
Abstract
Porcine periweaning failure-to-thrive syndrome (PFTS), an increasingly recognized syndrome in the swine industry of North America, is characterized by the anorexia of nursery pigs noticeable within 1 week of weaning, and progressive loss of body condition and lethargy during the next 1–2 weeks. Morbidity caused by PFTS is moderate, but case fatality is high. The etiology of PFTS is presently unknown and may include infectious agent(s), noninfectious factors, or both. PFTS was identified in a high health status farm with good management in early 2007. A diagnostic investigation was undertaken to identify the pathological lesions of, and infectious agents associated with, pigs demonstrating typical clinical signs. Affected (PFTS-SICK) and unaffected (PFTS-HLTHY) pigs from an affected farm, and unaffected pigs from 2 unaffected farms, were examined. The most prevalent lesions in PFTS-SICK pigs were superficial lymphocytic fundic gastritis, atrophic enteritis, superficial colitis, lymphocytic and neutrophilic rhinitis, mild nonsuppurative meningoencephalitis, and thymic atrophy. Rotavirus A and Betacoronavirus 1 (Porcine hemagglutinating encephalomyelitis virus) were identified only in PFTS-SICK pigs, but the significance of the viruses is uncertain because PFTS is not consistent with the typical presentation following infection by these pathogens. Porcine reproductive and respiratory syndrome virus, Porcine circovirus-2, Influenza A virus, Alphacoronavirus 1 (Transmissible gastroenteritis virus), Torque teno virus 1, Brachyspira hyodysenteriae, and Brachyspira pilosicoli were not identified in PFTS-SICK pigs. Suid herpesvirus 2 (Porcine cytomegalovirus), Porcine enteric calicivirus, Torque teno virus 2, pathogenic Escherichia coli, and coccidia were detected in both PFTS-SICK and PFTS-HLTHY pigs. It was concluded that there is a lack of compelling evidence that PFTS is caused by any of these pathogens.
Introduction
“Failure-to-thrive” is a term in the pork industry that refers to pigs that appear not to eat after weaning, and subsequently lose body condition and die with no other known etiology. Importantly, failure-to-thrive pigs are not lightweight pigs (i.e., runts) at birth or weaning. Commonly referred to as “starve-outs” by producers, such pigs have been observed at a low prevalence (up to 0.5% of total weaned) for many years. In the past, the condition received little attention since the prevalence and economic loss was low. However, herd outbreaks of failure-to-thrive pigs, resulting in nursery mortality that has risen well above historical baseline levels, were recently recognized in a number of North American farms (Dufresne L, Fangman TJ, Henry S: 2008, Post-weaning catabolic syndrome: complexities and perspectives. In: Proceedings of the Allen D. Leman Swine Conference, pp. 79–85; Gauvreau H, Harding J: 2008, Why are these nursery pigs dying? An ongoing field investigation into a farm with elevated nursery mortality associated with non-PRRS/PCV2 post weaning starvation. In: Proceedings of the Western Canadian Association of Swine Veterinarians Conference, pp. 47–54) The condition was recently named “periweaning failure-to-thrive syndrome” (PFTS), and a clinical case definition proposed as follows: “PFTS is characterized clinically by the progressive debilitation of weanling (nursery) pigs in the absence of discernible and detrimental infectious, nutritional, managemental, or environmental factors that can explain the clinical syndrome. At weaning, affected pigs are of average to above average body weight, and neither affected pigs nor their cohorts show evidence of residual illnesses from the suckling phase. Within 7 days of weaning, affected pigs are anorexic and lethargic. They deteriorate and within 2–3 weeks of weaning demonstrate marked muscle weakness and loss of body condition. Some affected pigs in all affected farms show repetitive oral behavior such as licking, chewing or chomping. In affected farms, batch morbidity and mortality varies over time, but case fatality is high” (reprinted with permission from Huang Y, Henry S, Friendship B, et al.: 2011, Case definition and diagnostic guidelines for Porcine “Periweaning Failure to Thrive Syndrome” (PFTS). J Swine Health Prod 19. In press). The etiology of PFTS is presently unknown and may include infectious agent(s), noninfectious factors, or both.
A 100-sow farrow-to-finish farm has experienced disease causing fluctuating nursery mortality beginning early 2007. The clinical presentation of this disease meets the definition of PFTS quoted above. This farm is historically free of Mycoplasma hyopneumoniae and Porcine reproductive and respiratory syndrome virus (PRRSV). Nursery mortality was elevated by 3.7 fold (7.2% for 2007–2009; 1.9% for 2004–2006), mostly attributable to failure-to-thrive piglets. Chomping behavior was observed in the failure-to-thrive pigs in this farm. Common mycotoxins (tested by the Veterinary Diagnostic Laboratory of North Dakota State University, Fargo, North Dakota) and herbicides (tested by ALS Laboratory Group, Saskatoon, Saskatchewan, Canada) in feed and water, respectively, were negative. Various managemental and medical interventions to control PFTS had been attempted, but failed to prevent mortality. The interventions included ventilation adjustments to prevent piglet chilling, changes in starter feed and water source, antibiotic treatments, H3N2 swine influenza vaccination of the breeding herd, and Porcine circovirus-2 (PCV-2) vaccination of piglets at weaning. Although thorough desiccation of the farrowing and nursery rooms with aerosolized hydrated lime appeared to be beneficial during periods of elevated PFTS mortality, the cyclical nature of clinical signs and mortality on the farm made it difficult to evaluate the true impact of this procedure. On the other hand, it supports the hypothesis that infectious agent(s) may be involved in pathogenesis of PFTS. The objective of the current study is to report the diagnostic investigation of PFTS on this farm with emphasis on describing the most prevalent lesions and identifying the infectious agents in PFTS-affected compared to age-matched, nonaffected pigs.
Materials and methods
Sample collection
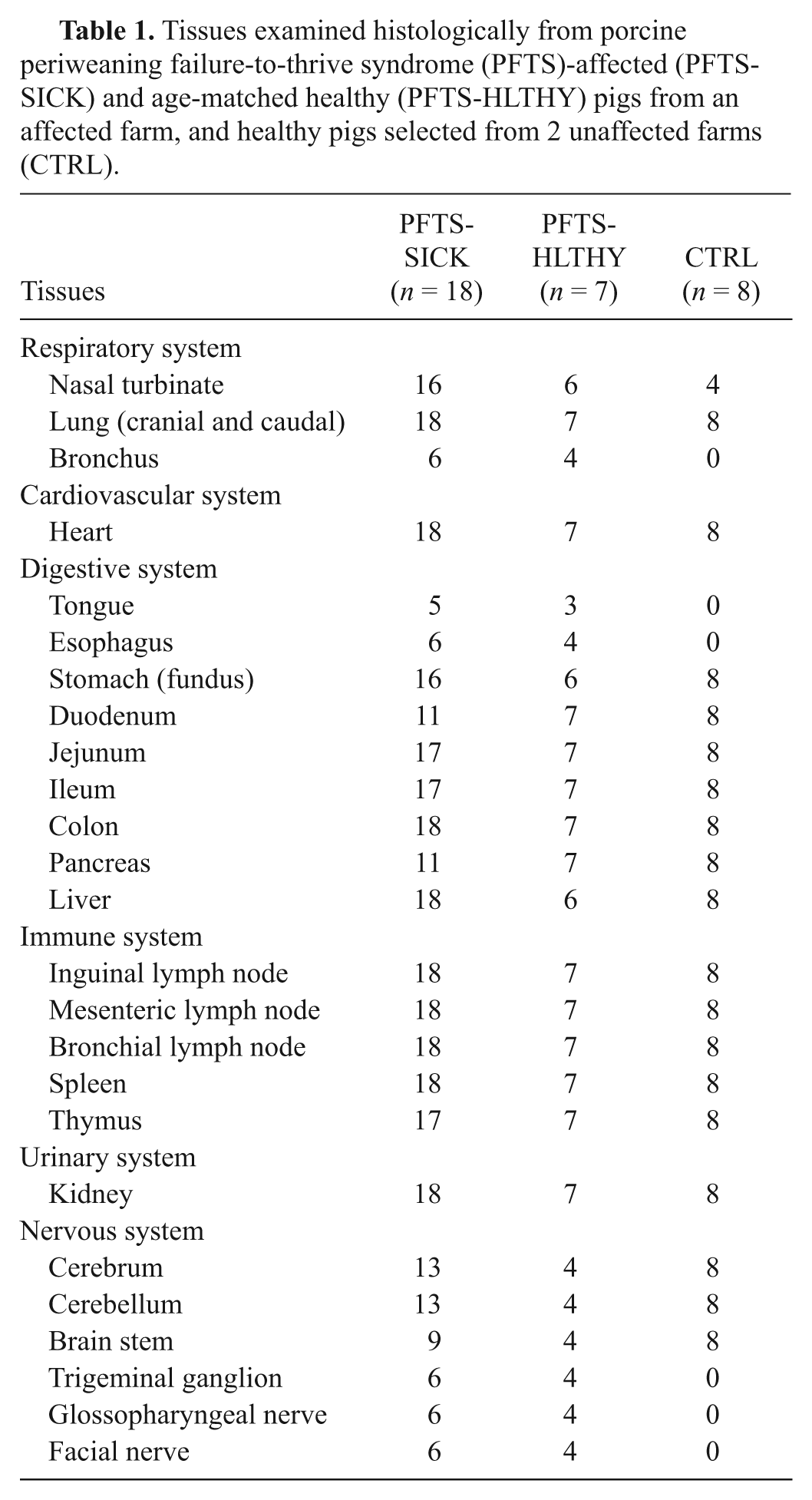
During periods of peak PFTS mortality, sick pigs with characteristic clinical signs (PFTS-SICK; n = 18), and healthy age-matched cohort pigs selected from the same weaning group and air space (PFTS-HLTHY; n = 7), ranging in age from 3 to 6 weeks old (1–3 weeks postweaning), were selected on the farm. Although chomping behavior was observed on the farm during outbreaks of PFTS, the chomping status of the pigs included in the current study was unknown as the authors failed to recognize the biological importance of this clinical sign at the onset of the investigation. Four age-matched healthy pigs from each of 2 unaffected farrow-to-finish farms were handled similarly on separate occasions and included as a comparative group in the investigation (CTRL; n = 8). PFTS-SICK and PFTS-HLTHY pigs were sedated with azaperone a (2.2 mg/kg intramuscularly) on the farm, and transported to the necropsy facility at the University of Saskatchewan (Saskatoon, Saskatchewan, Canada). The CTRL pigs were transported unsedated because the length of time in transit was shorter, thus aggression during transport was less of a concern. Necropsy examinations were performed immediately after humane euthanasia using intravenous barbiturate overdose. An extensive list of tissues (Table 1) were collected and processed routinely for bacterial culture, histology, and adjunct testing for common swine pathogens. Fecal samples were collected from a subset of PFTS-SICK (n = 5) and PFTS-HLTHY (n = 3) pigs at the time of necropsy. Fecal samples (n = 5) were collected from age-matched pigs from 1 CTRL farm but on a different day and not from the necropsied pigs.
Tissues examined histologically from porcine periweaning failure-to-thrive syndrome (PFTS)-affected (PFTS-SICK) and age-matched healthy (PFTS-HLTHY) pigs from an affected farm, and healthy pigs selected from 2 unaffected farms (CTRL).
Diagnostic tests
Adjunct diagnostic tests were performed on appropriate tissues and samples (Table 2) using published or in-house protocols. The primers and conditions of polymerase chain reaction (PCR) testing are summarized in Table 3. Immunohistochemistry (IHC) was performed using a streptavidin–biotin complex technique adapted for an automated slide stainer b as previously described. 8 The primary antibodies used for IHC in the present study are summarized in Table 4. Binding of the primary antibodies was detected using biotinylated goat anti-rabbit immunoglobulin (Ig)G and horse anti-mouse IgG, c and an avidin–biotin immunoperoxidase complex reagent, c with 3,3’-diaminobenzidine tetrahydrochloride d as the chromogen.
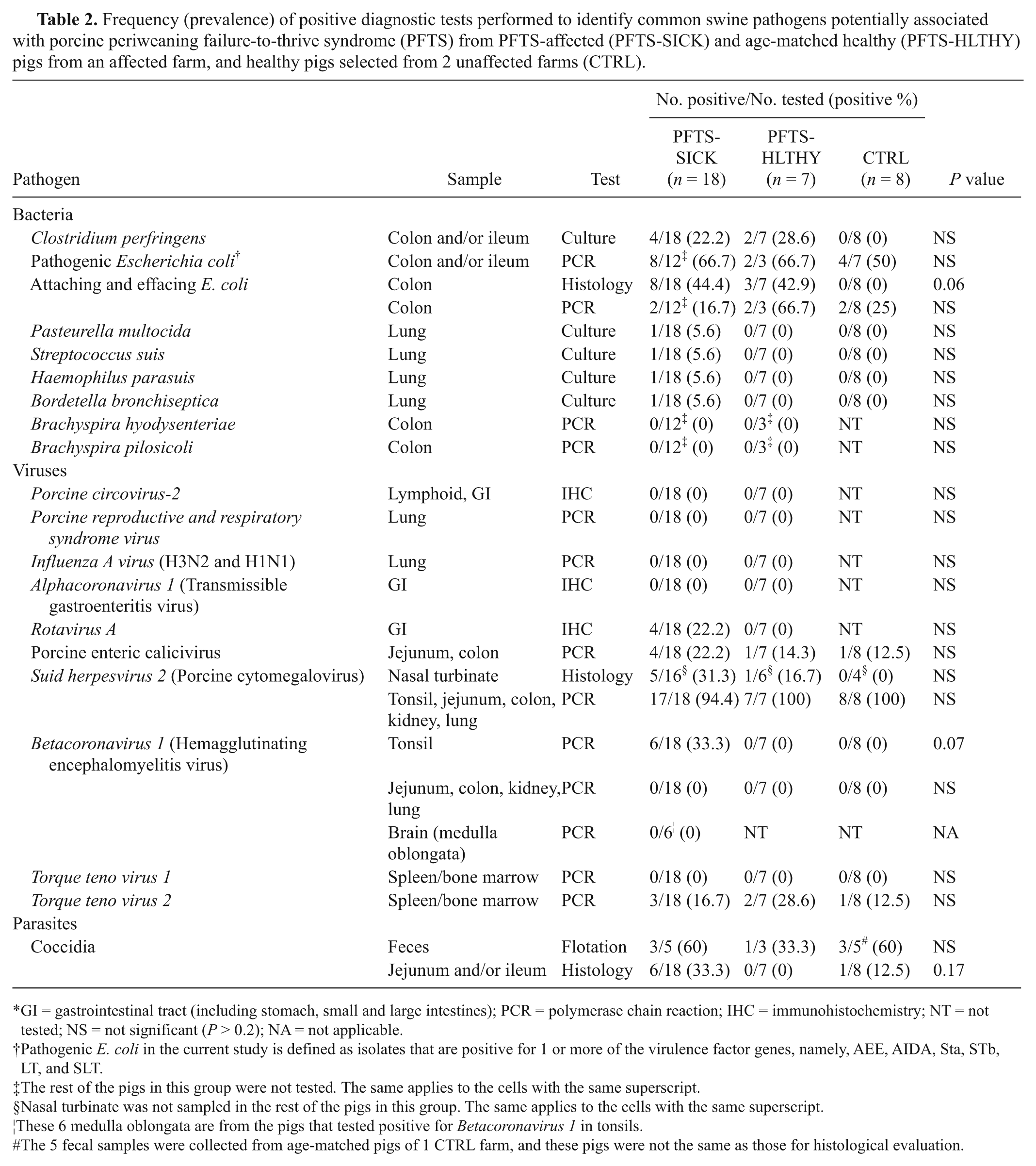
Frequency (prevalence) of positive diagnostic tests performed to identify common swine pathogens potentially associated with porcine periweaning failure-to-thrive syndrome (PFTS) from PFTS-affected (PFTS-SICK) and age-matched healthy (PFTS-HLTHY) pigs from an affected farm, and healthy pigs selected from 2 unaffected farms (CTRL).
GI = gastrointestinal tract (including stomach, small and large intestines); PCR = polymerase chain reaction; IHC = immunohistochemistry; NT = not tested; NS = not significant (P > 0.2); NA = not applicable.
Pathogenic E. coli in the current study is defined as isolates that are positive for 1 or more of the virulence factor genes, namely, AEE, AIDA, Sta, STb, LT, and SLT.
The rest of the pigs in this group were not tested. The same applies to the cells with the same superscript.
Nasal turbinate was not sampled in the rest of the pigs in this group. The same applies to the cells with the same superscript.
These 6 medulla oblongata are from the pigs that tested positive for Betacoronavirus 1 in tonsils.
The 5 fecal samples were collected from age-matched pigs of 1 CTRL farm, and these pigs were not the same as those for histological evaluation.
Summary of polymerase chain reaction (PCR) primers and conditions used in the current study.

Summary of the primary antibodies used in the current study.
Routine aerobic and anaerobic bacterial cultures were performed on the jejunum, colon, and lung. When hemolytic Escherichia coli was cultured, an agglutination test was applied to detect the presence of F4 fimbria.
The following pathogens were tested by either PCR or IHC (Table 2): PCV-2 (IHC, on lymphoid organs, stomach, small and large intestine), PRRSV (PCR, on lung), Influenza A virus (FLUAV; PCR, on lung), Alphacoronavirus 1 (ACoV-1; Transmissible gastroenteritis virus; IHC, on stomach, small and large intestine), Rotavirus A (RV-A; IHC, on stomach, small and large intestine), Porcine enteric calicivirus (PECV; PCR, on small and large intestine), Betacoronavirus 1 (BCoV-1; Porcine hemagglutinating encephalomyelitis virus; PCR, on tonsil, lung, kidney, small and large intestine), Suid herpesvirus 2 (SuHV-2; Porcine cytomegalovirus; PCR, on tonsil, lung, kidney, small and large intestine), Torque teno virus 1 and 2 (TTV-1, -2; PCR, on spleen or bone marrow), E. coli virulence factors (AEEC, AIDA, Sta, STb, LT, SLT; PCR, on cultured colonies from small and large intestine), Brachyspira hyodysenteriae and Brachyspira pilosicoli (PCR, on large intestine). Except for SuHV-2, BCoV-1, and PECV, the above tests were performed by a commercial diagnostic service. e The PCR detection of SuHV-2, BCoV-1, and PECV was performed in-house according to published methods with minor modifications (Table 3).7,18,21 Fecal flotation using Wisconsin double centrifuge technique 2 was performed to detect parasitic oocysts in the feces.
Statistical analysis
The frequency of gross and histologic lesions and pathogens in PFTS-SICK, PFTS-HLTHY, and CTRL pigs were compared using a Fisher exact test. Group differences were statistically significance if P < 0.05.
Results
Gross and histological lesions
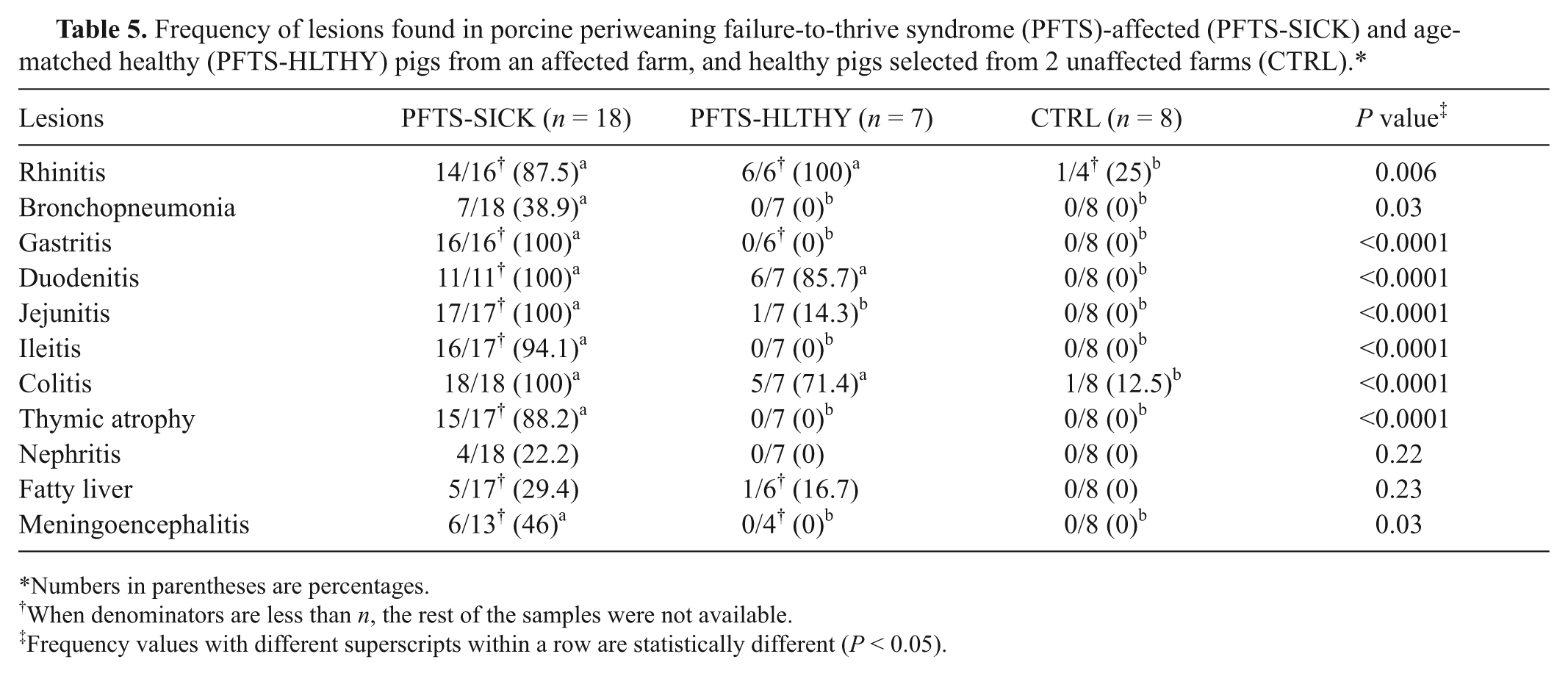
On gross examination, all of the PFTS-SICK pigs were emaciated with loss of body fat reserves. The thoracic thymus was severely atrophic and sometimes barely visible. Gastrointestinal (GI) tracts were empty, and often much smaller in diameter compared to PFTS-HLTHY and CTRL pigs. Seven of 18 (39%) PFTS-SICK pigs had mild to severe cranioventral consolidation in the lungs. No gross lesions were observed in the PFTS-HLTHY and CTRL pigs (Table 5).
Frequency of lesions found in porcine periweaning failure-to-thrive syndrome (PFTS)-affected (PFTS-SICK) and age-matched healthy (PFTS-HLTHY) pigs from an affected farm, and healthy pigs selected from 2 unaffected farms (CTRL).*
Numbers in parentheses are percentages.
When denominators are less than n, the rest of the samples were not available.
Frequency values with different superscripts within a row are statistically different (P < 0.05).
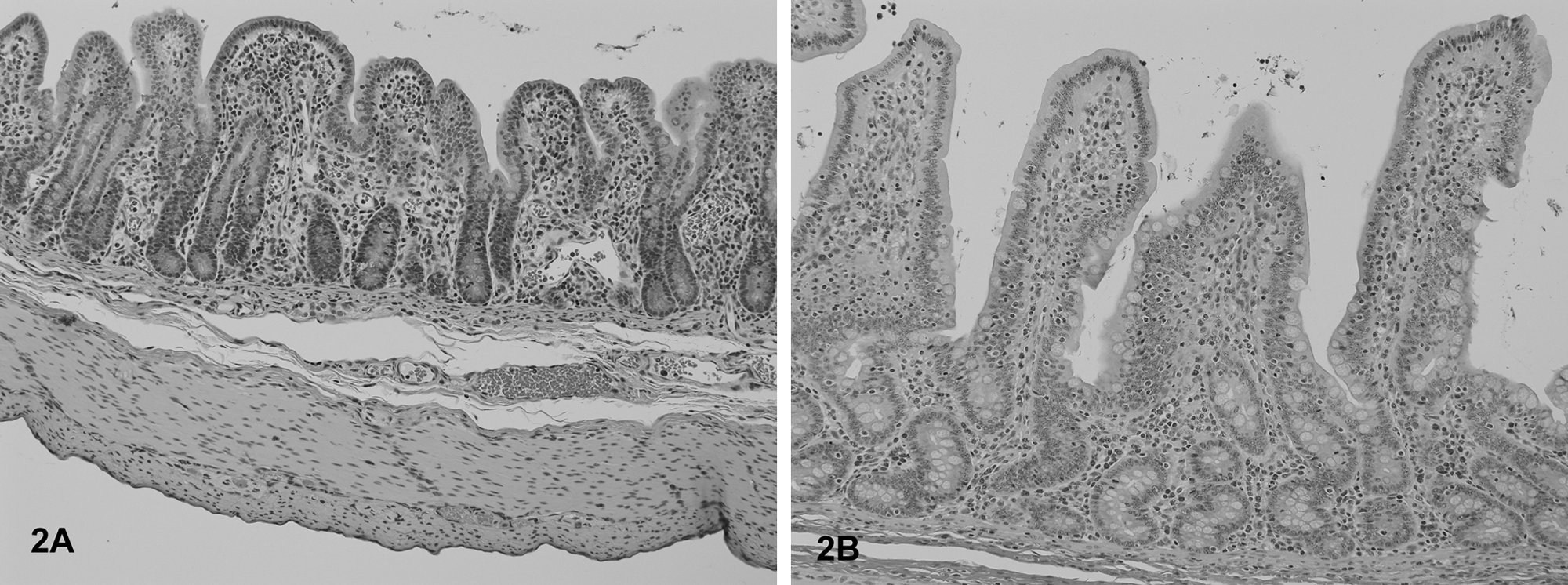
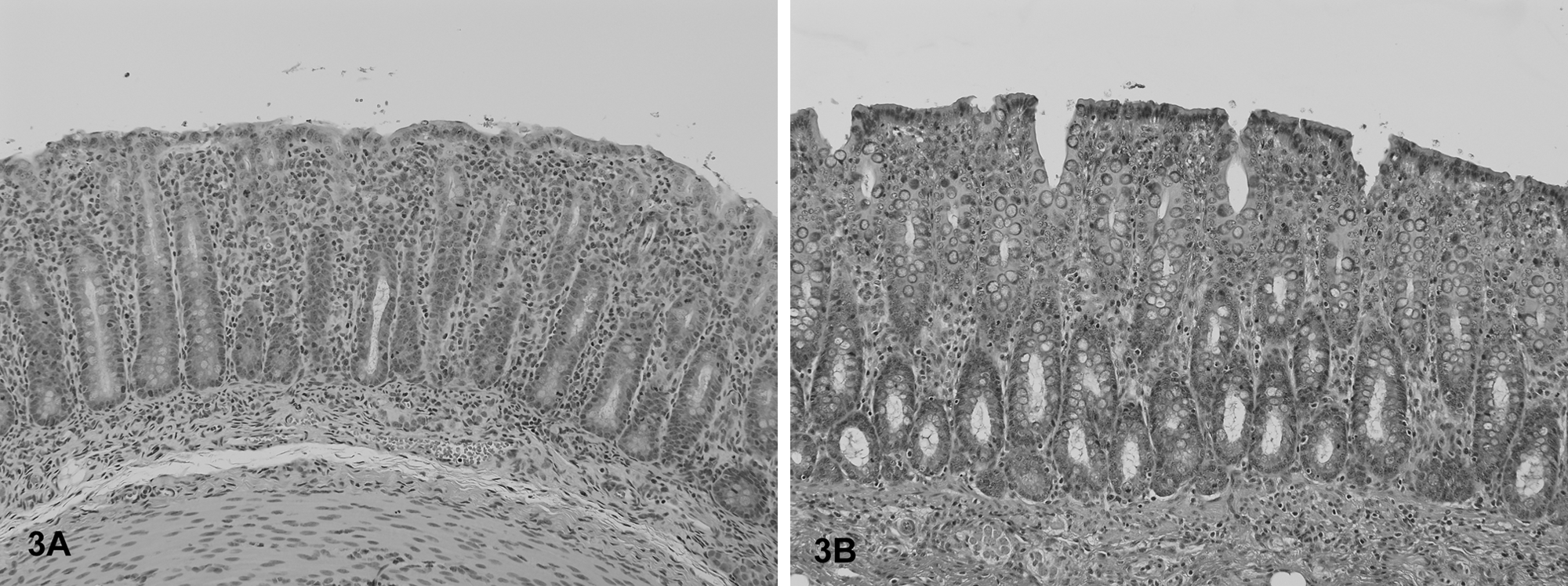
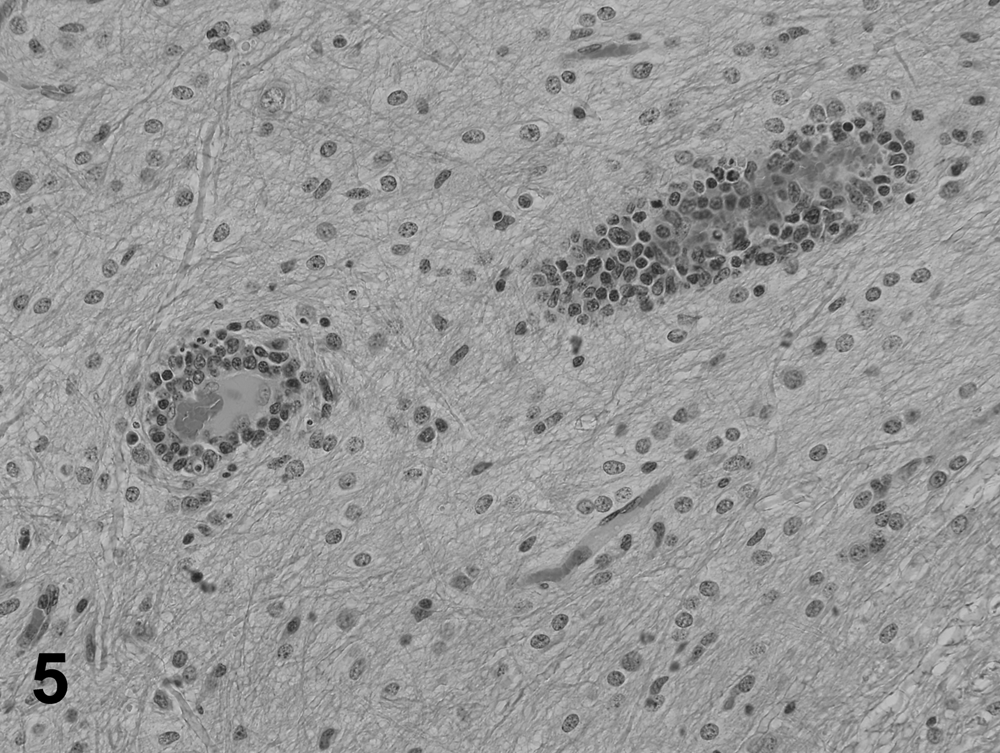
The most frequent histological lesions in PFTS-SICK pigs were thymic atrophy, superficial lymphoplasmacytic fundic gastritis, villous atrophy of the small intestine (duodenum, jejunum, and ileum), superficial colitis, lymphocytic and neutrophilic rhinitis, and mild nonsuppurative meningoencephalitis (Table 5). The frequency of these lesions was significantly higher in the PFTS-SICK than CTRL pigs (Table 5). In the thymus, the thickness of the cortex was severely reduced, yet the remnant of the medulla was still visible. In the stomach, the surface lining epithelial cells of the fundus were multifocally to diffusely attenuated, and mucus was lost in the cytoplasm. The glandular epithelial cells were occasionally necrotic. Moderate numbers of lymphocytes and plasma cells infiltrated the subepithelial lamina propria of the fundic stomach (Fig. 1). In fundus, no ganglionitis was observed. Unfortunately, pylorus was not examined in the current study, where ganglionitis, if present, will be more likely found. The small intestinal villi were moderately to severely shortened (Fig. 2) and covered by low cuboidal to occasionally squamous epithelial cells. Low to moderate numbers of different stages of coccidia were present in the small intestinal epithelium of 6 out of 18 (33.3%) PFST-SICK pigs (Table 2). The colonic epithelium was disorganized, hyperplastic, or attenuated. Increased numbers of lymphocytes and plasma cells were present in the colonic lamina propria (Fig. 3). In approximately half of the PFST-SICK pigs, coccobacilli were observed attaching to the colonic epithelium, leading to rounding, necrosis, and sloughing of the epithelial cells, morphologically consistent with attaching and effacing E. coli (AEEC) infection (Table 2). The nasal mucosa was infiltrated by moderate to large numbers of lymphocytes, plasma cells, and neutrophils. The epithelial cells of the submucosal glands were frequently necrotic and attenuated. Large basophilic intranuclear inclusion bodies were occasionally observed in the glandular epithelium in a small portion of PFTS-SICK pigs, consistent with SuHV-2 infection, known commonly as inclusion body rhinitis (Table 2, Fig. 4). Mild nonsuppurative meningoencephalitis, characterized by lymphocytic to histiocytic perivascular cuffing around the meningeal and parenchymal vessels (Fig. 5), was observed in 6 of the 13 PFTS-SICK pigs in which the brain was examined histologically. The frequency of nonsuppurative meningoencephalitis in PFTS-SICK pigs was significantly higher than in PFTS-HLTHY and CTRL pigs.
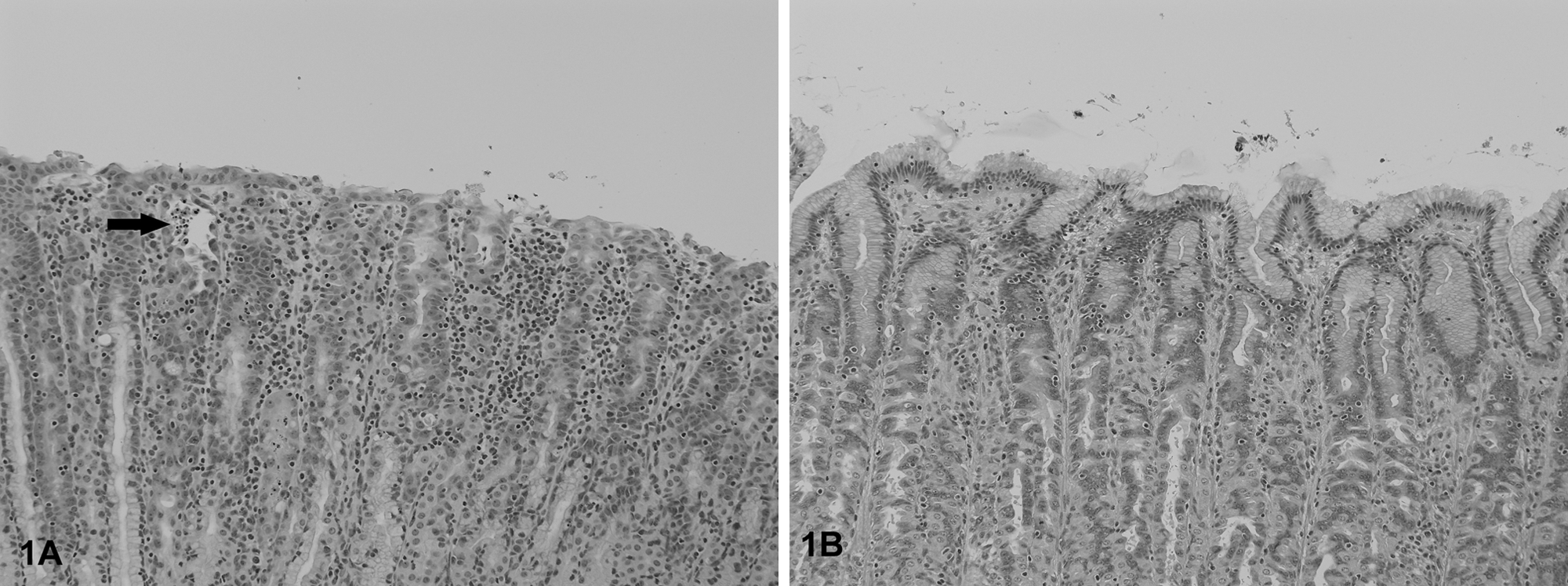
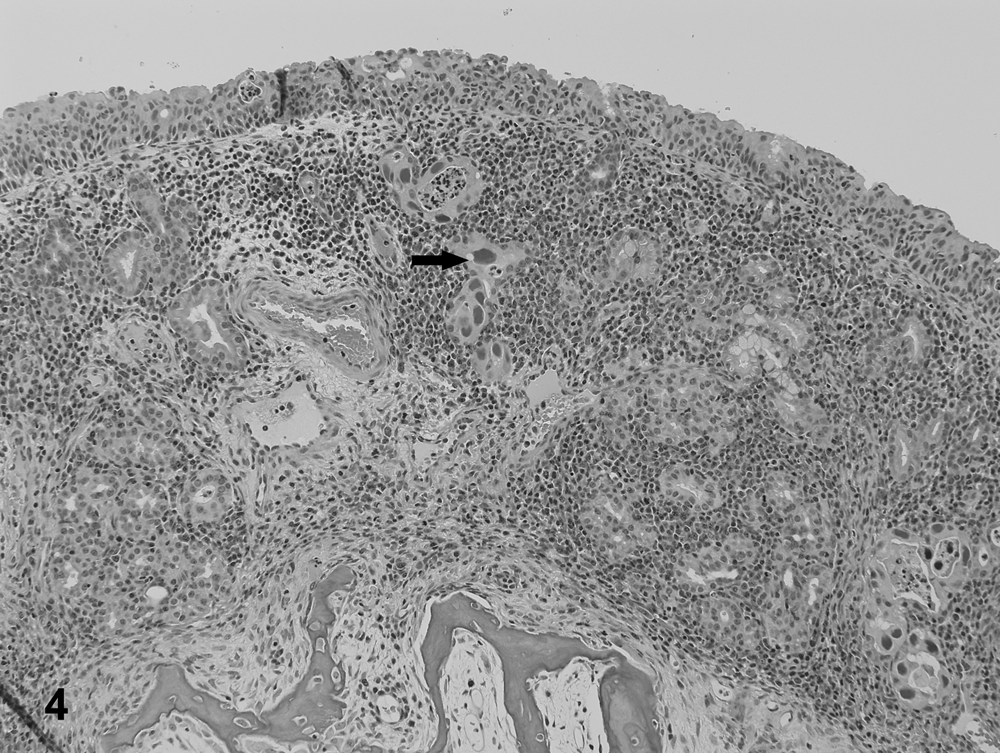
Nasal mucosa from a porcine periweaning failure-to-thrive syndrome–affected pig. Large numbers of inflammatory cells are infiltrating the lamina propria. Occasionally, the nasal glandular cells contain large, basophilic, intranuclear inclusion bodies (arrow), consistent with porcine cytomegalovirus.
Cerebellum of a porcine periweaning failure-to-thrive syndrome–affected pig. Two lymphocytic perivascular cuffing are shown in the white matter of the cerebellum.
Occasionally, other lesions were observed in the PFTS-SICK pigs, including various degrees of suppurative bronchopneumonia, lymphocytic nephritis, and fatty liver (Table 5). In the lung, bronchopneumonia was characterized by infiltration of neutrophils and consolidation of the parenchyma. Necrotizing bronchiolitis was not observed. The prevalence of bronchopneumonia in PFTS-SICK pigs was significantly higher than that in PFTS-HLTHY and CTRL pigs. No other significant lesions were observed in the tissues sampled.
Rhinitis was present in all PFTS-HLTHY pigs and was (16.7%) or was not (83.3%) associated with cytomegalovirus inclusion bodies (Table 5). Atrophic duodenitis (85.7%), jejunitis (14.3%), and superficial colitis (71.4%) were also present in the PFTS-HLTHY pigs (Table 5). The prevalence of rhinitis, atrophic duodenitis, and superficial colitis in PFTS-HLTHY pigs was significantly higher compared to CTRL, but not different from PFTS-SICK pigs (Table 5). One CTRL pig had mild rhinitis and 1 had colitis. No coccidia were observed in the intestines of PFTS-HLTHY pigs, but 1 single macrogamont was detected in 1 out of 8 (12.5%) CTRL pigs. No other histological lesions were observed in any PFTS-SICK or CTRL pigs.
Detection of bacteria
No biologically relevant bacteria were consistently cultured from PFTS-SICK pigs (Table 2). No F4-positive E. coli or Salmonella spp., or target DNA of B. hyodysenteriae and B. pilosicoli were detected from any pigs. Clostridium perfringens was cultured from the intestines of some PFTS-SICK (22%) and PFTS–HLTHY (29%) pigs, but not CTRL pigs. Two C. perfringens isolates from PFTS-SICK pigs were tested by PCR for toxin genes (alpha, beta1, beta2, and epsilon toxin genes). Both isolates were only positive for alpha toxin and classified as C. perfringens type A (data not shown). Pasteurella multocida, Streptococcus suis, Haemophilus parasuis, Bordetella bronchiseptica, and Streptococcus equisimilis were isolated once from different PFTS-SICK pigs (Table 2). There were no group differences in the frequency of the above bacteria (Table 2).
Detection of viruses
The viral detection results are summarized in Table 2. No PCV-2 antigen was detected in lymphoid tissue, stomach, or small and large intestine of any PFTS-SICK or PFTS-HLTHY pig by IHC. All PFTS-SICK and PFTS-HLTHY pigs tested negative for PRRSV, FLUAV, and ACoV-1. A low proportion of PFTS-SICK pigs were positive for RV-A in small intestine by IHC. Enteric calicivirus was detected in at least 1 pig in each group by PCR, and SuHV-2 was detected in all pigs except in 1 PFTS-SICK pig. Betacoronavirus 1 was present in the tonsils of one-third of the PFTS-SICK pigs, but not in any PFTS-HLTHY and CTRL pig. However, the lungs, kidneys, and small and large intestines were negative for BCoV-1 DNA. Further, when the brain stems (medulla oblongata) of the BCoV-1 tonsil–positive PFTS-SICK pigs were tested for BCoV-1 DNA by PCR, all were negative. All of the pigs were negative for TTV-1, and small numbers of pigs in each group were positive for TTV-2. Despite the identification of these viral agents, there were no statistical group differences in the detection frequencies among PFTS-SICK, PFTS-HLTHY, and CTRL pigs (Table 2).
Detection of parasites
Nonsporulated coccidia oocysts were detected in all 3 groups (Table 2). No other parasitic oocysts were observed by fecal flotation. No statistical differences in the detection frequencies of coccidia were identified among PFTS-SICK, PFTS-HLTHY, and CTRL pigs (Table 2).
Discussion
The current study aimed to describe the pathological changes in pigs affected by PFTS, and to identify common swine pathogens associated with, and possibly the cause of, PFTS. The most prevalent lesions in PFTS-SICK pigs were superficial lymphocytic fundic gastritis, atrophic enteritis, superficial colitis, lymphocytic and neutrophilic rhinitis, mild nonsuppurative meningoencephalitis, and thymic atrophy. The GI lesions, especially those present in the stomach, of the PFTS-SICK pigs may explain the clinical presentation of anorexia and wasting. Gastritis, enteritis, and colitis together can cause abdominal discomfort and loss of appetite, and thus feed refusal. However, it is also possible that the GI lesions were secondary to anorexia. Fasting in rats can cause small intestinal villous atrophy. 3 In pigs, Pittman et al. reported that jejunal atrophic enteritis was induced in 6 out of 6 pigs by 4 days of fasting that began on the day of weaning (Pittman JS, Moeser JA, Rovira A: 2011, Porcine peri-weaning failure to thrive syndrome (PFTS), Part II: gross lesions, histopathology and diagnostic analysis. In: Proceedings of the 42nd annual meeting of American Association of Swine Veterinarians, pp. 365–366.) In that study, lymphocytic gastritis was observed in 3 out of 6 fasted pigs. The same lesion however, was also seen in 3 out of 6 healthy pigs necropsied on the day of weaning (whether these pigs would be affected by PFTS was not known), 2 out of 6 healthy, and 2 out of 6 PFTS-affected pigs all on 4 days postweaning. By contrast, lymphocytic gastritis was not observed in pigs from a PFTS-free farm 4 days postweaning. The data suggest that in the PFTS-affected farm that Pittman studied, lymphocytic gastritis is a background lesion, regardless of treatment group. The same conclusion cannot be drawn in the current study, however, because superficial lymphocytic fundic gastritis was only observed in PFTS-SICK pigs and not in PFTS-HLTHY pigs. It should be noted that in the Pittman study, a detailed histological description of the lymphocytic gastritis is not available, thus comparison of this lesion to the gastric changes observed in the present study is not possible. The pathogenesis of the thymic atrophy observed in PFTS-SICK pigs is unknown in that it may either be a primary change associated with infection by an immunosuppressive pathogen (e.g., PCV-2 8 ), or secondary to prolonged sickness or anorexia. 4
Mild to severe suppurative bronchopneumonia was present in approximately 40% of the PFTS-SICK pigs. Although bronchopneumonia would have likely contributed to the clinical progression of the affected pigs, it cannot fully explain the clinical signs in all PFTS-SICK pigs. It is more likely that the pneumonia developed secondary to debilitation in some sick animals and was caused by opportunistic bacteria. Due to the difficulty in interpreting the biologic relevance of lesions observed in chronically debilitated pigs, it is suggested that diagnostic efforts focus on pigs in the early stages of illness. Thus, future investigations by the current authors will specifically target clinical pigs within 2 weeks of weaning.
A significant portion of PFTS-SICK pigs had very mild nonsuppurative meningoencephalitis, the cause of which was not determined by the current investigation. The significance of this lesion is uncertain. On the one hand, this may be a potential explanation for the chomping behavior, suspected to be centrally neurological in origin, which was observed in this PFTS-affected farm. On the other hand, inflammation in the histological sections examined, including 13 serial sections of brains from each of 3 PFTS-SICK pigs, was very mild, and may not be clinically relevant. Clinical neurological examination and additional histological examination of brain of additional PFTS-SICK pigs is underway and will help determine if the oral behavior and other clinical signs of PFTS are associated with neurological deficits.
It is noteworthy that certain lesions such as atrophic duodenitis, superficial colitis, and rhinitis were highly prevalent in both PFTS-SICK and PFTS-HLTHY pigs, and the frequencies of these lesions were not statistically different. The presence of these lesions in both groups leads to 3 possible interpretations: 1) atrophic duodenitis, superficial colitis, and rhinitis may be background lesions in pigs postweaning in this farm and not associated with PFTS; 2) these lesions may exist before or at weaning and increase the risk of PFTS; or 3) these lesions may exist before or at weaning and subsequently trigger PFTS in some but not all pigs. The data of the current investigation does not preferentially point to any of these 3 possibilities.
Several pathogens were only detected in PFTS-SICK pigs including RV-A, BCoV-1, C. perfringens type A, B. bronchiseptica, Streptococcus spp., H. parasuis, and P. multocida. It is the opinion of the authors that the clinical presentation of PFTS is not consistent with infection by these agents. Further, none of the above pathogens were detected more frequently (P > 0.05 for all) in PFTS-SICK compared to PFTS-HLTHY or CTRL pigs (Table 2). Thus, their causal role in PFTS is not supported by the results of the current study.
Rotavirus is a well-known cause of diarrhea and atrophic enteritis. Of the 4 rotavirus-positive pigs, 2 had very mild positive staining and were reported e as “suspicious.” The antibody against rotavirus used in the current investigation is polyclonal. Although the antibody can react with RV-A, whether it reacts with RV-B or RV-C is not known. Polymerase chain reaction typing of the rotavirus detected in PFTS-SICK pigs may help diagnosticians understand the role of rotavirus in PFTS. Rotavirus is highly prevalent in swineherds, especially in nursery pigs,6,13 and the clinical signs of PFTS are not consistent with rotavirus infection (i.e., lack of consistent diarrhea in PFTS-SICK pigs). The involvement of RV-B and RV-C in PFTS needs further clarification.
Betacoronavirus 1 can cause vomiting and wasting disease (VWD) in suckling pigs, can frequently be found in the brain stem and ganglions of the GI tract of infected pigs, and this distribution of BCoV-1 can explain the typical GI clinical signs. 1 Betacoronavirus 1, however, was not detected in the GI tract of any PFTS-SICK pig, nor in the brain stem (medulla oblongata) of PFTS-SICK pigs that were BCoV-1–positive in tonsil. Betacoronavirus 1 is found in tonsil of acutely infected pigs. 16 In the current study, however, the PFTS-SICK pigs were chronically affected at the time of necropsy. Thus, the presence of BCoV-1 in tonsil only and the absence of histological changes in the tonsil fail to explain clinical signs in PFTS-SICK pigs. Although the data from the current investigation does not definitely rule out BCoV-1 as the cause of PFTS, evidence for its causal role is insufficient in the authors’ opinion.
Attaching and effacing E. coli associated with superficial colitis was observed histologically in both PFTS-SICK and PFTS-HLTHY pigs, and PCR was employed to confirm the presence of AEEC DNA in affected colon. Surprisingly, AEEC DNA was detected in some pigs in all 3 groups, and group differences were not statistically significant. It has been reported that AEEC only causes mild disease in pigs and is usually associated with infection by other pathogens.10,11 It is possible that AEEC-associated colitis is a background lesion on this farm with no clinical significance. Alternatively, the underlying colitis associated with AEEC and/or other unidentified etiology could predispose pigs to PFTS.
Detection of PECV in all groups of pigs is consistent with the current understanding of the epidemiology and pathogenesis of PECV infection in swine. 20 Porcine enteric calicivirus only causes diarrhea and small intestinal villous atrophy in gnotobiotic piglets following experimental infections 5 and is not associated with diarrhea in naturally exposed piglets. 6 Accordingly, the role of PECV in PFTS is not determined but unlikely in the authors’ opinion.
Although coccidia oocysts (eggs) were identified in all groups, it was not possible to identify the genus and species by morphology. One Isospora spp. (I. suis) and several Eimeria spp. can infect pigs, but only I. suis can cause prominent disease. Isospora suis typically causes diarrhea and low mortality in 1–3-week-old pigs, 14 which does not match the clinical presentation of PFTS. Older pigs are more resistant to I. suis, 22 and the PFTS-SICK pigs were at the edge of the susceptible age. However, the histological presence of coccidia in the PFTS-SICK pigs was clearly associated with enteritis in some cases. Thus, further investigation is needed to determine the species and to determine whether the PFTS-SICK pigs were infected with a highly pathogenic strain or dose of coccidia. Moreover, it is necessary to determine whether I. suis/coccidia is a primary or secondary (i.e., sick pigs are more susceptible to the infection) pathogen.
Suid herpesvirus 2 is highly prevalent in suckling and nursery pigs and is not always associated with detrimental clinical signs. 7 The present study further confirmed the high prevalence of SuHV-2, as nearly all the pigs, regardless of group, were infected with SuHV-2. Sneezing is the usual manifestation of SuHV-2 infection, and mortality is usually low. 14 Rarely, systemic infection by the virus can cause death. 17 However, SuHV-2 inclusion bodies were not observed in organs outside the nasal cavity in any PFTS-SICK pigs. Thus, the role of SuHV-2 in the pathogenesis of PFTS is not clear but unlikely in the authors’ opinion.
In conclusion, the most frequently observed lesions in PFTS-affected pigs were superficial lymphocytic fundic gastritis, atrophic enteritis, superficial colitis, lymphocytic and neutrophilic rhinitis, nonsuppurative meningoencephalitis, and thymic atrophy. Although several pathogens were identified in PFTS-affected pigs, the current diagnostic investigation was not successful in identifying the definite cause of PFTS. There is a lack of compelling evidence to support the causal roles of the following pathogens in PFTS: PRRSV, PCV-2, FLUAV, ACoV-1, RV-A, SuHV-2, PECV, BCoV-1, C. perfringens, pathogenic E. coli, B. hyodysenteriae, B. pilosicoli, Bordetella spp., Streptococcus spp., H. parasuis, P. multocida, and coccidia.
Footnotes
Acknowledgements
The authors acknowledge the pathologists and technicians at the Prairie Diagnostic Services Inc. who assisted with various aspects of this work. The fecal flotation was technically supported by Brent Wagner, and the necropsies were assisted by Crissie Auckland.
Notes
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
This research was made possible with generous contributions from Saskatchewan Pork Development Board and Saskatchewan Agriculture and Food through the Agriculture Development Fund and Western College of Veterinary Medicine (WCVM) Disease Investigation Fund. Salary for Dr. Huang is provided by WCVM Interprovincial Graduate Fellowship Program.