
Abstract
A disease of the nervous system is reported in goats in the semiarid region of northeastern Brazil. Histological examination showed diffuse vacuolation of neurons and epithelial cells of the pancreas, thyroid, renal tubules, and liver. The swainsonine-containing plant Ipomoea verbascoidea was found on both farms where the goats originated. This plant was experimentally administered to 3 goats, inducing clinical signs and histologic lesions similar to those observed in spontaneous cases. On the lectin histochemical analysis, cerebellar cells and pancreatic acinar cells gave positive reactions to Triticum vulgaris agglutinin (WGA), succinylated Triticum vulgaris agglutinin (sWGA), Lens culinaris agglutinin (LCA), Canavalia ensiformis agglutinin (ConA), Pisum sativum agglutinin (PSA), Ricinus communis agglutinin (RCA120), Arachis hypogaea agglutinin (PNA), and Phaseolus vulgaris erythroagglutinin (PHA-E) suggesting storage of α-fucose, α-D-mannose, α-D-glucose, β-D-N-acetyl-glucosamine, N-acetyl-galactosamine, and acetyl-neuraminic acid. This pattern of lectin staining partially agrees with results previously reported for poisoning by swainsonine-containing plants. The chemical analysis of dried leaves of I. verbascoidea detected swainsonine (0.017%), calystegine B1 (0.16%), calystegine B2 (0.05%), and calystegine C1 (0.34%). It is concluded that I. verbascoidea causes α-mannosidosis in goats.
Introduction
Acquired α-mannosidosis is usually induced by the ingestion of plants containing swainsonine, including Swainsona spp. in Australia, Astragalus spp. and Oxytropis spp. in South and North America, China, and Africa,10,17,20,21and Ipomoea carnea subsp. fistulosa, 1 Ipomoea sericophylla,2,3,22 Ipomoea riedelii,2,3 Turbina cordata, 7 and Sida carpinifolia 11 in Brazil. The aim of the current study is to report the poisoning by Ipomoea verbascoidea (order Solanales, family Convolvulaceae), as a previously unknown cause of induced α-mannosidosis in goats.
Material and methods
Eight farms located in the municipality of Sertânia, state of Pernambuco, Brazil, were visited from September 2008 to September 2010, to determine disease epidemiology and to observe clinical signs of an outbreak of a suspected nervous system disorder in goats. Seven goats were sedated with 2% xylazine hydrochloride, euthanized with an overdose of 5% sodium pentobarbital, and necropsied. After fixation, the central nervous system (CNS) was sectioned in transversal cuts of 4–5 mm. Samples of cervical, thoracic, and lumbar spinal cord, medulla oblongata, pons, rostral colliculus, thalamus, cerebral cortex, cerebellar peduncles, cerebellum, trigeminal ganglia, retina, liver, kidneys, lungs, lymph nodes, spleen, thyroid, adrenal gland, stomach, celiac ganglion, large and small intestines, heart, skeletal muscle, and peripheral nerves were fixed in 10% buffered formalin with 4% phosphate, embedded in paraffin, cut at 4–6 µm, and stained by hematoxylin and eosin. Other fragments were fixed in 4% paraformaldehyde in phosphate buffered saline (PBS), cut at 3 µm, and stained with hematoxylin–phloxine and periodic acid–Schiff.
Samples of I. verbascoidea were collected from the paddocks in which the outbreaks occurred. Afterward, they were sent to the Federal Rural University of Pernambuco, Recife, Pernambuco, Brazil, for identification, and to the Professor Vasconcelos Sobrinho Herbarium of UFRPE, for registration (voucher no. 50017). Animal care and experimental design were approved by the Animal Ethics Committee of the Federal Rural University of Pernambuco and by the University of Cuiabá, Mato Grosso, Brazil.
For the experimental reproduction of the disease, I. verbascoidea was collected in the municipality of Sertânia, Pernambuco, Brazil. Experiments were completed using both green and desiccated plants. For experiments using the green plant, 6 kg of I. verbascoidea were collected daily and supplied as the main forage for 2 goats (goats 8 and 9) during 84 and 35 days, respectively. The fresh green plant was fed daily, once a day, in the morning, immediately after collection. The animals ingested the plant in a 2–3 hr period. After ingesting the plant, commercial food in the proportion of 1% of the goats’ weight was administered. Water and mineral salt ad libitum were also available.
To obtain dry material, 25 kg of green plant were collected and placed in an open-air shaded area for drying. The 25 kg of fresh plant resulted in 7.8 kg of dry plant (31.2% of dry matter). After drying, the plants were milled, mixed with the commercial food in the proportion of 50% I. verbascoidea and 50% commercial food, and administered to a goat (goat 10) for 42 days at a rate of 2% live weight. Two goats (goats 11 and 12), used as controls, received commercial food in the proportion of 2% live weight. The experimental goat and the control goats also received Cynodon dactylon (Tifton) hay ad libitum. The amount of plant material consumed daily was recorded for each animal, and the goats were weighed at the beginning and end of the experiment. The average daily dose of I. verbascoidea received was calculated based on total amount of plant consumed divided by the intake period (days; Table 1). The goats were examined daily including an examination of the CNS, as well as the administration of the head-raising test, which consists of raising the head of the animal for approximately 1 min, and then suddenly releasing it. 25
Daily means and total Ipomoea verbascoidea dose consumed by the goats: animal weight at the beginning and end of the experiment, clinical signs, and poisoning outcome.*
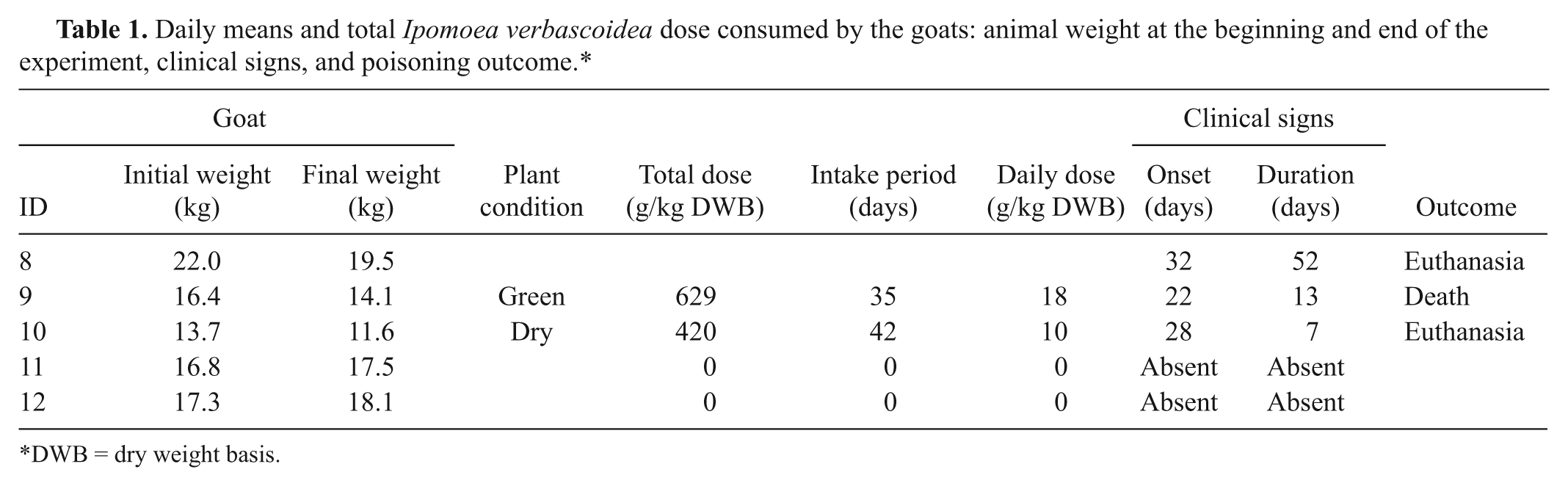
DWB = dry weight basis.
At the end of the experiment, the experimental animals were euthanized, necropsied, and examined histologically as mentioned above. Samples of the cerebellum, pancreas, and thyroid gland were treated with 0.3% hydrogen peroxide for 30 min at room temperature. After blocking, the samples were submitted to antigen recovery for 15 min in citrate buffer (pH 6.0) in a water bath at 100°C. The sections were treated with 5% skimmed milk at room temperature for 30 min and incubated with 14 lectins: Canavalia ensiformis agglutinin (concanavalin A [ConA]), Triticum vulgaris (wheat germ agglutinin [WGA]), succinylated Triticum vulgaris agglutinin (sWGA), Lens culinaris agglutinin (LCA), Glycine max (soybean agglutinin [SBA]), Dolichos biflorus agglutinin (DBA), Ulex europaeus agglutinin 1 (UEA 1), Ricinus communis agglutinin 1 (RCA120), Arachis hypogaea (peanut agglutinin [PNA]), Griffonia simplicifolia agglutinin 1 (GSA I), Pisum sativum agglutinin (PSA), Phaseolus vulgaris erythroagglutinin (PHA-E), Phaseolus vulgaris leucoagglutinin (PHA-L), and Sophora japonica agglutinin (SJA a ; Table 2). Afterward, the sections were treated with streptavidin–peroxidase conjugate b for 20 min and revealed with chromogen 3,3-diaminobenzidine b or red chromogen. c Sections were counterstained with Mayer hematoxylin, dehydrated, imbibed in xylene, and examined under a light microscope. The lectins were used at the dilution of 5 μg/ml in PBS, with the exception of C. ensiformis, which was used at the dilution of 0.5 mg/ml PBS. 4 Sections of tissues of a nonaffected goat (goat 11) were used as control.
Intensity of the lectin reaction tested in vacuolated cells of the cerebellum and pancreas gland of goats naturally poisoned by Ipomoea verbascoidea.*
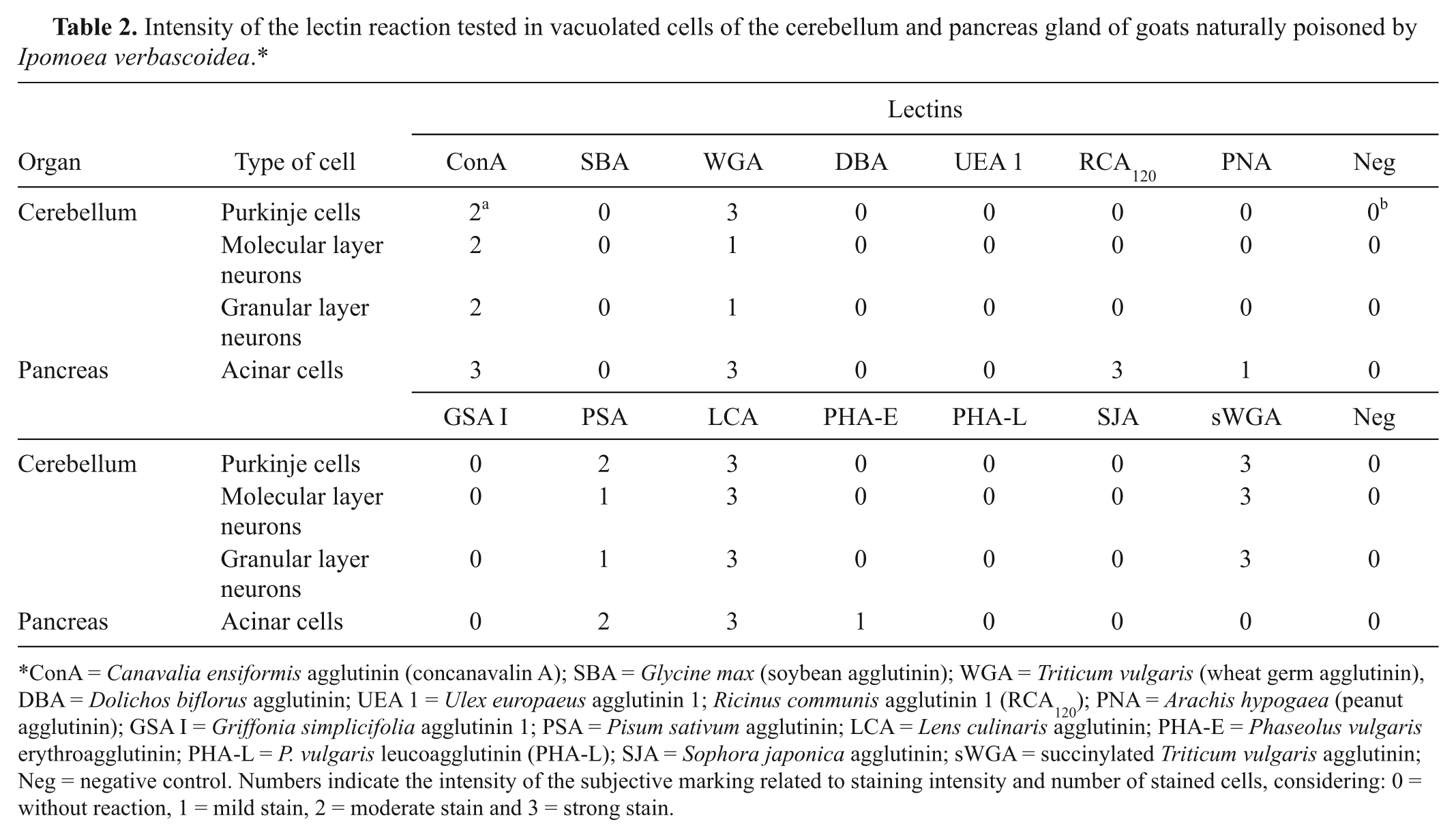
ConA = Canavalia ensiformis agglutinin (concanavalin A); SBA = Glycine max (soybean agglutinin); WGA = Triticum vulgaris (wheat germ agglutinin), DBA = Dolichos biflorus agglutinin; UEA 1 = Ulex europaeus agglutinin 1; Ricinus communis agglutinin 1 (RCA120); PNA = Arachis hypogaea (peanut agglutinin); GSA I = Griffonia simplicifolia agglutinin 1; PSA = Pisum sativum agglutinin; LCA = Lens culinaris agglutinin; PHA-E = Phaseolus vulgaris erythroagglutinin; PHA-L = P. vulgaris leucoagglutinin (PHA-L); SJA = Sophora japonica agglutinin; sWGA = succinylated Triticum vulgaris agglutinin; Neg = negative control. Numbers indicate the intensity of the subjective marking related to staining intensity and number of stained cells, considering: 0 = without reaction, 1 = mild stain, 2 = moderate stain and 3 = strong stain.
The swainsonine content of I. verbascoidea was measured by liquid chromatography–mass spectrometry (LC-MS) using procedures previously described. 12 In general, duplicate samples (100 mg) of the air-dried plant material were extracted, and the swainsonine was isolated by solid phase extraction using ion exchange resin. The final aqueous extract was then analyzed by LC-MS. The presence of swainsonine was verified using gas chromatography–mass spectrometry (GC-MS) after a portion of the aqueous extract had been dried and the residue derivatized by the addition of N,O-Bis(trimethylsilyl)trifluoroacetamide (BSTFA) and pyridine to convert swainsonine to its trimethylsilyl ether derivative. Calystegines were analyzed by GC-MS from the same BSTFA-derived sample. Identification of calystegines was made by comparison of relative retention time and mass spectrum to previously published data,23,24 and their concentration estimated based on peak area measurement relative to previously determined swainsonine concentration.
Results
The plant was identified as I. verbascoidea Choisy (Fig. 1), a sub-erect to climbing shrub, with erect, decumbent, or rambling stems, up to 1–5 m tall, covered with a yellowish or white woody, somewhat floccose tomentum, as are the leaves, petioles, peduncles, bracts and, generally, the sepals. The leaves are oblong or ovate-circular, the flowers are solitary or in more or less 3 flowered cymes, and the seeds are ovoid, brown, densely covered with long white or sometimes fulvous cottony hairs. 29

Ipomoea verbascoidea Choisy; Municipality of Sertânia, State of Pernambuco, Brazil. The plant is a liana that grows and covers other plants. Inset: detail of I. verbascoidea leaves and flower.
The disease was observed on 2 farms. On farm A, within 2 years, 7 (8%) out of 89 goats of different ages presented neurological alterations, and 4 (4.5%) died. On farm B, within the same period, 32 (80%) out of 40 goats presented neurological alterations, and 24 (60%) died. At farm A, an intense growth of I. verbascoidea was observed in the grazing area of the goats during a visit in the rainy season. Three other species of Ipomoea (I. bahiensis, I. martii, and I. sericophylla) were also found, but in lower abundance. On farm B, only I. verbascoidea was found. All these species of Ipomoea were found at 6 other farms in the region where the farmers reported in previous years the occurrence of a disease similar to that observed on farms A and B.
Clinical signs consisted of head tremors, motor incoordination with ataxia and dysmetria especially in the posterior limbs, lateral walking, abnormal postures, and imbalance followed by falls (Fig. 2). The clinical signs were more evident when the goats were forced to move or when they were stressed. Other alterations, such as progressive weight loss, rough coat, prolonged recumbency, and death were also observed in the animals affected with the disease. On farm A, 14% of the goats had complete recovery from the nervous alterations, and 28% had sequelae, consisting mainly of muscle and head tremors and a tottering walk. On farm B, only 3% of the goats that showed cerebellar dysfunction had complete recovery, and 22% had sequelae.

Goat naturally poisoned by Ipomoea verbascoidea showing paraparesis of hind legs.
The intoxication was seasonal occurring during the rainy season, which extends from December–January through April–May, when the plants are growing. Sickness reoccurred in the goats that were affected in the previous year when they were re-exposed to I. verbascoidea in pasture. Three goats from farm A and 4 from farm B, which presented moderate or intense neurological signs, were necropsied, and significant gross alterations were not observed.
Microscopically, neurons with diffuse fine cytoplasmatic vacuolation were detected in several areas of the CNS, mainly in the Purkinje neurons (Fig. 3) and neurons of the cerebellar nuclei, pons, and medulla oblongata. Axonal spheroids were observed in the cerebellar granular layer, as well as a white cerebellar substance, cerebellar peduncles, and cerebellar nucleus. Vacuolations were also present in neurons of the cerebral cortex, trigeminal and celiac ganglion, and neurons of spinal cord. There was also vacuolation in the pancreatic acinar cells, thyroid follicular cells, hepatocytes, and renal tubular epithelial cells. These lesions were not observed in goats 3 and 5, which were prevented from consuming I. verbascoidea for more than 15 days prior to euthanasia. Results of the experimental reproduction of the disease, including the average daily consumption, the total I. verbascoidea dose consumed by the goats, the animals’ weight at the beginning and end of the experiment, the clinical signs, and the outcome of the poisoning are summarized in Table 1.
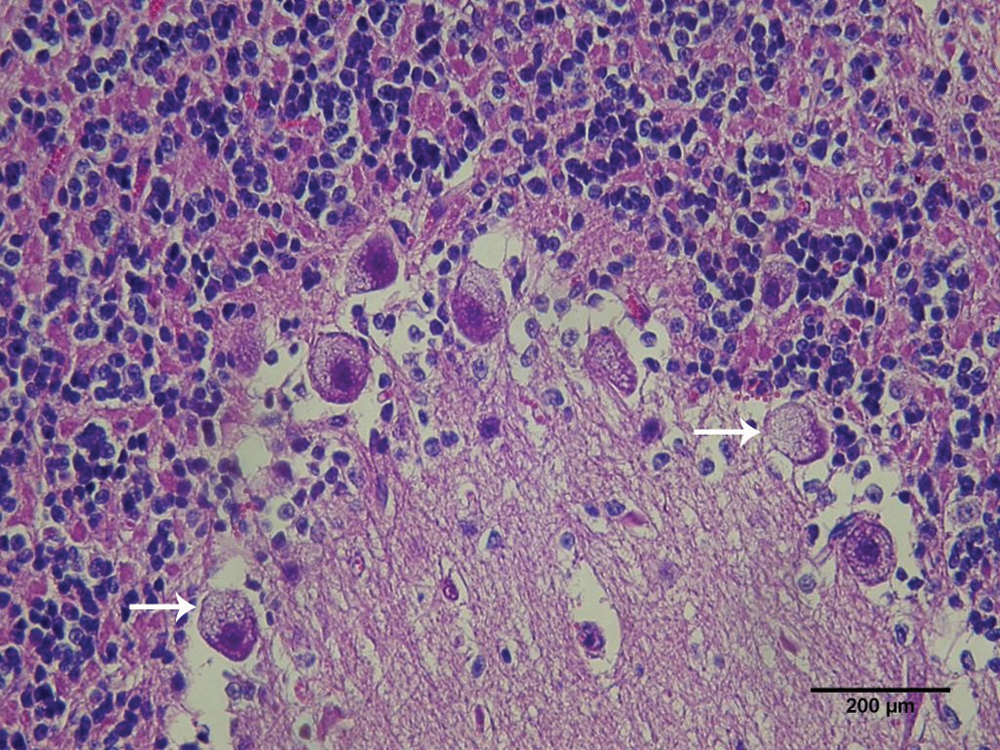
Goat. Purkinje neurons with thin cytoplasmatic vacuolation (arrows). Hematoxylin and eosin stain. Bar = 200 µm.
Goats 8 and 9, which received green plant, presented the first clinical signs on the 32nd and 22nd days, respectively. Goat 10, which was fed dry I. verbascoidea, first displayed clinical signs on the 28th day of the experiment. Initially, clinical signs consisted of rough coat, depression, and mild motor incoordination. In the 3 experimental goats, the neurologic signs were similar; 3–7 days after initial signs, the 3 goats showed motor incoordination, hypermetria, spastic paresis, ataxia, nystagmus, head tremors, abnormal postures, side walking, extended lateral recumbency, and anorexia. When the goats were forced to move, they lost their balance and fell to the side or back. All goats examined were positive when given the head-raising test. Goat 9 also presented liquid and greenish feces, lasting for 5 days, recovering on the 6th day following the start of the diarrhea. During this period, this goat ingested the plant normally.
No significant alterations were observed at necropsy. Microscopic lesions similar to those observed in spontaneous cases were observed in the affected goats, except in goat 9, which was prevented from eating the plant for 15 days prior to euthanasia. Results of the lectin histochemical analysis are presented in Table 2.
Chemical analysis of I. verbascoidea dried leaves showed a swainsonine concentration of 0.017% (dry weight basis). The estimated concentrations of calystegines were 0.16% for calystegine B1, 0.05% for calystegine B2, and 0.34% for calystegine C1.
Discussion
The characteristic histologic lesions, the experimental reproduction of the disease, and the detection of swainsonine in the plant demonstrated that I. verbascoidea causes α-mannosidosis in goats. Ipomoea verbascoidea is widely found in Africa, from the tropics to the southern region, 13 and in Brazil in the northeastern semiarid region of the country. 29 Other species of Ipomoea were found on some of the farms visited. Ipomoea bahiensis and I. martii have not been tested for their toxicity, and I. sericophylla was recently reported as the cause of neurological disease in 2 other farms in Sertânia, Brazil. 22
Ipomoea verbascoidea is found only during the rainy season, and poisoning occurs during this period, with higher frequency between March and May. Ipomoea riedelii also causes poisoning during the rainy season in the state of Paraíba (neighboring state of Pernambuco).2,3 In contrast, Ipomoea carnea subsp. fistulosa, which is found mainly around natural springs or partially flooded areas throughout the year, can cause poisoning mainly in the dry season when there is a shortage of forage.1,28,32
Clinical signs and histologic lesions observed in spontaneous and natural cases of I. verbascoidea poisoning are similar to those reported in other swainsonine-containing plants mentioned in the introduction.1,3,5-7,16,28,31 The positive results on the lectin histochemical analysis of cerebellar cells, pancreatic acinar cells, and follicular thyroid cells for ConA, WGA, sWGA, LCA, PSA, PHA-E, RCA120, and PNA suggest the storage of compounds rich in α-fucose, α-D-mannose, α-D-glucose, β-D-N-acetyl-glucosamine, and N-acetyl-galactosamine, as well as acetyl-neuraminic acid residues. 14 This pattern of lectin staining partially agrees with results previously reported for poisoning by swainsonine-containing plants, generally, and for mannosidosis in human beings, cats, and calves.1-3,5,8,11,30
In the present study, the concentrations of swainsonine, calystegine B1, calystegine B2, and calystegine C1 in dried leaves of I. verbascoidea were 0.017%, 0.16%, 0.05%, and 0.34%, respectively. Swainsonine concentrations are variable in swainsonine-containing plants in Brazil.2,3,7,15 A threshold for toxicity is difficult to establish for swainsonine because there is a high variability in alkaloid concentration in all of these plants. A conservative approach suggests that levels exceeding 0.001% of dry matter are most likely toxic. 23 In the experiments reported herein, it can be estimated that the goat that ingested the dry plant (10 g/kg body weight), with a concentration of 0.017% of swainsonine, ingested 1.7 mg of swainsonine daily.
Calystegines B1, B2, and B 3 were also found in the dry leaves of I. verbascoidea. Calystegines B1, B2, B3, and C1 are reportedly found in I. fistulosa 15 and I. sericophylla,2,3 but not in I. riedelii and T. cordata.2,3,7 The toxic effects of calystegines in animals were not determined, but calystegines are inhibitors of glycosides, mainly β-glucosidase, and α-galactosidase. The occurrence of ataxia in cattle was associated with the presence of different calystegines in Ipomoea spp. 9 In contrast, calystegines B2 and C1 showed no inhibition of β-glucosidase in human lymphoblasts, but increased the activity of the enzyme, suggesting that calystegines have little risk of inducing intoxication of livestock. 19
An important observation in I. verbascoidea poisoning was the absence of cytoplasmic vacuolization in nerve and epithelial cells in goats 3, 5 (naturally poisoned), and 9 (experimentally poisoned). These 3 goats were prevented for 15 or more days from eating the plant suggesting that a period of 15 days is sufficient for the disappearance of cytoplasmic vacuolization in neurons and epithelial cells. This observation should be considered in the histologic diagnosis of the disease.
Goats appear to develop a preference for grazing on I. verbascoidea, resulting in the development of neurologic disease. This behavior, sometimes reported as addiction, had also been observed in ruminants grazing other swainsonine-containing plants, including Swainsona spp., 18 locoweeds,26,27 I. fistulosa subsp. fistulosa,8,28,32 I. sericophylla, I. riedelii,2,3,22 S. carpinifolia,5,6 and T. cordata. 7 Because of this addictive behavior, goats should not be returned to areas containing Ipomoea spp. plants after clinical recovery has occurred. 3
Footnotes
Acknowledgements
The authors thank Dr. Marcelo Ataíde Teixeira, senior botanist of the Federal Rural University of Pernambuco (UFRPE), Recife, Pernambuco, Brazil, for identification of I. verbascoidea samples, and the Professor Vasconcelos Sobrinho Herbarium of UFRPE, for registration.
Notes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. AQ:Please verify that this statement is accurate and correct.
Funding was provide by the Brazilian Foundation Agency FACEPE (grant no. 0760-5.05/09).