
Abstract
A recently emerged H1N1 Influenza A virus (pandemic H1N1 (pH1N1)) with a Swine influenza virus (SIV) genetic background spread globally from human-to-human causing the first influenza virus pandemic of the 21st century. In a short period, reverse zoonotic cases in pigs followed by a widespread of the virus in the pig population were documented. The implementation of effective control strategies, rapid diagnosis, and differentiation of such virus from endemically circulating SIV in the various swine populations of the world is needed. To address the problem, a multiplex reverse transcription polymerase chain reaction assay utilizing a combination of the PB1, H1, and N1 primers that can rapidly and simultaneously subtype and screen for the presence of pH1N1 virus infection in Thai pigs was developed. The assay had 100% specificity and did not amplify genetic material from other subtypes of SIV, seasonal H1N1 human influenza (huH1N1) virus, highly pathogenic influenza H5N1 virus, and other important swine respiratory viral pathogens. The assay was able to both detect and subtype pH1N1 virus as low as 0.1–50% tissue culture infective doses/ml (TCID50/ml). The assay was used to screen 175 clinical samples obtained from SIV suspected cases, of which 6 samples were pH1N1 positive and were confirmed through virus isolation and whole genome sequencing. The results of the study suggested that the assay would be useful for the rapid diagnosis of pH1N1 in suspected Thai swineherds, where genetics of the endemically circulating SIV differ from the strains circulating in North American and European herds.
Influenza A virus (family Orthomyxoviridae, genus Influenzavirus A) infects and causes acute disease of the respiratory tract in many animal species including pigs and human beings. The virus is divided into 2 subtypes, according to the major surface viral antigens, hemagglutinin (HA) and neuraminidase (NA) proteins. 20 Virus isolates within each subtype are diverse, with changing genetic and antigenic composition at the HA and NA regions, allowing for antigenically novel influenza A viruses to potentially cause epidemics and/or pandemics when introduced into an immunologically naïve host populations. 3 Swine influenza virus (SIV), a member of influenza A viruses, remains one of the major problems in pig-raising countries throughout the world. Clinical manifestations caused by SIV in pigs are similar to human influenza, including coughing, sneezing, labored abdominal breathing, and anorexia.7,8 Currently, there are 3 well-established SIV subtypes (H1N1, H3N2, and H1N2) circulating in pigs. 17 The genetic makeup of H1N1 SIV in North American pigs prior to 1998 was designated “Classical swine flu” because all 8 gene segments were from swine origin. 17 Since the emergence of the H3N2 subtype in 1998, contemporary H1N1 viruses in the North American swine population evolved to a triple reassortment of swine, avian, and human viruses with antigenically distinct character of the HA protein. 19 In contrast, H1N1 virus introduced into European pigs in 1979 contains wholly avian genes and are antigenically distinct from the classical swine H1N1 viruses. To date, European H1N1 isolates contain H1 segments from both human and avian lineages. 22 In Thailand, H1N1 SIV isolates are separated into 2 clusters, which differ from the North American and European H1N1 viruses, but contain HA and NA surface genes, respectively, from the classical and Eurasian swine origin. The internal genes of the first cluster include the polymerases (PB1, PB2, and PA), matrix (M), nucleoprotein (NP), and nonstructural protein (NS) of Eurasian swine origin, while in the second cluster, the NS genes are of classical swine origin. 15
The emergence of the novel swine origin H1N1 Influenza A virus (pandemic H1N1 [pH1N1]) in April of 2009 initiated the first human pandemic of the 21st century. 9 Implementation of effective control strategies, rapid diagnosis, and identification of newly emerged influenza A viruses in both the human and swine populations were required. Many conventional and real-time reverse transcription (RT) polymerase chain reaction (PCR) protocols for the detection of pH1N1 virus in the human population have been published.5,11,21 On the one hand, rapid diagnosis of pH1N1 in human beings through such methods is easily achieved, as pH1N1 gene composition can be discriminated from the currently circulating seasonal human H1N1 viruses. On the other hand, it is difficult to differentiate pH1N1 virus from various endemic swine H1N1 (sH1N1) viruses currently circulating in pigs in different regions of the world, particularly through previously published RT-PCR or real-time RT-PCR assays.1,12 Aside from subtyping the virus, sequencing of all 8 gene segments from the suspected swine samples is required to accurately differentiate pH1N1 from endemic H1N1 SIV. Based on available genetic data, a multiplex RT-PCR assay that uses combinations of specific primers targeting the PB1, H1, and N1 genes and can rapidly and simultaneously subtype and screen for the presence of pH1N1 virus infection in Thai pigs without the need for whole genome sequencing was developed in the current study. The multiplex RT-PCR was validated against the H1 (including H1N1 and H1N2 subtypes) SIVs, seasonal, and the pH1N1 subtype that currently circulates in both the pig and human population.
A panel of reference influenza A viruses and common swine respiratory viral pathogens were used in the current study for RT-PCR assay validation (Table 1). All influenza A viruses were propagated in Madin–Darby canine kidney (MDCK) cells as described previously. 7 The SIV titers were determined by calculation of a tissue culture infective dose (TCID50)/ml based on the Reed–Muench method using influenza type A protein-specific immunocytochemistry staining of infected MDCK cells. 7 Multiple aliquots of stock viruses were stored at 80°C for use throughout the study. Viral RNA was extracted from each virus isolate using a commercial RNA kit a according to the manufacturer’s recommendations. The primers constructed in the present study were designed by using the ClustalX program (version 1.8) b and the OLIGO primer design software (version 9.1) c and are shown in Table 2. The PB1-P primer specific for pH1N1 virus was designed based on the PB1 gene of reference viruses indicated in Table 1. Two additional primer pairs, H1-A and N1-SEu, specific to pH1N1 virus were designed for subtyping swine H1N1 viruses. The N1-SEu primer was also designed based on N1 gene sequences of Eurasian swine (avian-like) viruses for differentiating North American swine H1N1 viruses from pH1N1 viruses. 6 The primer sets were selected for amplification in an easily discriminating multiplex format of the 429-, 704-, and 516-bp PCR products, corresponding to PB1, H1, and N1 genes, respectively.
Single-step multiplex reverse transcription polymerase chain reaction assay for detection and differentiation of the 2009 H1N1 Influenza A virus pandemic in Thai swine populations: influenza A viruses used in the current study for assay validation.
Multiplex RT-PCR was performed in a single-step reaction using a commercial RT-PCR system. d Three sets of primers, PB1-P, H1-A, and N1-SEu, specific for PB1, H1, and N1, respectively, of pH1N1 virus were used at a final concentration of 0.5 µM each. A combination of 3.0 µl of RNA sample with a reaction mixture containing 12.5 µl of kit-supplied master mix, d 5 U of Avian myeloblastosis virus (AMV) reverse transcriptase, and RNase-free water was used in a final volume of 25 µl (Table 2). Cycling conditions of multiplex RT-PCR assay included a reverse transcription step at 48°C for 45 min, initial denaturation step at 94°C for 3 min, 40 cycles of denaturation (94°C for 30 sec), annealing (55°C for 30 sec), and extension (72°C for 1 min), and a final extension at 72°C for 10 min. A total of 10 µl of PCR product was analyzed on a 1.5% agarose gel e stained with ethidium bromide and visualized by ultraviolet transilluminator.
Primers constructed for the single-step multiplex reverse transcription polymerase chain reaction assay for detection and differentiation of the 2009 H1N1 Influenza A virus pandemic in Thai swine populations.
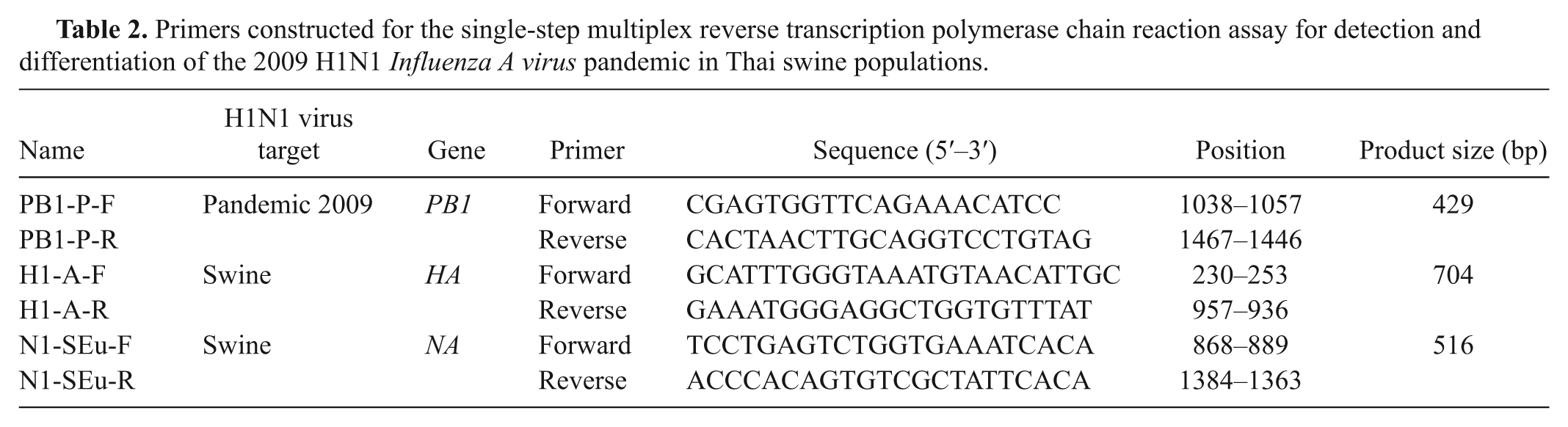
The specificity of the assay was determined by testing the primers with common swine respiratory viral pathogens, including Porcine reproductive and respiratory syndrome virus (PRRSV), Porcine circovirus-2 (PCV2), Suid herpesvirus 1 (SuHV-1), and Classical swine fever virus (CSFV), pH1N1, SIV subtypes H1N1, H1N2, and H3N2, highly pathogenic H5N1, and seasonal H1N1 human influenza (huH1N1). Because RNA from North American sH1N1 viruses was not available, the N1-SEu primer was evaluated online using the Basic Local Alignment Search Tool (BLAST) in GenBank and with multiple sequencing alignments of North American swine H1N1 viruses, and it was also tested with Eurasian sH1N1 viruses to confirm N1-SEu primer specificity. The sensitivity test of the assay was determined by using RNA extracted from 10-fold serial dilutions of stock virus (H1N1, A/Swine/Thailand/CU-RA4/2009) starting at 105–0.01 TCID50/ml.
Results showed that the multiplex RT-PCR assay can be used to subtype the sH1N1 viruses and that it can be applied to detect and differentiate pH1N1 viruses (isolated from pigs and human beings) from other SIVs, including sH1N1 and huH1N1 viruses, simultaneously (Fig. 1). The multiplex RT-PCR primers had 100% specificity to H1N1 SIV, as genetic material from all other SIV subtypes and other swine respiratory viral pathogens was not amplified (Fig. 2A). Online BLAST search and the multiple sequence alignment indicated that N1-SEu primer was specific to Eurasian avian-like swine viruses. The multiplex RT-PCR assay using PB1-, H1-, and N1-specific primers was able to both detect and subtype pH1N1 virus as low as 0.1 TCID50/ml (Fig. 2B). The PB1 primer specific to pH1N1 was able to detect 0.001 TCID50/ml of pH1N1 (data not shown).
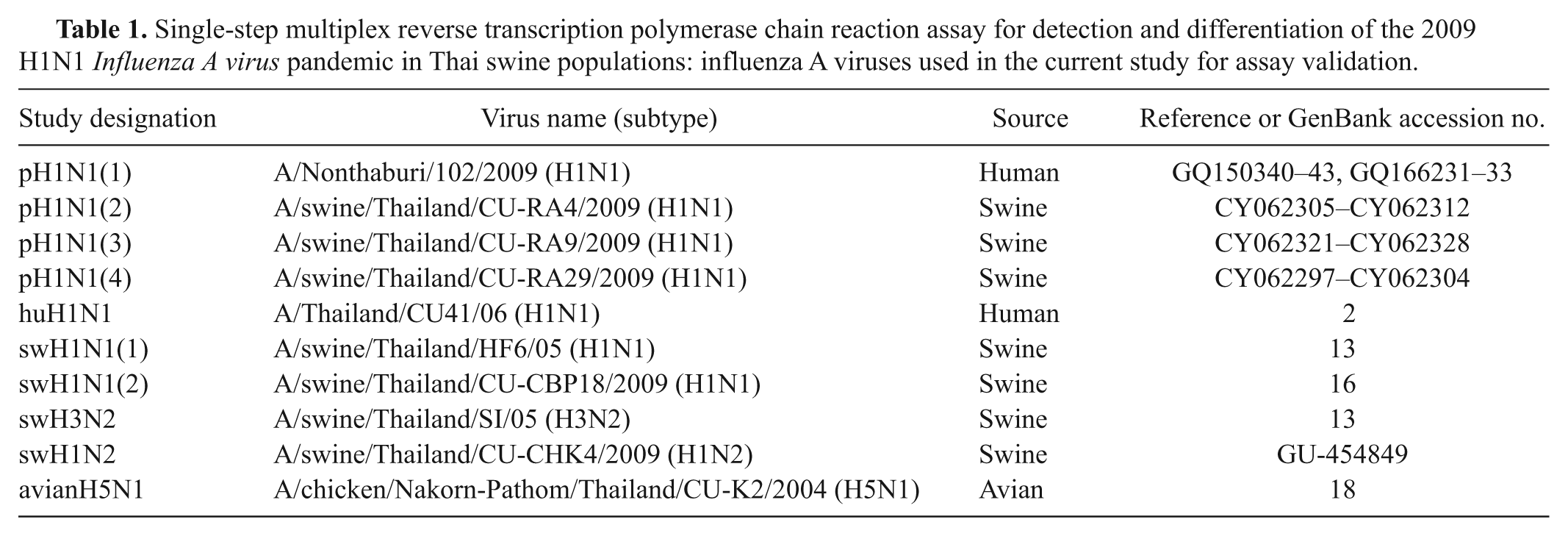
Detection, differentiation, and subtyping of H1N1 influenza A viruses by multiplex reverse transcription polymerase chain reaction (RT-PCR) assay. Lane 1: pandemic (p)H1N1(1) virus; lane 2: pH1N1(2) virus; lane 3: pH1N1(3) virus; lane 4: pH1N1(4) virus; lane 5: swine (s)H1N1(1) virus; lane 6: sH1N1 (2) virus; lane 7: negative control; lane M: 100-bp plus ladder. Arrows: expected RT-PCR products for H1 (704 bp), N1 (516 bp), and PB1 (429 bp) genes.
One hundred and seventy-five nasal swab samples obtained from SIV suspected cases during December 26, 2009, and March 9, 2010, submitted to Chulalongkorn University Emerging Infectious Disease in Animals (CU-EIDA) and Chulalongkorn University Veterinary Diagnostic Laboratory (CU-VDL) were tested directly using the assay reported herein for H1N1 and pH1N1 screening test. Isolation of SIV was performed on the same samples, and selected H1N1 SIV–positive samples were further characterized by whole genome sequencing to confirm pH1N1-positive samples, as described previously. 14 Results indicated that 15 samples were SIV H1N1 positive (6 nasal swab samples were positive for pH1N1 and 9 nasal swabs were positive for sH1N1). Virus isolation on all samples was positive for SIV on the same 15 nasal swab samples. Whole genome sequencing on H1N1-positive samples confirmed that 6 nasal swabs were pH1N1 positive while the remaining nasal swabs were positive for endemic Thai H1N1 SIV.
The initial identification and confirmation of pH1N1 viruses were performed by screening with RT-PCR and/or real time RT-PCR assay followed by virus isolation 4 and DNA sequencing, 10 which are respectively labor-intensive and not readily available in laboratories in remote areas of many countries, particularly Thailand. In order to facilitate rapid diagnosis, the multiplex RT-PCR assay targeting the PB1 gene of pH1N1 virus, H1 gene of all influenza A subtypes, and N1 gene of Eurasian avian-like swine viruses was developed for simultaneous detection, subtyping, and differentiation of pH1N1 viruses from endemic circulating Thai SIV. Such an assay is rapid and utilizes a conventional PCR machine; thus, it is cost saving, requires minimal expertise, and is available in local laboratories around the country. The PB1-P primer was sensitive and specific for the detection and differentiation of pH1N1 virus from Thailand and European swine influenza isolates. The H1-A primer can detect and subtype all H1 of SIVs, including pH1N1 virus. The N1-SEu primer can detect and subtype all N1 of Eurasian swine origin, including pH1N1 virus while differentiating pH1N1 virus from North American swine influenza isolates. In addition, the assay was able to detect pH1N1 in human beings, and can differentiate pH1N1 from seasonal huH1N1 viruses. Available data on SIV and pH1N1 in many Southeast Asian countries is limited. However, this area of the world also raises pigs for both consumption and as backyard farm animals where they can closely interact with other animal species including human beings. The development of such a basic tool is, thus, important for the rapid diagnostic of SIV in neighboring countries to provide important, yet missing, preliminary data in this area, which can help the prevention and control of the disease in pigs on a global scale.
Footnotes
Acknowledgements
The authors thank Ms. S. Wannaratana, Ms. D. Prakairungnumtip, Dr. T. Pohuang, Ms. N. Charoenvisal, Ms. P. Ninvilai, Ms. K. Rungprassert, and all staff from the Virology Unit, Faculty of Veterinary Science, Chulalongkorn University for their assistance in laboratory work. The authors would like to acknowledge Prof. Y. Poovorawan and Dr. S. Payungporn from the Center of Excellence in Clinical Virology, Faculty of Medicine, Chulalongkorn University for their contribution of the human pH1N1 virus and huH1N1 virus. The authors also thank Ms. P. Hirsch for reviewing the brief research report. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIH. The authors are also grateful to Chulalongkorn University Centenary Academic Development Project for supporting facilities to the Emerging and Re-emerging Infectious Disease in Animals, Research.
a.
NucleoSpin® RNA Virus, Macherey-Nagel, Düren, Germany.
b.
CLUSTAL X program (version 1.8), Conway Institute UCD Dublin, Ireland.
c.
OLIGO primer design software (version 9.1), Molecular Biology Insights Inc., Cascade, CO.
d.
AccessQuick RT-PCR System, Promega Corp., Madison, WI.
e.
TopVision™ LE GQ Agarose, Fermentus Inc., Glen Burnie, MD.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The current work has been funded in whole with federal funds from the National Institute of Allergy and Infectious Disease, National Institute of Health, Department of Health and Human Services, under contract no. HHSN266200700007c. The authors thank the Emerging Health Risk Cluster, Rachadapiseksompoch Endowment Fund, Chulalongkorn University.