
Abstract
The microanatomy of healthy skin from 12 different body sites was investigated in 14 alpacas (Vicugna pacos). The microanatomy of alpaca skin is typical of domestic animal skin in general, and closely resembles that from llamas.
Keywords
Introduction
Alpacas (Vicugna pacos, formerly Lama pacos) are growing in popularity, and often present with skin disorders that provide a diagnostic and therapeutic challenge for the veterinary clinician. 2,3 Skin biopsy specimens from alpacas are commonly submitted to the Animal Health Diagnostic Center at Cornell University (Ithaca, New York); submissions have increased from 2 in 1997 to 17 in 2006.
After an English language literature search, only 2 articles addressing selected aspects of the microanatomy of healthy alpaca skin were found. In 1 article, investigators described the microanatomy of skin biopsy specimens from the thorax of 3 alpacas. 10 In the other article, investigators described the histologic features of the epitrichial sweat glands in skin biopsy specimens from 10 alpacas. 6 For the most part, histologic information gathered from healthy skin of llamas is extrapolated to alpacas. 1,4,7 This may or may not be appropriate. The objective of the present study was to characterize the histologic features of healthy skin from alpacas.
Materials and methods
Skin biopsy specimens were collected from 14 alpacas necropsied at the Cornell University College of Veterinary Medicine. Alpacas ranged from 3 months to 15 years of age, included 9 females and 5 males, and included animals with white, brown, or black hair coats (Table 1). The breeds (Huacaya or Suri) were not recorded. No alpaca had a history of skin disease, and no macroscopic skin lesions were present at necropsy. All alpacas died or were euthanized because of acute diseases. The skin samples were collected within 24 hr after death. Haired areas were carefully clipped to avoid disturbing the skin surface. Skin biopsy specimens were taken with a 6-mm biopsy punch from 12 different sites on the body: muzzle, lips, cheek, pinnae, lateral neck, mid-back, rump, lateral thorax, ventral thorax, lateral thigh, pastern, and interdigital space. Skin specimens were fixed in 10% neutral buffered formalin, embedded in paraffin, processed routinely, and stained with hematoxylin and eosin (HE) and acid orcein Giemsa (AOG), which stains mast cell granules and elastin fibers.
In addition to routine histologic examination, the following features were specifically examined: skin thickness, epidermal thickness, stratum corneum thickness, number of cell layers in the stratum granulosum, number of cell layers in the stratum spinosum, ratio of melanocytes-to-keratinocytes in the stratum basale, and number of perivascular mast cells.
The thickness of the skin, epidermis, and stratum corneum was measured using a computerized light micro-scope a and morphometric software. b Representative areas of each location were measured for each of the 14 alpacas, and the numbers were averaged for each body site. It should be noted that much of the stratum corneum is lost during routine histologic processing, making measurement of this layer in paraffin-embedded sections unreliable. 8
Results
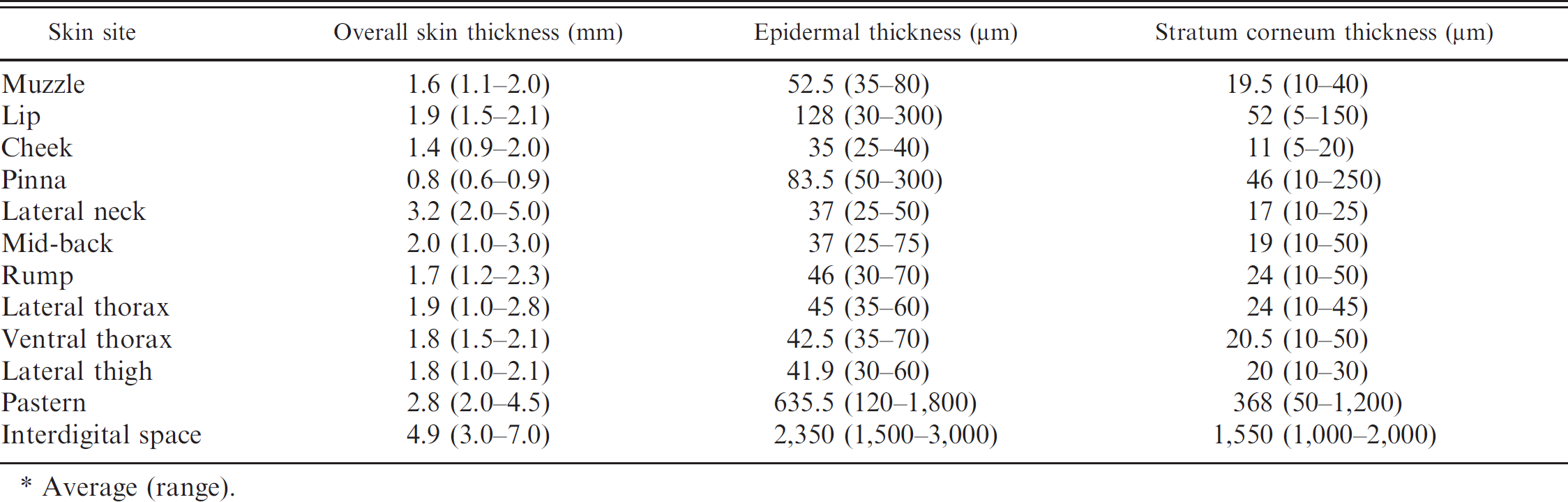
The average skin thickness varied from 0.8 mm to 4.9 mm (Table 2); the thickest skin was from the interdigital space (4.9 mm), neck (3.2 mm), and pastern (2.8 mm). The average epidermal thickness varied from 35 μm to 2,350 μm (Table 2; Figs. 1–3); the thickest epidermis was from the interdigital space (2,350 μm) and pastern (635.5 μm). The average stratum corneum thickness varied from 11 μm to 1,550 μm (Table 1); the thickest stratum corneum was from the interdigital space (1,550 μm) and pastern (368 μm).
The epidermis at all 12 body sites was composed of a stratum basale, stratum spinosum, stratum granulosum, and stratum corneum. A stratum lucidum was not seen. The stratum spinosum consisted of 2–3-cell layers in most body sites (Fig. 2). The stratum spinosum was thicker in the lip, pastern, and interdigital space (Fig. 3), consisting of up to 5-, 9-, and 15-cell layers, respectively. The stratum granulosum was discontinuous and, when present, consisted of 1-cell layer in most body sites (Fig. 2). In the interdigital space, lip, and pastern, the stratum granulosum was consistently present and ranged from 2- to 6-, 2- to 3-, and 1- to 3-cell layers in thickness, respectively (Fig. 3). The stratum corneum in most body sites had a basket weave configuration (Fig. 1), except in the lip, pastern, and interdigital space where the configuration was compact and laminated (Fig. 3). Rete ridges were only observed in the lip, pastern, and interdigital space (Fig. 3).
Age, sex, and fleece color data on 14 alpacas from the current study.
Numerous melanocytes were present in the stratum basale (1 melanocyte per every 2–10 basal keratinocytes) of the muzzle, lip, cheek, pinna, pastern, and interdigital space. In these sites, melanin granules were present in all layers of the epidermis (Fig. 2). In other body sites, few melanocytes were seen, and melanin granules were rarely seen. Merkel cells were not seen. The basement membrane zone was usually imperceptible.
Measurements of parameters in healthy skin from alpacas in the current study. *
Average (range).
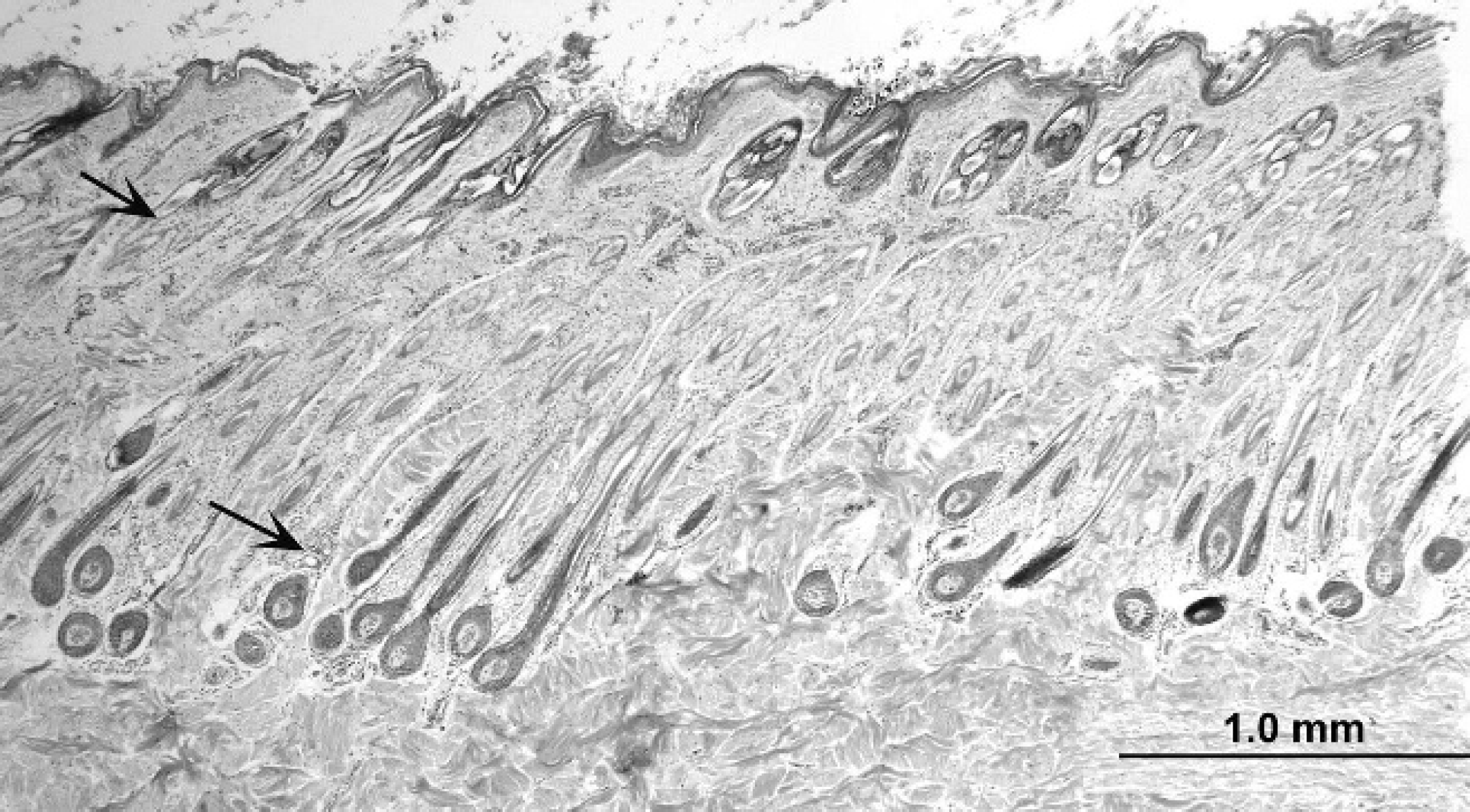
Alpaca (Vicugna pacos); skin, lateral neck. Note thin epidermis and compound hair follicle arrangement. Sebaceous glands (upper arrow) and epitrichial sweat glands (lower arrow) are small. Collagen fibers in the superficial dermis are smaller than those in the deep dermis. Hematoxylin and eosin. Bar = 1 mm.
In general, collagen bundles were smaller in the superficial dermis than in the middle and deep dermis (Fig. 1). Oval fibrocyte nuclei were more abundant in the superficial than in the deep dermis. The interdigital space was composed of larger collagen bundles at all levels (Fig. 3). Skeletal muscle bundles were present in the lip. Elastic fibers were not visible in HE-stained sections, but were found to be sparsely scattered throughout the dermis in AOG-stained sections. The density of elastic fibers was increased in sections from the lip and rump, especially around adnexae. No lymphatic vessels were recognized.
Sections from all skin sites had small-to-mediumsized blood vessel plexuses that extended from the superficial to the deep dermis (Fig. 2). The vessel walls were thick and had large round-to-oval endothelial nuclei. Erythrocytes were seen within vessel lumens. Mononuclear cells containing eosinophilic cytoplasmic granules (“globular leukocytes” 1 ) were rarely seen: 0–1 cell per vascular plexus, in 1 of every 5–6 plexuses. The pastern and cheek had large blood vessel plexuses that were seen throughout the dermis. In all AOG-stained sections, there were 0–3 mast cells per 400× microscopic field in perivascular areas.
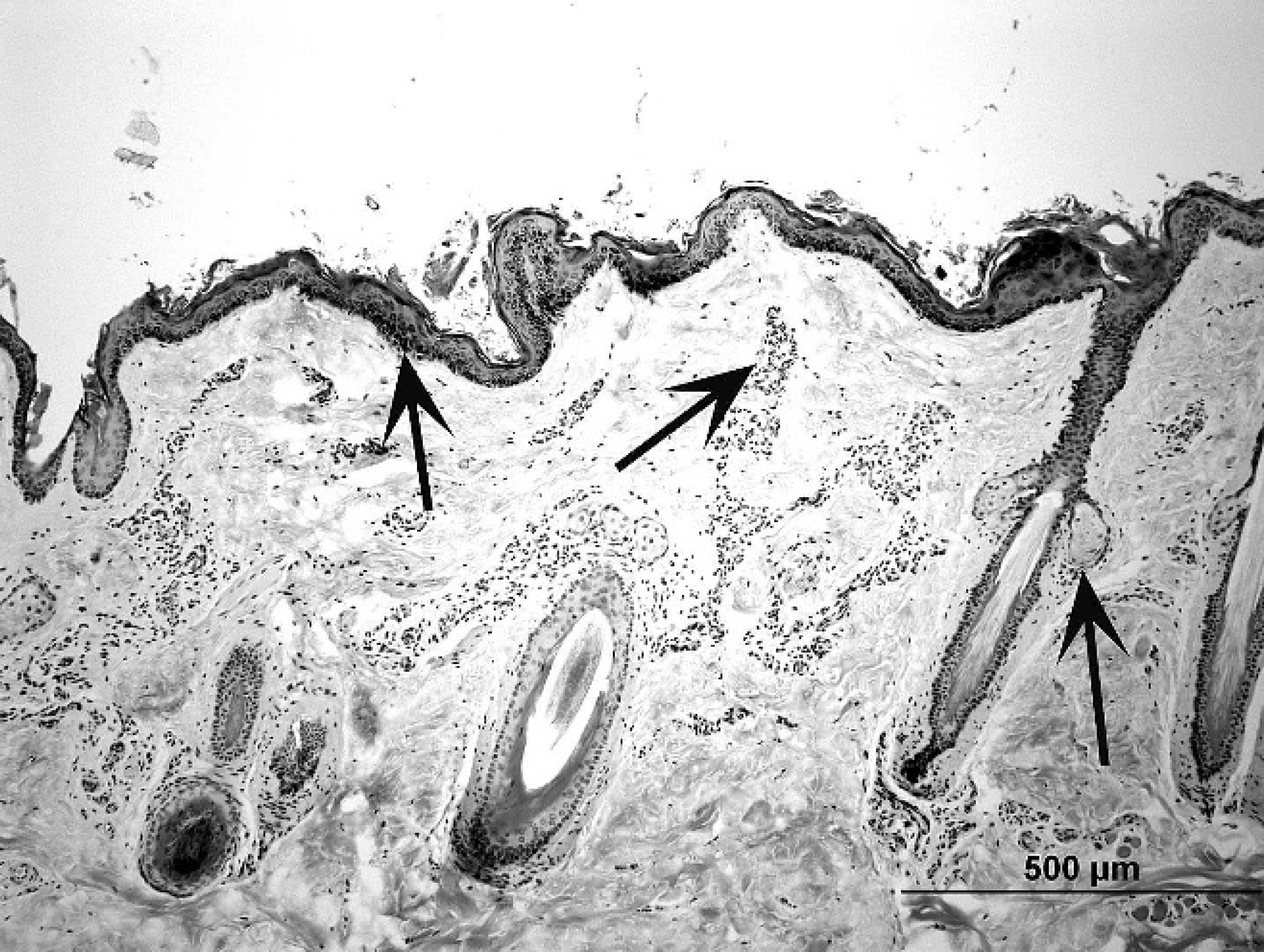
Alpaca (Vicugna pacos); skin, lip. Note compact stratum corneum, prominent melanin granules in the epidermis (left arrow), simple hair follicle arrangement, and prominent vascular plexuses (middle arrow). Small sebaceous glands are connected by short ducts to hair follicle infundibulum (right arrow). Hematoxylin and eosin. Bar = 500 μm.
Compared with the other locations studied, hair follicle density was relatively low in the muzzle, lip, cheek, pinna, and pastern. Compound hair follicle anatomy characterized most body sites (Fig. 1). Primary follicles were surrounded by multiple smaller secondary follicles, which all fused together near the entrance of the sebaceous ducts and shared a common follicular infundibulum. Larger simple hair follicles were seen in the muzzle, lip, and pastern (Fig. 2). Hair follicles were not seen in the interdigital space. Typical hair follicle anatomy was present: infundibulum, isthmus, and inferior segment. Anagen hair follicles were characterized by a dermal hair papilla, hair matrix, inner root sheath (composed of the inner root sheath cuticle, Huxley layer, Henle layer, and trichohyalin granules), outer root sheath, and basement membrane zone. Subjectively, there was a mix of anagen and telogen hair follicles with no apparent difference in the various body sites. Hair follicles were oriented obliquely to the skin surface. Sinus hair follicles were seen only in specimens from the muzzle (7/14 alpacas), and were as described for llamas. 1
Well-developed arrector pili muscles were associated with larger hair follicles in locations where the fleece was long and dense (lateral neck, mid-back, rump, lateral thorax, lateral thigh). The arrector pili muscles were smaller in locations where the hair coat was short.
Sebaceous glands were present at all body sites, except for the interdigital space. In most areas, only 1 or 2 small sebaceous glands were associated with each hair follicle (Figs. 1, 2). Sebaceous glands were sometimes difficult to find in the lateral and ventral thoracic specimens. Sebaceous ducts were short and wide, and composed of 2-cell layers of nonkeratinized, stratified squamous epithelium.
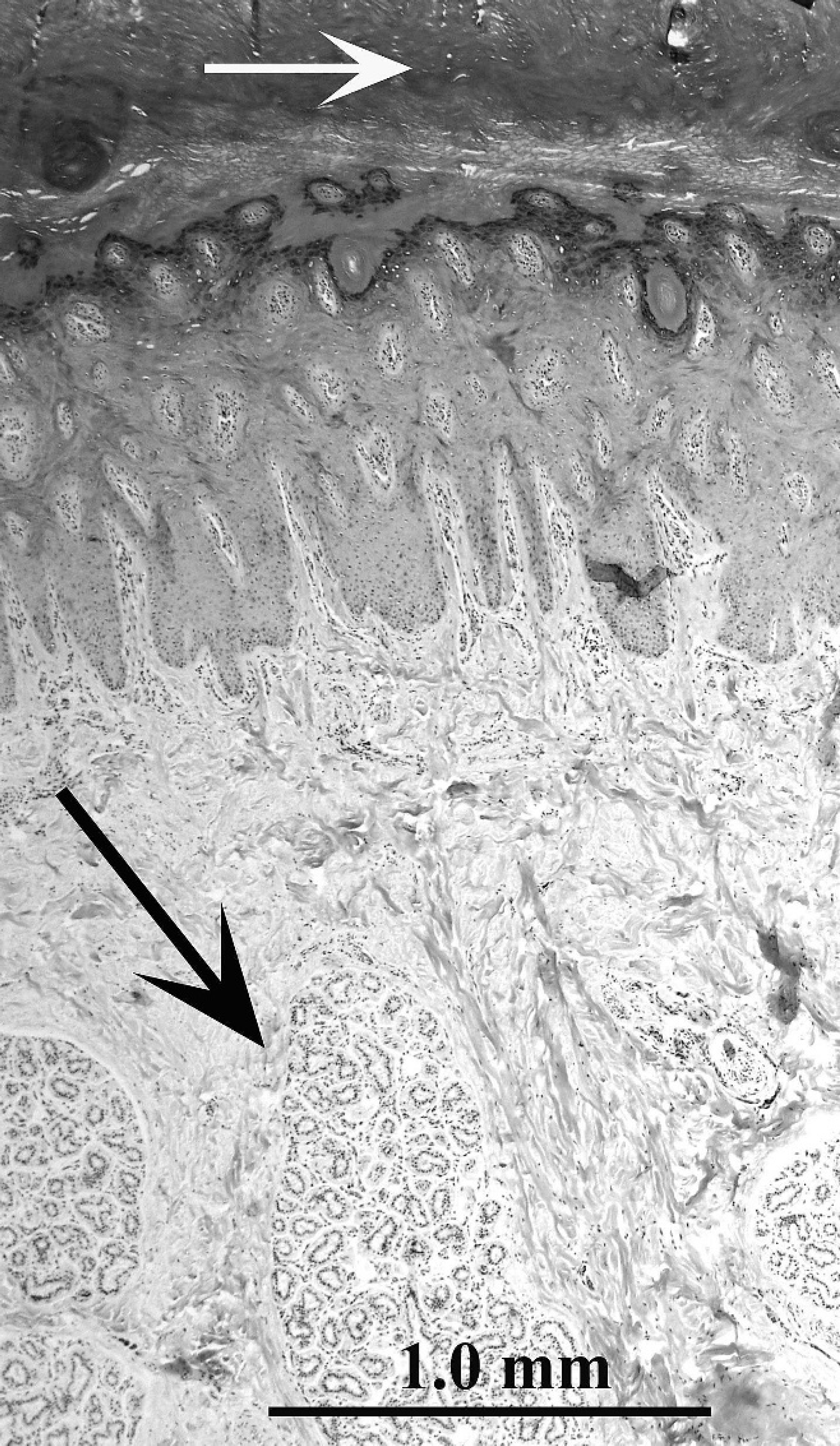
Alpaca (Vicugna pacos); skin, interdigital space. Note thick epidermis with thick, compact stratum corneum (white arrow) and rete ridge formation. Prominent atrichial sweat glands (“interdigital glands”; black arrow) are seen. Hematoxylin and eosin. Bar = 1 mm.
Epitrichial sweat glands were small (Fig. 1) and were present at all body sites, except for the lip and the interdigital space. The glandular portion of the gland was tubular and often coiled, with flattened-to-columnar secretory epithelium. The sweat duct was coiled or straight and consisted of 2 layers of flattened or cuboidal epithelial cells. Large atrichial sweat glands (“interdigital glands”) were present in the interdigital space (Fig. 3). Epitrichial and atrichial sweat glands rarely contained luminal secretions. The subcutis generally consisted of loose connective tissue containing small amounts of adipose tissue. There were no apparent differences in any of the histologic parameters evaluated in respect to age, sex, or fleece color. Unfortunately, breed data had not been recorded.
Discussion
The increasing popularity of alpacas, their propensity for developing skin disease, and the resultant upsurge in skin biopsy specimen submissions to veterinary diagnostic laboratories necessitates the accumulation of information specific for the micro-anatomy of healthy skin from these animals. Previous usage of the llama as the template for normal skin microanatomy in the alpaca required validation.
The histologic appearance of healthy skin from alpacas is typical of that of mammalian and domestic animal skin in general. 5,8,9 The current study reveals that the histology of healthy skin from alpacas is, indeed, very similar to that reported for llamas, 1 and is in agreement with the findings of previous small studies. 6,10
Typical mast cells could not be identified in HE-stained sections of alpaca and llama skin. 1,10 However, AOG-stained sections revealed 0–3 mast cells per 400× microscopic field in perivascular areas, which is in agreement with a previous small study. 10 Special stains were not performed in the llama study. Elastin fibers, unless abnormal, were not visible in HE-stained skin sections. 1,5,8,9 In the present study, elastin fibers were easily identified in AOG-stained sections of alpaca skin. In summary, the current small pilot study indicates that the microanatomy of the healthy skin from alpacas closely resembles that from llamas. Diagnostic pathologists may find this information useful in the evaluation of skin biopsy specimens from alpacas with various dermatoses.
Footnotes
a.
Olympus VANOX AH3, Olympus Imaging America Inc., Center Valley, PA.
b.
Image-Pro® Plus, Media Cybernetics Inc., Bethesda, MD.