
Abstract
The histological lesions caused by Edwardsiella tarda in a variety of fish species, including tilapia, have been well characterized. There are apparent differences in the type of inflammatory response manifested by these different species, which may be due to the fish species itself, the phase of infection, or the virulence factors produced by different strains of E. tarda. In catfish, systemic abscesses involving muscles of the flank or caudal peduncle are the most common lesions. By contrast, infection in tilapia and red sea bream is more likely to be associated with granulomatous inflammation. Necrotic meningitis, encephalitis, and vasculitis with fibrinoid necrosis of the blood vessels walls, as well as the formation of a plaque-like structure in the brain, are described in the current study. The presence of E. tarda was confirmed by microbiological isolation and a positive nested polymerase chain reaction in paraffin wax–embedded tilapia tissues.
Edwardsiella tarda is a Gram-negative, motile, short, rod-shaped bacterium (1 µm × 2–3 µm) of the family Enterobacteriaceae. It is well known for causing a severe disease in freshwater and marine fish in both farmed and wild populations. It was initially found to cause disease in farmed channel catfish (Ictalurus punctatus) in the United States. 8 In Japan, the bacteria were isolated from diseased eels (Anguilla japonica) and subsequently named Paracolobactrum anguillimortiferum. 2 Currently, the disease known as Edwardsiella septicemia occurs with high frequency in tropical fish species, which are common intestinal carriers, and thereby results in possible contamination of the carcasses during fish processing.
The clinical, gross, and microscopic changes caused by E. tarda have been relatively well characterized in a range of different fish species; in catfish, the changes comprise the formation of abscesses within muscles of the flank or caudal peduncle. The lesions rapidly increase in size and develop as large cavities filled with gas. When the lesions are incised, a foul odor is emitted.5,8 Other gross lesions in E. tarda infection include eye tumefaction, cloudy and opaque corneas, and hemorrhage at the base of the dorsal and anal fins. Internally, generalized hyperemia, hemorrhages, swelling, and necrosis of organs such as liver, spleen, kidney, gastrointestinal tract, and musculature are also present. Microscopic lesions present as abscesses disseminated systemically.1,9,10,12
Individual fish species may manifest differences in the type of inflammatory response against E. tarda. The lesions in Japanese eel and Japanese flounder were described as a kind of suppurative inflammation, whereas the infection in tilapia and red sea bream was documented as a more granulomatous inflammation preceded by macrophage infiltration and a reduced number of bacteria inside the granulomas. 9
In Colombia, the production of tilapia has increased significantly (Food and Agriculture Organization of the United Nations: 2003, National aquaculture sector overview: Colombia. Available at http://www.fao.org/fishery/countrysector/naso_colombia/en. Accessed on December 2, 2011), and there has been a parallel rise in the incidence of bacterial diseases such as those caused by E. tarda. Edwardsiella tarda infection in red tilapia (Oreochromis spp.) cultured in Colombia is characterized by clinical, gross, and microscopic changes similar to those described for this fish species elsewhere4,9; nonetheless, during a routine diagnostic exercise in the Veterinary Pathology Laboratory at the National University of Colombia (Bogotá, Distrito Capital, Colombia), novel histological lesions in the vessels of the brain of a red tilapia were associated with infection with E. tarda.
Five red tilapia of approximately 40–50 g body weight were received for diagnostic evaluation because of heavy mortalities on a commercial farm. On arrival at the laboratory, the fish were anesthetized with MS 222 (tricaine methanesulfonate), the external surface was sterilized, and on opening the abdominal cavity, samples for microbiology were immediately taken and cultured in brain–heart infusion agar enriched with 5% defibrinated ovine blood and incubated at 28°C for 36 hr. Tissue samples for histopathology included gastrointestinal tract, liver, kidney, spleen, heart, gills, eyes, and brain. The samples were immersed in 3.7% buffered formalin and were processed by routine histological technique of paraffin wax embedding and hematoxylin and eosin staining. Finally, a nested polymerase chain reaction (nPCR) technique of the same tissues embedded in paraffin wax was implemented.
Gross lesions included corneal opacity and the loss of one eye in one of the submitted fish. Reddening of the anal papilla and marked pallor of the gills were also seen. Internally, kidney and liver were pale and seeded with white 1 × 1–mm nodules. Flocculent material was present in the swim bladder and kidney. Congestion and hemorrhage on the intestinal serosa were pronounced. One animal had an abscess at the base of the pelvic fin as well as one in the brain. All of these findings were strongly suggestive of infection by E. tarda.
The lesions observed grossly in the liver, kidney, spleen, pancreas, peritoneum, and peritoneal fat corresponded microscopically to pyogranulomatous inflammation. Severe gastritis and necrotic and hemorrhagic enteritis were also found with abundant mucus production. Bacteria accompanied by inflammation were present in the gills. The inflammatory reaction also involved the eyes and periorbital tissues. A granuloma was observed in the skeletal muscle.
Microscopically, only 1 fish was remarkable due to the histological findings in the brain and is the subject of the current report. The lesions consisted of inflammatory, necrotic, and proliferative changes affecting the walls of the blood vessels of the meninges and brain parenchyma. Inflammatory responses in the brain were mainly represented by meningitis, with fibrinoid necrosis of vessel walls. This was accompanied by an abundant influx of mononuclear leukocytes and fewer neutrophils, some of which were necrotic, characterized by nuclear pyknosis and cytoplasmic eosinophilia (Fig. 1). Vessels in the brain parenchyma had similar changes (Fig. 2). The most conspicuous change, however, was the formation of a plaque-like structure within the optic lobe. This appeared as a compact nest or “entanglement” of blood vessels in which some were seen as spreading out of or converging on the “plaque.” Hyperplasic, tightly packed muscle cells of the vessel’s walls were the main component of the plaque; some fibrinoid necrosis was still visible as well as some necrotic inflammatory cells. Where the muscle cells were cross-sectioned, they appeared as rounded cells with a slightly basophilic staining cytoplasm. Most of the nuclei of the cells were round, and less frequently ovoid, whereas the nucleoli were prominent (Fig. 3).
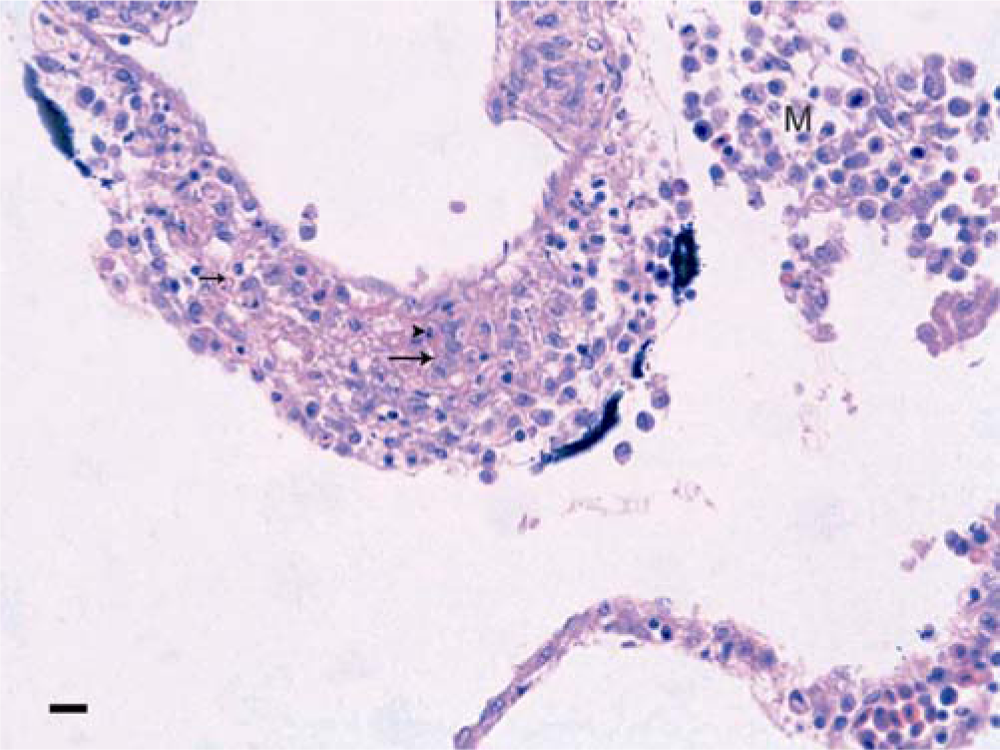
Red tilapia; meninges. Eosinophilic material (arrows), most probably fibrin, exudes between the muscle cells of the vessel wall; necrotic cells, mostly leukocytes, show karyorrhectic nuclei and deeply eosinophilic cytoplasm (arrowhead). M = mononuclear meningitis. Hematoxylin and eosin. Bar = 50 µm.
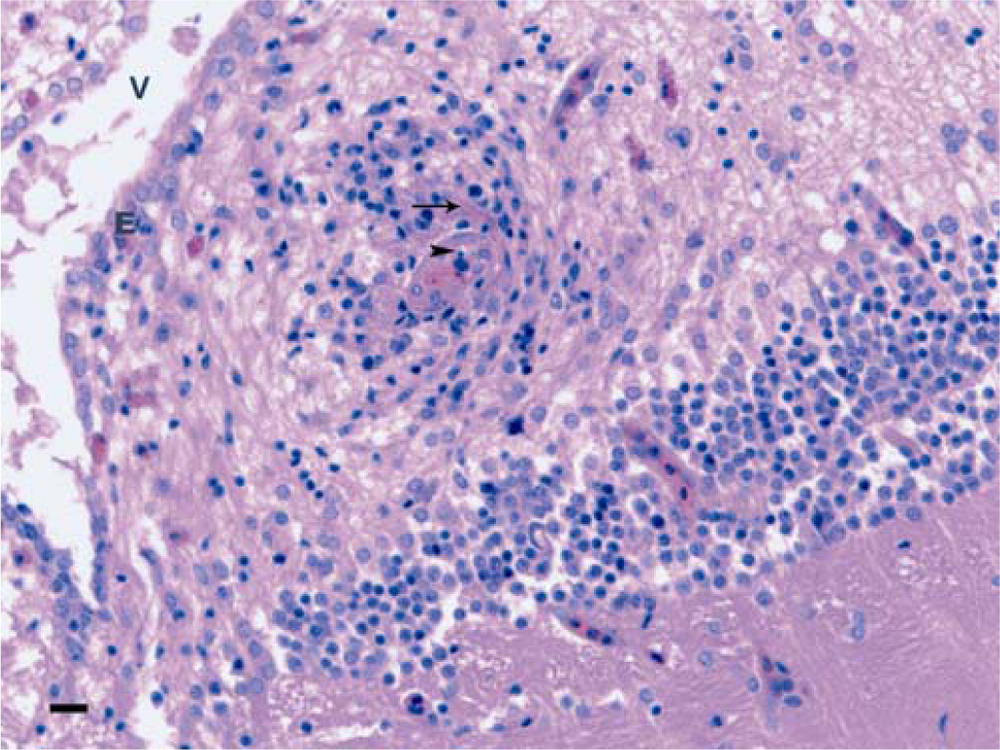
Red tilapia; mononuclear encephalitis with vasculitis. An eosinophilic exudate is visible in the upper part of the vessel (arrow), probably fibrin. Remnants of nuclei, indicating necrosis probably of leukocytes, are in the lumen of the vessel (arrowhead). V = ventricle; E = ependyma. Bar = 50 µm.
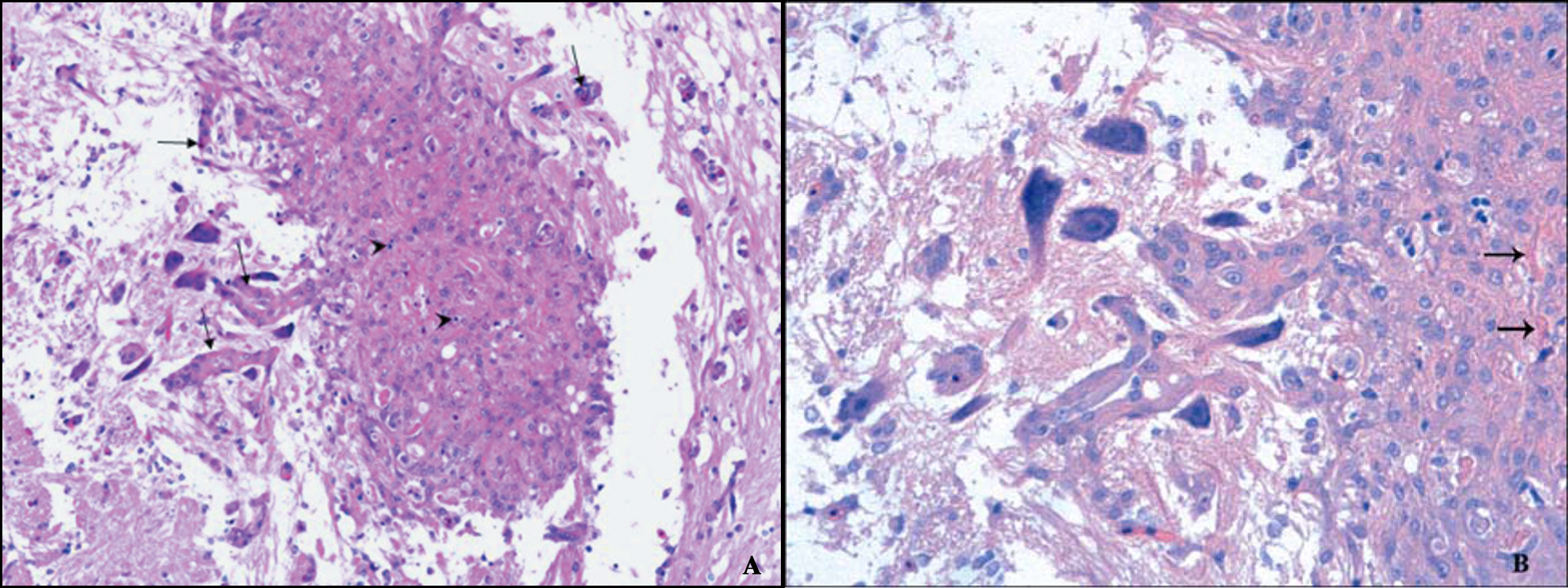
Red tilapia.
Microbiology allowed the isolation of E. tarda from this fish as well as a positive nPCR for E. tarda from the paraffin wax–embedded tissue (Fig. 4). To confirm the presence of E. tarda in tilapia tissue, genomic DNA was extracted from the paraffin wax–embedded organs, including brain, spleen, kidney, and liver, as well as from a negative control. Briefly, 10 sections (10-µm thick) were deposited in 2-ml vials, deparaffinized with xylol, and washed with absolute ethanol and 70% ethanol. Pellets were diluted with 200 µl of 1× Tris–ethylenediamine tetra-acetic acid (TE) buffer and were digested with 20 mg/ml of proteinase K a at 60°C overnight. Proteinase K was denatured by boiling at 90°C for 10 min, then a 67% solution of guanidine thiocyanate a was added, plus 1.25 mg/ml of glycogen. b Samples were vortexed and incubated at 60°C for 10 min. DNA was precipitated with 500 µl of isopropanol, mixed by vortex, and centrifuged to 12,000 × g for 15 min. Pellets were washed with 70% ethanol, centrifuged, dried, and diluted in 200 µl of 1× TE buffer.
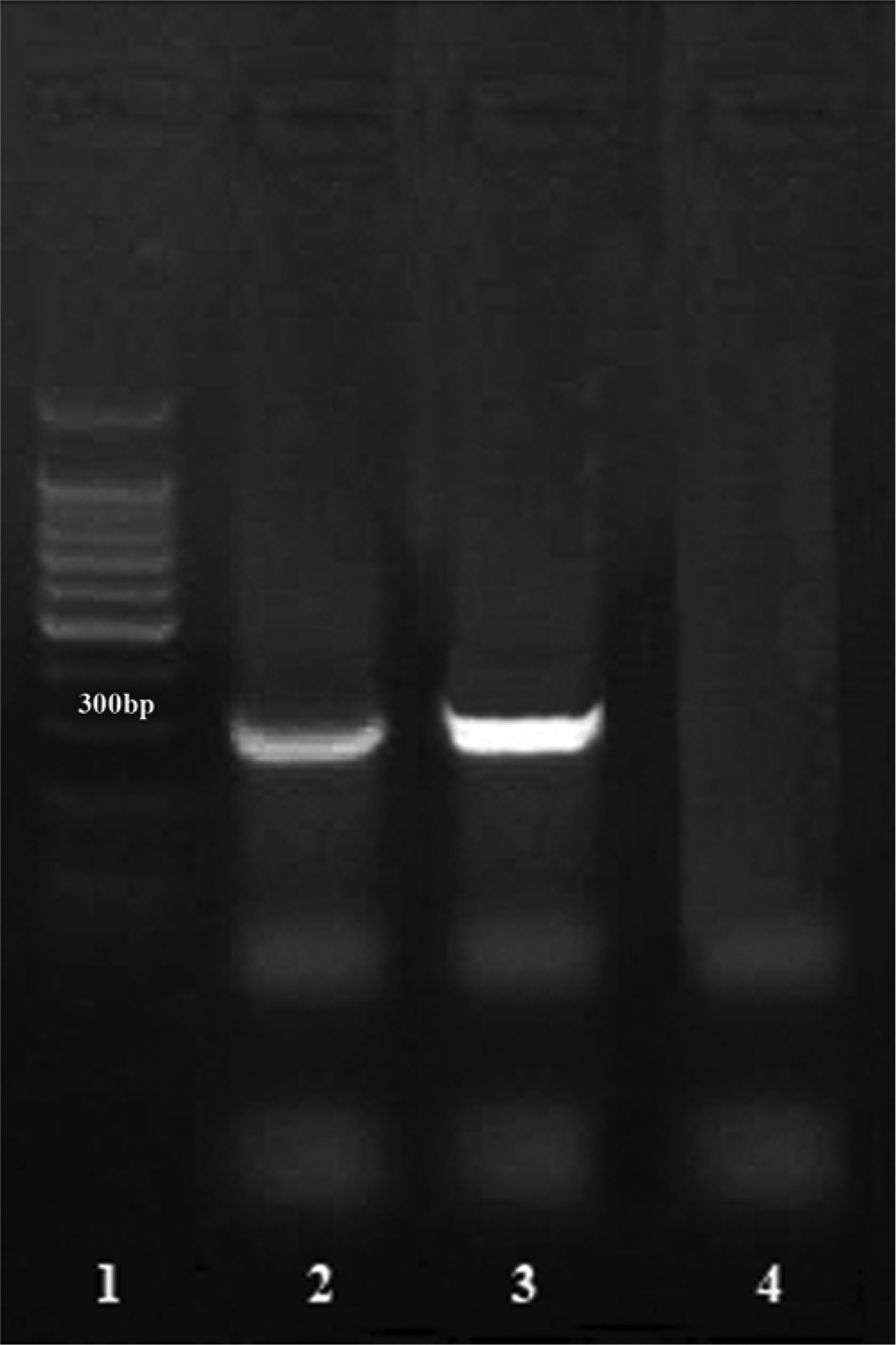
Red tilapia. Positive nested polymerase chain reaction (nPCR) for Edwardsiella tarda from the paraffin wax–embedded tissue. Lane 1: DNA ladder 100–1,000 bp; lane 2: case positive nPCR (305 bp); lane 3: positive control; lane 4: negative control.
The PCR was performed as previously described, 6 with modification. Because of the origin of the DNA sample, it was necessary to increase the sensitivity of the technique by developing an nPCR with a 305-bp fragment of the E. tarda GyrB gene. An external round of PCR with primers gyrBF1 (5′-GCATGGAGACCTTCAGCAAT-3′) and gyrBR1 (5′-GCGGAGATTTTGCTCTTCTT-3′) and an internal round of PCR with gyrBintF (5′-GCGCCTGCG CGAACTCTCTT-3′) and gyrBintR (5′-GGGTCATCGCC GCACGGAAA-3′) were amplified. The external round of PCR mixture consisted of 3 µl of DNA sample, 5 µl of 10× PCR buffer, 3 µl of 25 mM MgCl2, 1 µl of 25 mM deoxyribonucleotide triphosphate (dNTP), 0.25 µl of Taq polymerase, c and 36.5 µl of distilled water. The amplification conditions were 1 step at 94°C for 5 min, 30 cycles at 94°C for 1 min, 51.5°C for 30 sec, and 72°C for 30 sec, and a final extension step at 72°C for 5 min in a thermocycler. d The internal round of PCR mixture consisted of 3 µl of DNA sample, 1× PCR buffer, 25 mM MgCl2, 0.2 mM dNTP, 0.04 U/µl of Taq polymerase, c and 25 µl of distilled water. The amplification conditions were 1 step at 94°C for 3 min, 30 cycles at 94°C for 30 sec, 63°C for 30 sec, and 72°C for 30 sec, as well as a final extension step at 72°C for 3 min in a thermocycler. d Five microliters of each PCR product was subjected to electrophoresis in a 1% (w/v) agarose gel. a
Most of the lesions in the outbreak of edwardsiellosis in tilapia were typical of those already described in this species. In addition to fibrino-necrotic vasculitis, however, lesions in the brain of 1 fish comprised an apparently proliferative reaction of the vessel’s muscle cells. That the lesions were most probably caused by E. tarda could be deduced from the positive microbiological results and from the nPCR technique from the same tissues embedded in paraffin wax and processed for routine hematoxylin and eosin staining.
Vasculitis with exudation of plasma proteins, including fibrin, and necrosis of leukocytes are relatively well-known responses to different bacterial infections of mammals, including human beings. Some examples include Actinobacillus pleuropneumoniae, Actinobacillus suis, and Streptococcus suis in pigs and Histophilus somni in bovids. 7 Although less well documented in fish pathology, extensive necrosis of media and endothelium with fibrin exudation or thrombosis are also described in fish affected with Renibacterium salmoninarum. Vasculitis in fish is also caused by virus and mycotic invasion of vessels. 11 In human beings, 2 main pathogenic mechanisms are thought to be responsible for vasculitis: direct damage of vessels by infectious agents and immune-mediated mechanisms (immune complex, anti–neutrophil cytoplasmic antibodies, and anti–endothelial cell antibodies). This is likely the case in animals, including fish. 3 It is unclear which of the 2 mechanisms was operating in the reported lesions of tilapia, but an immune complex–mediated vasculitis induced by E. tarda would be the most plausible explanation. The proliferative character of the vascular muscle lesions in tilapia points to a chronic response, which indeed is the most frequent course (i.e., pyogranuloma formation) of the disease in this fish species; this makes it highly probable that antibodies recognizing the bacterium or some of its products are being deposited in the vessel’s walls. 7 However, no explanation could be found for lesions similar to the plaque-like formation found in the brain parenchyma of the tilapia described in the current study.
Footnotes
a.
Promega Corp., Madison, WI.
b.
Fermentas, Inc., Glen Burnie, MD.
c.
Tucantaq Corpogen, Bogotá, DC, Colombia.
d.
PxE 0.2, Thermo Fisher Scientific Inc., Wilmington, DE.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This report was supported by a grant from the Welfare Department of National University of Colombia.