
Abstract
Francisella asiatica is a Gram-negative, pleomorphic, facultative intracellular, bacterial pathogen that causes acute to chronic disease in a wide variety of warm-water cultured and wild fish species. Outbreaks of francisellosis in warm water fish have been documented in Taiwan, Japan, United Kingdom, Hawaii, and Latin America (including Costa Rica) but the organism has only been reported from the United States on one occasion from hybrid striped bass in California. In 2010, the bacterium was detected from diseased tilapia by culture on cystine heart agar supplemented with hemoglobin and by utilizing an F. asiatica–specific real-time polymerase chain reaction assay. The tilapia (Oreochromis niloticus) were cultured in an indoor, closed, recirculating aquaculture facility in the Midwest of the United States. The identity of isolates recovered from diseased fish was confirmed as F. asiatica by amplification and sequence comparison of the 16S ribosomal RNA and intracellular growth locus C (iglC) gene. Gross and microscopic examination of affected tissues revealed the presence of marked anterior renomegaly and splenomegaly with severe granulomatous disease.
Keywords
Tilapia is a generic term used to designate members of 3 different genera (Sarotherodon, Oreochromis, and Tilapia) that are commercially important food fish belonging to the family Cichlidae. 1,2,6 Due to their hardiness, fecundity, feed efficiency, and rapid growth, tilapia and their hybrids are among the most important group of farm-raised fish in the world. Moreover, the United States is the second biggest consumer of tilapia with total demand of 50 million kg in 1998. 1,2,6 In the United States, tilapia farming typically occurs in high-intensity recirculating systems that use greenhouses or industrial waste heat to maintain optimum growing conditions. 6
Francisella asiatica (syn. F. noatunensis subsp. orientalis) and F. noatunensis (syn. F. noatunensis subsp. noatunensis, F. piscicida, and F. philomiragia subsp. noatunensis) are Gram-negative facultative intracellular bacteria and the causative agents of francisellosis in fish. 14,16 These bacteria are emergent pathogens of tilapia, hybrid striped bass, salmonids, three line grunt, and many other important cultured fish species. 3,7,8,10,13,15-17 In previous work, it was found that as few as 23 F. asiatica bacteria injected in the peritoneum were capable of causing mortality in tilapia, and that even fewer were enough to cause significant pathological lesions in the head kidney and spleen. 19
Beginning in April 2010, an indoor tilapia aquaculture farm in the Midwest United States experienced increased morbidity and mortality in cultured tilapia fingerlings (10–100 g). Historically the farm had experienced problems with streptococcosis and vibriosis and initiated a vaccination program in late 2009 and early 2010 using autogenous vaccines to prevent the diseases caused by Streptococcus iniae and Vibrio vulnificus. Approximately 200 tilapia (Oreochromis niloticus) cultured in this facility were received and analyzed in the Louisiana Aquatic Diagnostic Laboratory (LADL) of the Louisiana State University, School of Veterinary Medicine (Baton Rouge, LA) in 2010. Following complete necropsy, moderate parasitic infestation with Dactylogyrus sp., Trichodina sp., and epitheliocystis were detected. Consistent gross and microscopic lesions similar to those reported during high mortality events in cultured tilapia in Costa Rica, Taiwan, United Kingdom, and Hawaii since 1994 5,7,9,11,12,20 were also noted. Grossly, fish had marked anterior renomegaly and splenomegaly (on average 10 times normal). Numerous pale tan foci (granulomas) were scattered throughout the head kidney and spleen, with lesser numbers in the trunk kidney. Affected spleens contained numerous, up to 2 microns wide, cystic, fluid-filled cavities randomly distributed throughout the tissue. The fluid within the cystic cavities was clear, tan, and serous. Often splenic tissues were effaced such that they had a pale tan homogeneously spongy consistency with abundant fibrinous adhesions. White, opaque, oily material (adjuvant) was variably present in the coelomic cavities, in some cases encased in a thin fibrous tunic. Mesenteries were variably and irregularly thickened, tan, and fibrous with multiple fine nodules (granulomas) and adhesions to viscera. Gills were pale pink and mildly thickened.
Freshly collected fish tissues were fixed, processed, and examined by light microscopy following standard histological methods. 20 Microscopically, head kidneys were markedly enlarged with up to 90% parenchymal replacement by coalescing granulomatous inflammation (Fig. 1A). Granulomas often had necrotic cores (Fig. 1B) that contained numerous 1–2 micron wide coccoid bacteria. Lesser numbers of bacteria were variably present in surrounding layers of epithelioid macrophages and in peripheral macrophages. Inflammation spared interrenal tissues but extended into associated spinal nerves. Spleens were enlarged and similarly effaced by granulomatous inflammation. Irregular ectatic endothelium lined vessels containing few cells (mostly macrophages) and proteinaceous fluid were admixed with areas of necrosis and cavitation (Fig. 1C). Splenic granulomatous inflammation was less discrete than in head kidneys, with abundant bacteria in macrophages throughout (Fig. 1D). Multifocally throughout affected tissues, necrotizing vasculitis with thrombosis was present. In many cases, vasculitis was associated with adjacent granulomatous inflammation. Livers were less affected, with small foci of necrosis and bacteria laden macrophages. Granulomatous inflammation in trunk kidneys was relatively mild, largely sparing nephrons. Gills had moderate epithelial hyperplasia with few small granulomas and isolated macrophages filled with bacteria; epitheliocystis-infected cells and monogeneans were also present. Multifocally, granulomatous inflammation caused mild thickening of the lamina propria of the intestines with few bacteria present. Atrial endothelial hypertrophy was seen in the hearts. Multifocally, there was fibrosis and lipocyte atrophy of the mesentery with numerous small granulomas that contained nonstaining cores (presumed adjuvant reaction from autogenous vaccine). Gonads, eyes, and brains were not examined.
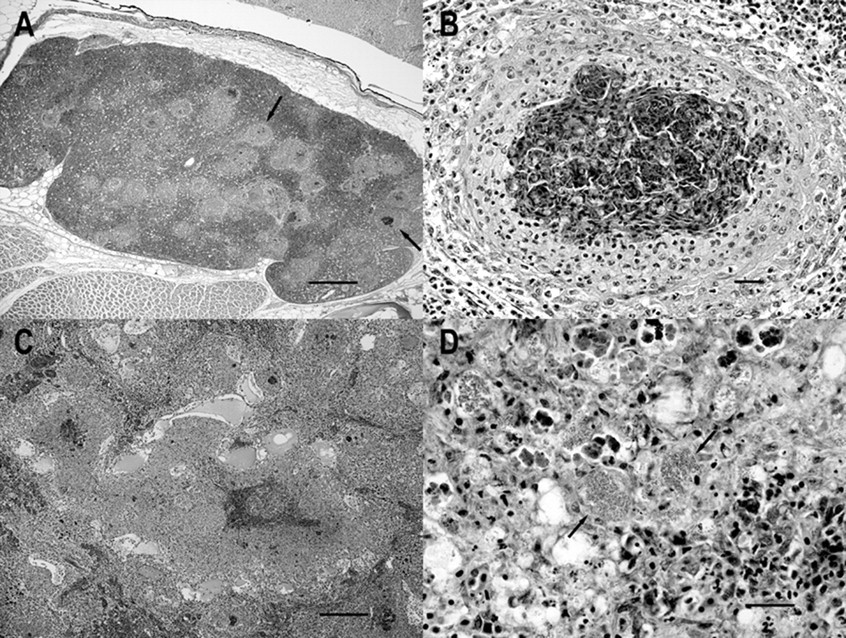
Photomicrographs of Nile tilapia (Oreochromis niloticus) infected with Francisella asiatica.
The spleen and anterior kidney from freshly euthanized fish were aseptically collected and used for bacteriological analysis by homogenizing approximately 100 mg of tissue in sterile water, and streaking 20 µl of the suspension on trypticase soy agar with 5% sheep blood (TSA) a and cystine heart agar supplemented with bovine hemoglobin solution (CHAH). b Plates were incubated at 28°C for 2–5 days. Colonies observed from primary isolation agar plates were re-plated for purity of culture under the same conditions. Once single colonies were observed and purity of the isolate determined, the isolate was re-suspended in liquid medium that consisted of a modified Mueller–Hinton II cation adjusted broth supplemented with 2% IsoVitaleX and 0.1% glucose (MMH). c,20
Three to 5 days post-inoculation of the CHAH-inoculated plates, gray, smooth, and convex colonies were observed. Similar descriptions have been previously made of F. asiatica colonies in agar media. 7,10,20 The F. asiatica failed to grow on TSA agar with 5% sheep blood; however, other primary, secondary, and contaminating organisms, including S. iniae, V. vulnificus, Vibrio mimicus, and Aeromonas spp. were isolated occasionally.
Extracted DNA from splenic tissue (approximately 50 mg) from diseased fish (n = 20) was initially used as a template in a previously described TaqMan probe real-time polymerase chain reaction (PCR) assay, specific for the detection of F. asiatica from fish tissue. 18 The DNA was extracted following the manufacturer’s protocol. d One hundred percent of the analyzed fish spleens were found to be positive for F. asiatica in the assay.
Isolates (10-051, 10-075-#1, 10-075-#5, and 10-075#8) recovered from fish cultured in different systems from the same farm and submitted to the LADL during different occasions were subjected to molecular analysis. A loop of each isolate was suspended in 400 µl of sterile water, washed, centrifuged at 3,000 × g for 5 min, and then re-suspended in 200 µl of phosphate buffered saline. The bacterial suspension was subjected to DNA extraction and purification as per the manufacturer’s protocol. d DNA was stored at 4°C until further use. Two different sets of primers were used to amplify and sequence the partial 16S ribosomal RNA (rRNA) and partial intracellular growing loci (iglC) gene sequences following previous published protocols. 19,20
The partial 16S rRNA and iglC sequences obtained were compared with those stored in GenBank using the BLASTN program. The ClustalW program was used to create a neighbor-joining tree based on the partial sequences of the genes (Figs. 2, 3). 5,17,21,22 As expected, the isolates recovered from cultured tilapia in the present study showed >99% identity to other F. asiatica sequences stored in GenBank.
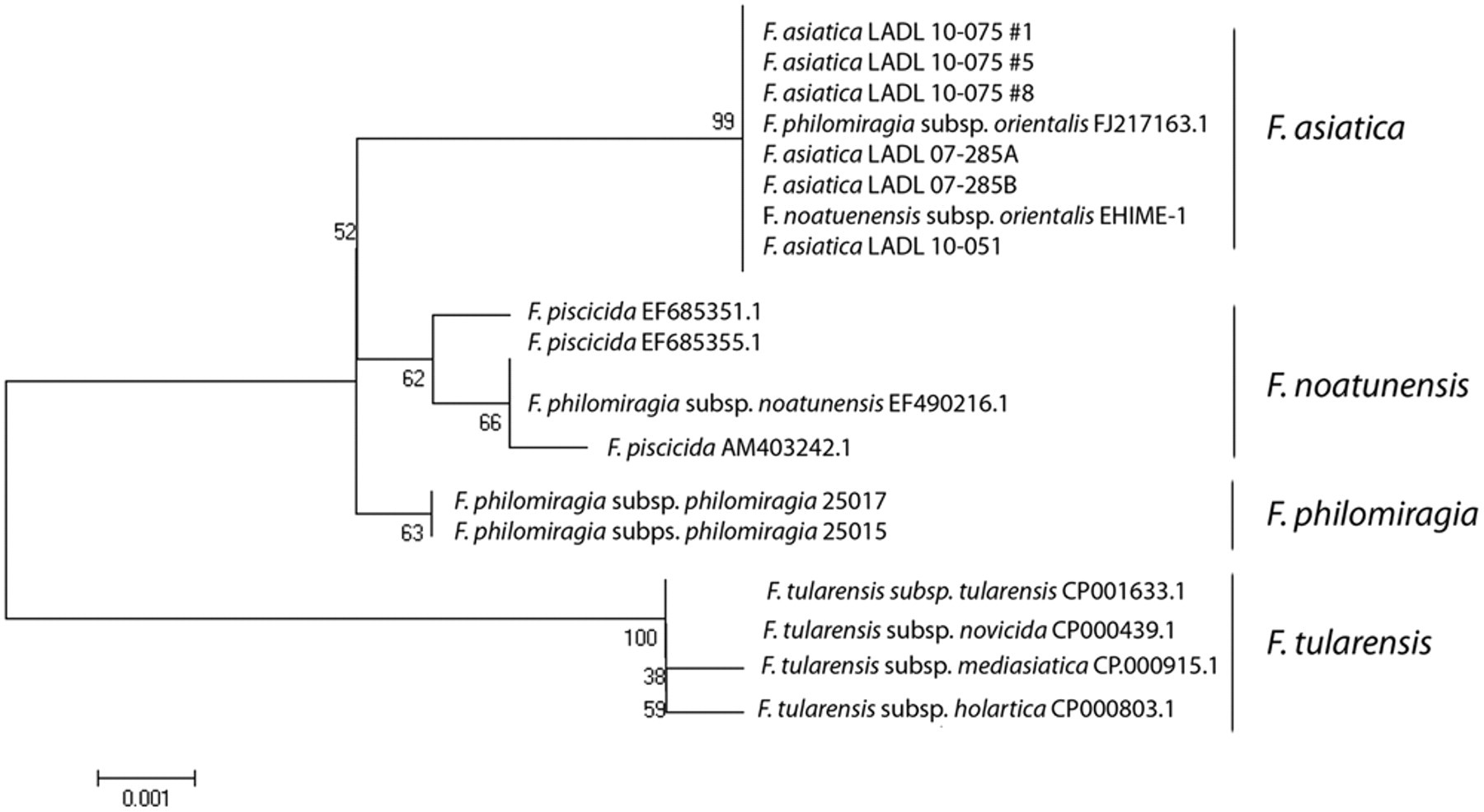
Evolutionary relationships of 17 members of the genus Francisella according to comparison of the partial sequence analysis of the 16S ribosomal RNA. The evolutionary history was inferred using the neighbor-joining method. The bootstrap consensus tree inferred from 1,000 replicates is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitions reproduced in less than 50% bootstrap replicates are collapsed. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) is shown next to the branches. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 1,264 positions in the final dataset. Phylogenetic analyses were conducted in MEGA4. 5,17,21,22
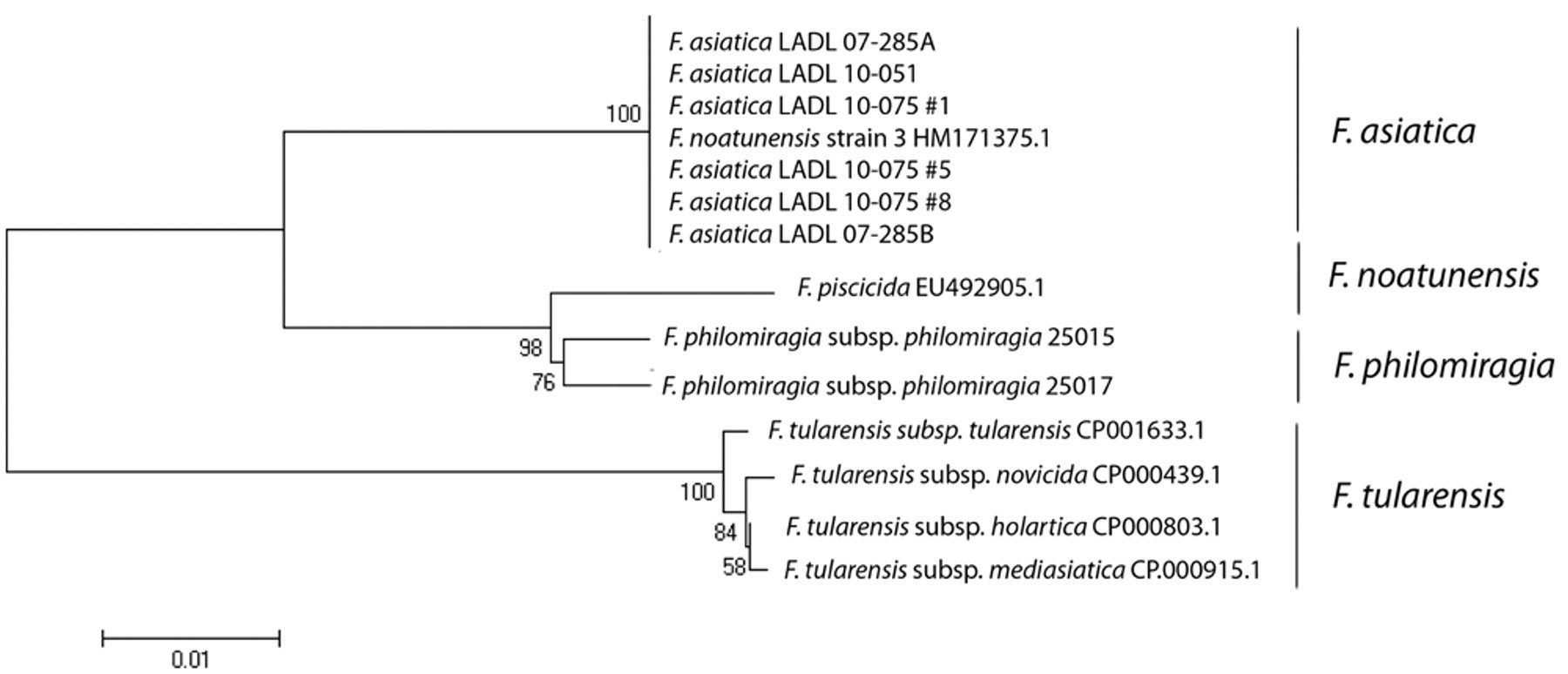
Evolutionary relationships of 13 members of the genus Francisella according to comparison of the partial sequence analysis of the intracellular growth loci C gene (iglC). The evolutionary history was inferred using the neighbor-joining method. The bootstrap consensus tree inferred from 1,000 replicates is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitions reproduced in less than 50% bootstrap replicates are collapsed. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) is shown next to the branches. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 606 positions in the final dataset. Phylogenetic analyses were conducted in MEGA4. 5,17,21,22
Intensification of aquaculture practices utilizing water reuse systems may lead to a reduction in water quality and elevated stress levels in farmed fish. Such changes have resulted in an increase in the incidence and severity of disease agents in tilapia. 4,7,11-13 Francisella asiatica is a recently recognized, highly virulent fish pathogen that has decimated tilapia farms in many countries. Like F. asiatica in tilapia (warm-water), other Francisella spp. are known to cause significant disease in cold-water species. 14,16 Currently, the number of species capable of being infected is unknown, but it is likely that more species will be added to a growing list. Fry imported to the farm in the current report were found to be free of Francisella at arrival. The presence of severe and widespread disease in young tilapia indicates that infection can progress rapidly with abundant production of bacteria in hosts. More research is necessary to identify the initial source of such emerging pathogens, and to understand the pathogenesis of the disease in fish. Moreover, it is reasonable to speculate that the extensive damage caused by F. asiatica in hematopoietic tissues (anterior kidney and spleen) may be immunosuppressive, and make the immunocompromised fish susceptible to fewer numbers of other opportunistic bacteria (V. mimicus and Aeromonas spp.) that may be found in system water on tilapia farms. Future research in this area will help clarify this hypothesis. An autogenous bacterin e against S. iniae and V. vulnificus administered by injection seemed to afford some protection as these organisms were only found on rare occasions from moribund specimens submitted from the farm to the LADL after the vaccination program started; however, mortalities attributed to francisellosis and secondary organisms increased during the study period.
The current project reports the first isolation of F. asiatica from cultured tilapia in the United States. In previous reports, Piscirickettsia-like organisms were associated with epizootics of systemic diseases that caused mortality in cultured tilapia in the continental United States. 12 Unfortunately, neither the isolation of the bacteria in bacteriological media, nor the molecular identification and characterization of the organisms were possible at that time. 12 Diagnosis of piscine francisellosis has been challenging due to various issues, namely the highly fastidious properties of the microorganism, low numbers of bacteria needed to cause significant mortality, and relatively little information available to the scientific community due to the emergent nature of the disease. The present report highlights the value and necessity of molecular diagnostic tools (conventional and real-time PCR) for rapid and accurate diagnosis of fish diseases.
Footnotes
a.
Trypticase soy agar with 5% sheep blood, Remel Inc., Lenexa, KS.
b.
Cystine heart agar supplemented with bovine hemoglobin solution, BD, Sparks, MD.
c.
Mueller-Hinton II cation adjusted broth supplemented with 2% IsoVitaleX and 0.1% glucose, BD, Sparks, MD.
d.
High Pure PCR Template Preparation Kit, Roche Diagnostics, Mannheim, Germany.
e.
Autogenous bacterin against Streptococcus iniae and Vibrio vulnificus, Aqualife Sciences Inc., Kirkland, WA.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.