. Sequencing the capsid gene of 56 randomly selected samples confirmed them to be Porcine astrovirus type 4 (PAstV-4) with 58–100% nucleotide identity within these viruses. Phylogenetic analysis revealed 2 possible subgroups. The results indicate that PAstV is present on swine farms in the United States and that it may be associated with diarrhea either alone or in combination with other enteric viruses. Further studies are needed to determine strain diversity among porcine astroviruses so that appropriate control strategies can be devised and implemented.
Enteric viruses belonging to the Coronaviridae, Reoviridae, Parvoviridae, Circoviridae, Picornaviridae, Caliciviridae, and Astroviridae families are associated with diarrhea and other related diseases in various mammalian and avian species.1,4,5,11,18 The family Astroviridae consists of 2 genera, Avastrovirus and Mamastrovirus, based on the host range of avian and mammalian species, respectively. Astroviruses are small, nonenveloped viruses containing a single-stranded, positive-sense RNA of 6.4–7.3 kb in size. The genome consists of untranslated regions (UTR) at both 5′- and 3′-ends. The single strand of RNA has a poly-A tail at the 3′-end but no cap at the 5′ position. There are 3 open reading frames (ORFs): ORF1a, ORF1b, and ORF2 starting from the 5′-end, along with a retrovirus-like frame shift structure between ORF1a and ORF1b. The ORF1a, ORF1b, and ORF2 code for nonstructural protease, RNA-dependent RNA polymerase, and structural protein (capsid), respectively. 10
Neonatal diarrhea in piglets places an enormous economic burden on swine farmers worldwide. Association of Porcine astrovirus (PAstV) with diarrhea in 3-week-old piglets has also been reported.2,3,14 Recently, the virus was detected in 2-month-old diarrheic piglets from a farm in China. 6 In addition, PAstVs have also been reported from healthy pigs of all age groups.8,12 Studies published in 2011 from the United States and Canada report on the presence of novel astroviruses, indicating strain diversity of these viruses in pigs.7,13 A study on 24 healthy and 12 diarrheic piglets on a high-density farm in the United States found a high diversity of astroviruses on that single farm. 13 The present study was undertaken to detect and partially characterize astroviruses in diarrheic pigs.
Intestinal contents from pigs (n = 269) submitted to the Minnesota Veterinary Diagnostic Laboratory (MVDL; St. Paul, Minnesota) for determination of the cause of diarrhea were included in this study. The samples were submitted during October 2010 to December 2010, originated from 13 different U.S. states, and included 4 samples from Mexico and 2 from Canada (Table 1). The chief complaints included preweaning diarrhea (n = 122), postweaning diarrhea at 4–12 weeks of age (n = 81), and diarrhea (n = 66) in pigs greater than 12 weeks of age. The samples were initially examined for the presence of Rotavirus A, B, and C, Transmissible gastroenteritis virus (TGEV), Porcine circovirus-2 (PCV-2), and Porcine hemagglutinating encephalomyelitis virus (PHEV) by real-time reverse transcription polymerase chain reaction (RT-PCR) using specific primers and probes for each virus. 9
Samples tested in the current study.*
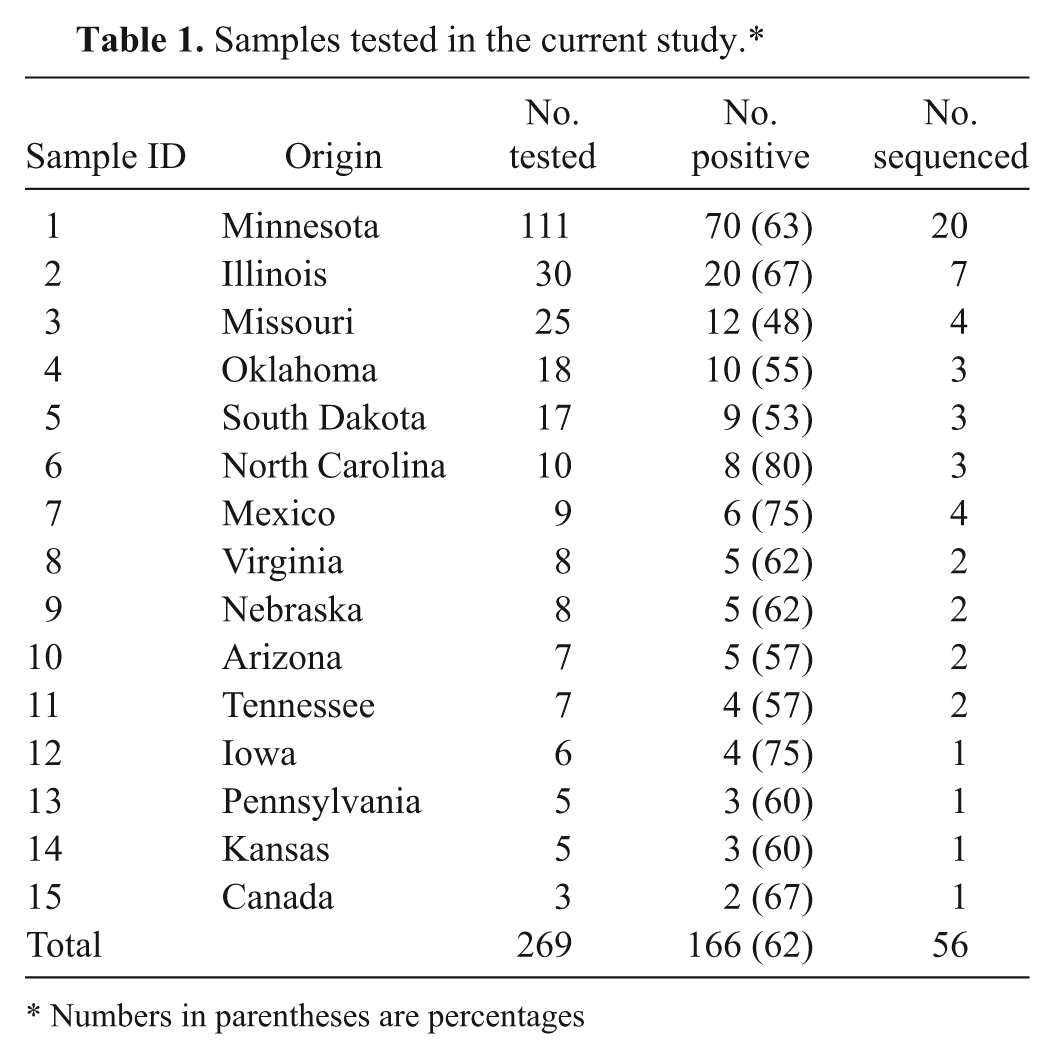
Numbers in parentheses are percentages
For the detection and molecular characterization of astroviruses, RNA was extracted from the 269 samples using a commercial RNA kit. a Extracted RNA was subjected to RT-PCR using primers specific for partial amplification of ORF1b/ORF2 (PAstV-F: 5′-TGACATTTTGTGGATTTACAG TT-3′and PAstV-R: 5′-CACCCAGGGCTGACCA-3′). 12 A commercial one-step RT-PCR kit a was used for RT-PCR, and the reaction mixture consisted of: 10 µl of 5× RT-PCR buffer, 320 µM of each deoxyribonucleotide triphosphate, 2 µl of enzyme mix, 0.6 µM of primer, 100 ng of template, and DNAase-RNAase free water to make a total volume of 50 µl. Amplification reaction consisted of 50°C for 30 min for reverse transcription, initial denaturation at 94°C for 15 min, 35 cycles of denaturation at 94°C for 30 sec, annealing at 45°C for 45 sec, extension at 72°C for 45 sec, and final extension at 72°C for 10 min. The PCR products were separated by electrophoresis on 1.2% agarose gel, and the presence of a band at the 799-bp position confirmed the presence of PAstV.
Of the 269 samples tested, 166 (62%) were found positive for PAstV. The percentage of PAstV-positive samples was 64% in preweaning pigs, 62% in postweaning pigs, and 58% in pigs of >12 weeks of age. The presence of PAstV in 62% of diarrheic pigs in the current study is high when compared to a Colombian study in which only 24% of samples collected from diarrheic piglets were astrovirus positive. 16 However, the results of the present study are similar to those of a Canadian study in which astrovirus was detected in 80% of healthy finisher pigs. 8 A 2011 study on fecal virome analysis on a high-density U.S. farm also reported the presence of astrovirus in 10 out of 12 diarrheic and 17 out of 24 healthy piglets. 13
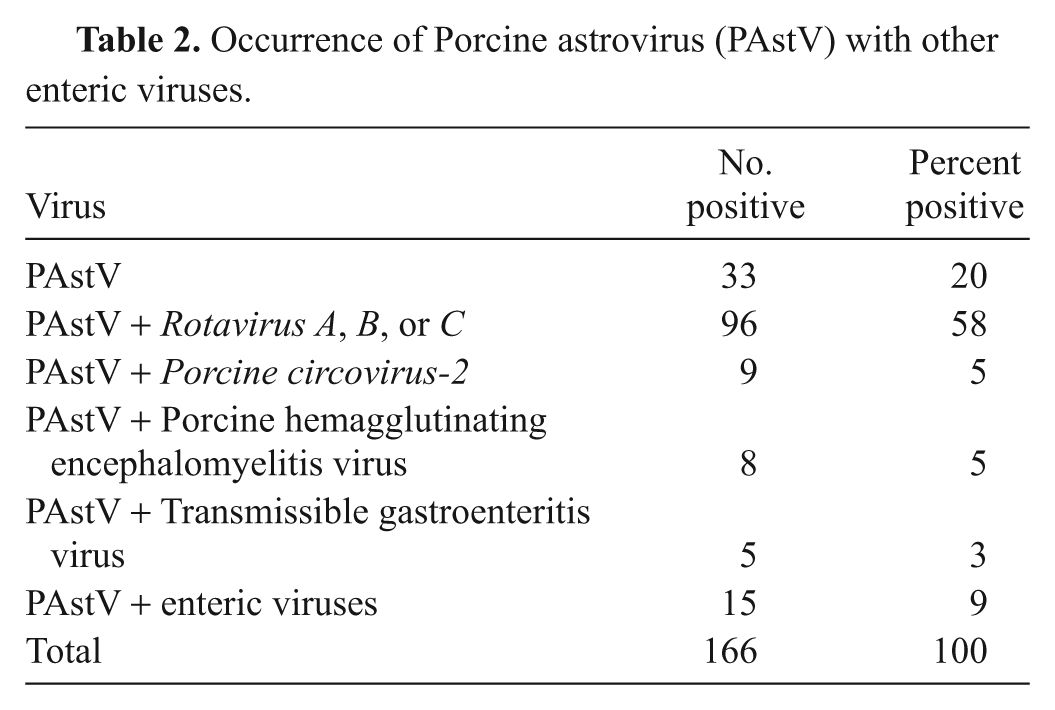
Of the 166 positive samples, 20% were positive for PAstV alone while 58% were positive for a combination of astrovirus and rotavirus. The remaining 22% showed the presence of astrovirus along with PHEV, TGEV, or PCV-2 (Table 2). These results are in line with those of a previous study from South Africa in which astrovirus-like particles were found in association with adenovirus and rotaviruses from piglets with acute diarrhea. 3 In another study, astroviruses were detected in association with calicivirus and rotavirus in feces of diarrheic piglets by electron microscopy. 2
Occurrence of Porcine astrovirus (PAstV) with other enteric viruses.
The PCR products from 56 samples representing 1–2 strains from each of the 13 U.S. states (Table 1) were purified using a commercial PCR purification kit a and sequenced at the Advanced Genomic Analysis Center (University of Minnesota, St. Paul, MN). Sequencing was conducted in both directions using the same primers as used in RT-PCR reactions. Forward and reverse sequences were aligned together using Sequencher software (http://genecodes.com/) followed by BLAST analysis (http://www.ncbi.nlm.nih.gov/blast/Blast.cgi). The nucleotide sequences thus obtained were then aligned by ClustalW method using MEGA 4.0 software. 15 The evolutionary distances were computed using the Maximum Composite Likelihood Model. A phylogenetic tree of aligned sequences was constructed by the neighbor-joining method using 1,000 bootstrap replicate values. All sequences were submitted to GenBank under accession numbers JF272545 to JF272600.
On the basis of phylogenetic analysis of ORF2, all PAstVs have been divided into 5 distinct lineages (PAstV 1–5) within the genus Mamastrovirus.7,13 The strains in the current study matched broadly with lineage 4 of PAstV but were further subdivided into 2 subgroups. The strains in one subgroup were closer to reference strain PAstV Hungary/2007 (GenBank accession no. GU562296) while those in the second subgroup were closer to PAstV 4-35 (GenBank accession no. JF713713; Fig. 1). The percent similarity among strains of subgroup 1 was 92–98% while those in subgroup 2 did not resolve in a single homogeneous group. The homology between the 2 subgroups was 57–79%, which is relatively low, indicating the possibility of antigenic variation among PAstVs as has been reported in several other studies.8,13,16 Human astrovirus strains having less than 95% homology at the nucleotide level have been shown to be serologically distinguishable. 17 It was not possible to ascertain if the 2 subgroups observed in the present study represent separate serotypes. Complete gene sequencing will help answer this question.

Phylogenetic tree of nucleotide sequences for the capsid gene of Porcine astrovirus (PAstV). Sequences starting with PAstV 1101-1156 are from the present study, and the GenBank accession numbers are from previously published sequences. Phylogenetic tree was constructed by the neighbor-joining method using p-distance substitution model with 1,000 bootstrap replicates and a cut-off value of 70% by using MEGA 4.0.14
The PAstV sequences from Canada and China (GenBank accession no. HM756260, HM756261 from Canada, and HQ647383 from China) were also used for comparison. The similarity with these strains was found to be only 39–49%, while similarity with human astroviruses was only 41–49% (Fig. 1). Sequencing of ORF2 revealed that the Canadian and Mexican strains in the current study were related to the U.S. strains. The nucleotide similarities of these strains with the U.S. strains were 74–99.7% and 74–100%, respectively.
As has been previously reported,7,8,12 a conserved region, UUUGGAGGGGCGGACCN AA N5-8 AUGNC (where N stands for any of 4 nucleotides), was observed at the overlapping region of ORF1b and ORF2 just before the start codon AUG in all PAstV sequences of the present study. Alignment of this conserved region showed a unique insertion of 3 nucleotides (CTA) upstream of the ORF2 start codon in all sequences of this study. This highly conserved region is supposedly the longest conserved region in PAstVs as compared to other astroviruses 12 and has been proposed to act as a regulatory element serving as a promoter for subgenomic RNA transcription. 10 The finding of this unique insertion of 3 nucleotides upstream of the ORF2 start codon is in line with previous studies,7,8 but its importance has yet to be determined.
Two studies published in 2011 confirmed the presence of multiple lineages of PAstVs with each lineage, likely representing an independent origin.7,13 On correlating phylogenetic analysis of partial capsid gene sequence with clinical history, it was observed that group I strains were from apparently healthy pigs while group III strains were detected in suckling piglets. 8 Age restricted pattern of infection for these strains was suspected. In fact, a fecal virome analysis showed that 19-day-old piglets had on average of 1.5 viral coinfections as compared to 5.2 in older animals. 13 Using a chi square test, a relationship between age and the presence of virus was not observed. In addition, no grouping on the basis of different states or region was found.
Further studies are needed to determine the exact role of astrovirus, whether alone or in combination with other enteric viruses, in causing diarrhea of pigs. Studies are also needed to determine why astroviruses are found in both apparently healthy pigs and diarrheic pigs. Characterization of viruses by complete gene sequence of polymerase and capsid genes will help obtain a clearer picture of viral serotypes circulating in swine farms.
Footnotes
a.
Qiagen Inc., Valencia, CA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.