
Abstract
Brucella abortus is a bacterium of brucellosis causing abortion in cattle. The diagnosis of bovine brucellosis mainly relies on serologic tests using smooth lipopolysaccharide (S-LPS) from B. abortus. However, the usefulness of this method is limited by false-positive reactions due to cross-reaction with other Gram-negative bacteria. In the present study, the eryC gene encoding B. abortus D-erythrulose 1-phosphate dehydrogenase, which is involved in the erythritol metabolism in virulent B. abortus strain but is absent from a B. abortus vaccine strain (S19), was cloned. Recombinant EryC was expressed and purified for the evaluation as a diagnostic reagent for bovine brucellosis. Other B. abortus proteins, Omp16, PP26, and CP39 were also purified and their seroreactivities were compared. Recombinant EryC, Omp16, PP26, and PP39 were all reactive to B. abortus-positive serum. The specificity of recombinant Omp 16, PP26, CP39, and EryC, were shown to be approximately 98%, whereas that of B. abortus whole cell lysates was shown to be 95%. The sensitivity of Omp16, PP26, CP39, and EryC were 10%, 51%, 64%, and 43%, respectively, whereas that of B. abortus whole cell lysates was 53%. These results suggested that B. abortus EryC would be a potential reagent for diagnosis for bovine brucellosis as a single protein antigen.
Introduction
Brucella abortus (order Rhizobiales, family Brucellaceae), the etiologic agent of bovine brucellosis, is facultative intracellular Gram-negative bacteria. 5 Bovine brucellosis primarily affects cattle, and causes abortion and decreases reproductive efficacy, leading to economic losses. 23 Brucella abortus is also able to infect human beings, resulting in a public health problem, especially in developing countries. 23
Bovine brucellosis control programs mainly rely on individual tests and eradication by slaughter. Therefore, an early and correct diagnosis is essential for the treatment of bovine brucellosis. Current diagnosis of bovine brucellosis is mainly based on the detection of antibodies against B. abortus. 42 Thus, various serologic tests are used to diagnose B. abortus-infected animals as early as possible after infection. 1,18 Most serologic tests, including the standard tube agglutination test (TAT), are based on the smooth lipopoly-saccharide (S-LPS) of B. abortus as an antigen. 21
Although the S-LPS is widely used, as it is a highly immunogenic and dominant antigen, it has a critical defect in that it cross-reacts with other Gram-negative bacteria, such as Yersinia enterocolitica O:9, 15 Escherichia coli O:116/O157, 25 Salmonella urbana, 26 Francisella tularensis, 2 Vibrio cholerae, 17 Pseudomonas maltophilia, 8 and Pasteurella multocida. 11 These cross-reactions generate false-positive results, thus impeding accurate diagnosis of bovine brucellosis. 41
For the purpose of discriminating infected and vaccinated animals, the B. abortus RB51 vaccine strain, which expresses weak O-antigen of LPS, has been practiced. 37 However, possibly serologic tests using the S-LPS could not distinguish between B. abortus infection and the B. abortus S19 vaccine strain. This is another significant impediment in diagnosing bovine brucellosis in countries where vaccination using B. abortus S19 is practiced. 27 To alleviate this problem, a few B. abortus proteins have been evaluated as alternate diagnostic reagents for bovine brucellosis. 9,13,14,20,30,39 These include the outer membrane protein 16 kDa (Omp16), 4 the periplasmic protein 26 kDa (PP26), 30 the cytoplasmic protein 39 kDa (CP39), 9 and brucellin. 19 However, Omp16 showed weak cross-reactivity with phylogenetically related bacteria such as Ochrobactrum anthropi and Phyllobacterium rubiacearum 6 (also known as Phyllobacterium myrsinacearum), and CP39 showed relatively weak serologic responses. 9 Therefore, novel B. abortus–specific antigens that are able to induce high specificity and sensitivity for bovine brucellosis are needed. 7
Brucella abortus has a tropism for reproductive organs and colonizes in the placenta and fetus of pregnant cattle, where B. abortus uses the carbon source erythritol, 22 which accumulates in the reproductive organs during the later phases of gestation. 31,34 There are 4 open reading frames (ORFs) in the ery operon of B. abortus that are involved in erythritol metabolism. The eryA gene encodes putative erythritol kinase, the eryB gene encodes an erythritol phosphate dehydrogenase, the eryC gene encodes a D-erythrulose 1-phosphate dehydrogenase (EryC), and the eryD gene encodes a regulator of ery operon. 32 The b . abortus S19 vaccine strain is unable to use erythritol because the eryC gene is truncated. 35,36 Therefore, EryC may constitute a B. abortus target antigen for diagnostic tests, as it would distinguish the individuals infected with virulent B. abortus strain from B. abortus S19 vaccinated individuals.
In the current study, the B. abortus eryC gene was eloned, expressed, and purified. Seroreactivity of recombinant EryC, as a single protein antigen, was validated as a potential diagnostic reagent for bovine brucellosis using enzyme-linked immunosorbent assay (ELISA). Brucella abortus Omp16, PP26, and CP39 were also expressed and purified, and their seroreactivities were compared.
Materials and methods
Bacterial strains and culture conditions
Escherichia coli strains (Top10 and M15) a were used for cloning and expression of B. abortus genes, respectively, and were cultured on lysogeny broth (LB) media. The virulent B. abortus S2308 strain was obtained from the National Veterinary Research & Quarantine Service (Anyang, Korea) and was grown in tryptic soy broth (TSB) b supplemented with 5% bovine serum albumin (BSA) c at 37°C.
Serum samples
A total of 300 serum samples from cattle (100 TAT-positive and 200 TAT-negative) were provided by the Gyeonggi-do Veterinary Service (Suwon, Korea). The TAT-positive serum samples (n= 100) were used as positive controls for bovine brucellosis in the validation of B. abortus proteins. A group of randomly selected TAT-negative serum samples (n = 43) was used as negative controls for recombinant Omp16, PP26, and CP39, and the other group of TAT-negative serum samples (n = 93) was used as negative controls for recombinant EryC. Brucella abortus whole cell lysates were also used as ELISA antigen for bovine brucellosis. Control B. abortus–positive and B. abortus–negative serum samples were provided by the U.S. Department of Agriculture (USDA; Washington, DC) and were used for validation of recombinant B. abortus proteins by Western blot analysis.
PCR amplification and cloning of omp16, pp26, cp39, and eryC
The sequences of B. abortus S2308 omp16, pp26, cp39, and eryC genes were obtained from GenBank, d and were amplified by polymerase chain reaction (PCR) using oligonucleotide primers harboring appropriate restriction enzyme sites (Table. 1). The PCR-amplified genes were ligated into the pT7 blue vector, e and their sequences were confirmed.
Expression and purification of recombinant Omp16, PP26, CP39, and EryC
All 4 genes were ligated into the pQE30 expression vector f after digestion with corresponding restriction enzymes, affording pQE30::omp16, pQE30::pp26, pQE30::cp39, and pQE30::eryC. These constructs were then used to transform E. coli Top10, and then amplified using a commercial kit. f The expression constructs (pQE30::omp16, pQE30::pp26, pQE30::cp39, and pQE30::eryC) were used to transform E. coli M15 creating M15[pQE30::omp16], M15[pQE30::pp26], M15[pQE30::cp39], and M15[pQE30::eryC]. Cells were grown to an optical density (OD) of 0.6 at a wavelength of 600 nm (OD6600) in LB media containing 100 μg/ml ampicillin and 50 μg/ml kanamycin, treated with 0.5 mM of isopropy1 beta-D-thiogalactoside (IPTG) at 37°C for 4 more hr, recovered by centrifugation, and resuspended in binding buffer (6.0 M guanidine HC1, 0.1 M sodium phosphate, 0.01 M Tris-HC1 at pH 8.0). The mixture was rocked for 1 hr at room temperature and centrifuged, and the supernatant was collected. A high-yield, homogenous preparation of B. abortus proteins were obtained using a nickel–nitrilotriacetic acid (NiNTA) resin, according to the standard procedures described by the manufacturer. g Briefly, the supernatants were added to the equilibrated Ni-NTA agarose in a 1:10 volume ratio. The bead slurry was then washed with 10 volumes of 50 mM sodium phosphate, 300 mM NaC1, 10% glycerol, and 20 mM imidazole (pH 8.0). Recombinant Omp16, PP26, CP39, and EryC were eluted with 300 mM or 500 mM imidazole in 50 mM sodium phosphate, 300 mM NaC1, and 10% glycerol (pH 6.0). The eluted proteins were dialyzed with phosphate buffered saline (PBS; pH 7.4) for 48 hr at 4°C.
Western blot analysis
The recombinant B. abortus proteins were analyzed by a discontinuous sodium dodecy1 sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and the expression of the proteins were visualized by staining with Coomassie Brilliant Blue 250R. Proteins resolved by SDS-PAGE were electrophoretically transferred onto a nitrocellulose membrane for Western blot analysis. Membranes were incubated in PBS containing 0.05% Tween 20 and 5% normal goat serum (PBST-NGS) at room temperature for 1 hr and then incubated with a 1:300 dilution of B. abortus–positive and B. abortus–negative sera, provided from USDA, for 1 hr. The membranes were washed 4 times and subsequently treated with horseradish peroxidase (HRP)-conjugated goat antibovine immunoglobulin G (IgG) antibodies h at a 1:10,000 dilution for 1 hr at room temperature. Protein binding was detected using a commercial immunoblot system. 1
Oligonucleotide primers for cloning Brucella abortus genes.*
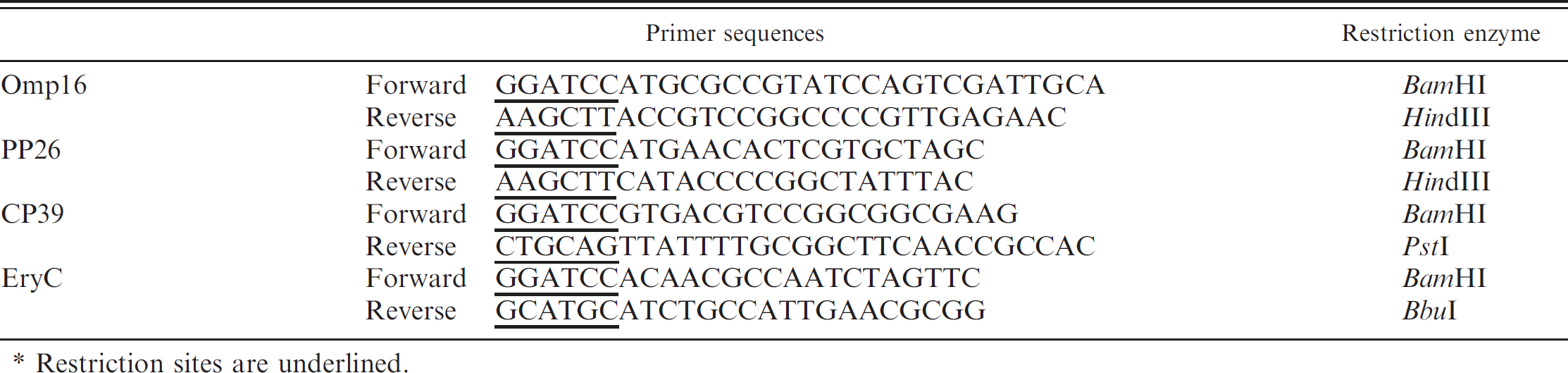
Restriction sites are underlined.
Whole cell lysate preparation
Brucella abortus cells were cultured for 3–4 days in TSB supplemented with 5% BSA and harvested by centrifugation at 4,000 rcf for 10 min. To eliminate the residual media, the recovered bacteria were washed with PBS and heat inactivated at 120°C for 15 min after adjusting the number of bacteria at McFarland 3.0 (9 × 108/ml). 16 After centrifugation, the pellet was lyophilized and dissolved in appropriate buffer for further study.
Indirect enzyme-linked immunosorbent assays
The ELISAs were performed to evaluate the reactivity of recombinant B. abortus proteins as previously described. 28 Ninety-six–well enzyme immunoassay (EIA) plates j were coated overnight with 1.5 g/ml Omp16, 1.0 μg/ml PP26, 2.0 μg/ml CP39, or 1.5 μg/ml EryC, or with 2 μg/ml B. abortus whole cell lysates diluted in 100 μl of polycarbonate buffer (pH 9.6) at 4°C. Plates were then washed and blocked with PBST-NGS. Serum samples, diluted (1:300) in PBST-NGS, were added to the wells and incubated for 1 hr at 37°C. The bound antibodies were detected with HRP-conjugated goat antibovine IgG h (1:20,000 dilution). After washing 6 times with PBST, an enzyme substrate solution containing O-phenylenediamine c and hydrogen peroxidase was added to the plates, and the plates were incubated for 15 min in the dark. The reaction was stopped by the addition of 2.5 N H2SO4. The absorbance was read at 490 nm using an automatic ELISA plate reader. k Each serum sample was assayed in duplicate, and the mean absorbance of carbonate buffer control wells was subtracted for the mean absorbance of B. abortus antigen-coated wells. The cutoff was defined as the mean absorbance value plus 3 standard deviations of the optical density (OD) values measured in the negative control samples, as previously described. 29 The TAT-negative serum samples were included as negative controls. Specificities and sensitivities of ELISA results using the recombinant B. abortus proteins were calculated as previously described. 12
Statistical analysis
Statistical significances of the specificities and sensitivities for each protein antigen were determined by the analysis of variance test using the GraphPad Prism (version 4.0) program, 1 and differences were considered significant when P values were <0.05.
Results
Cloning, expression, and purification of B. abortus omp16, pp26, cp39, and eryC
The PCR-amplified omp16, pp26, cp39, and eryC genes were ligated into the pT7 blue vector, subcloned into the pQE30 vector after enzyme digestion, and expressed in E. coli M15 cells. All 4 purified recombinant Omp16, PP26, CP39, and EryC proteins were analyzed by SDS-PAGE, and the gels were stained with Coomassie Brilliant Blue 250R (Fig. 1A). The molecular masses of Omp16, PP26, and CP39 were approximately 16, 26, and 39 kDa, respectively. Two protein bands were observed in the lane of purified EryC (Fig. 1A). The size of the upper band in SDS-PAGE gel was approximately 36 kDa, corresponding to the expected molecular mass of EryC. The unexpected lower band was shown at the position of approximately 26 kDa in the lane.
Western blot analysis of recombinant Omp16, PP26, CP39 and EryC
The identification of purified recombinant B. abortus Omp16, PP26, CP39, and EryC were confirmed by Western blot analysis using B. abortus–positive or B. abortus–negative sera. All 4 B. abortus proteins were reactive to B. abortus–positive serum (Fig. 1B), but not reactive to B. abortus–negative serum (data not shown). Recombinant Omp16 and CP39 reacted strongly to B. abortus–positive serum, whereas PP26 did relatively low. Only the upper EryC protein band reacted to B. abortus–positive serum, suggesting that the lower EryC protein band contains no antigenic epitope for generating corresponding antibody in cattle. These results suggest that all 4 recombinant B. abortus proteins used in the current study are reactive to anti–B. abortus sera.
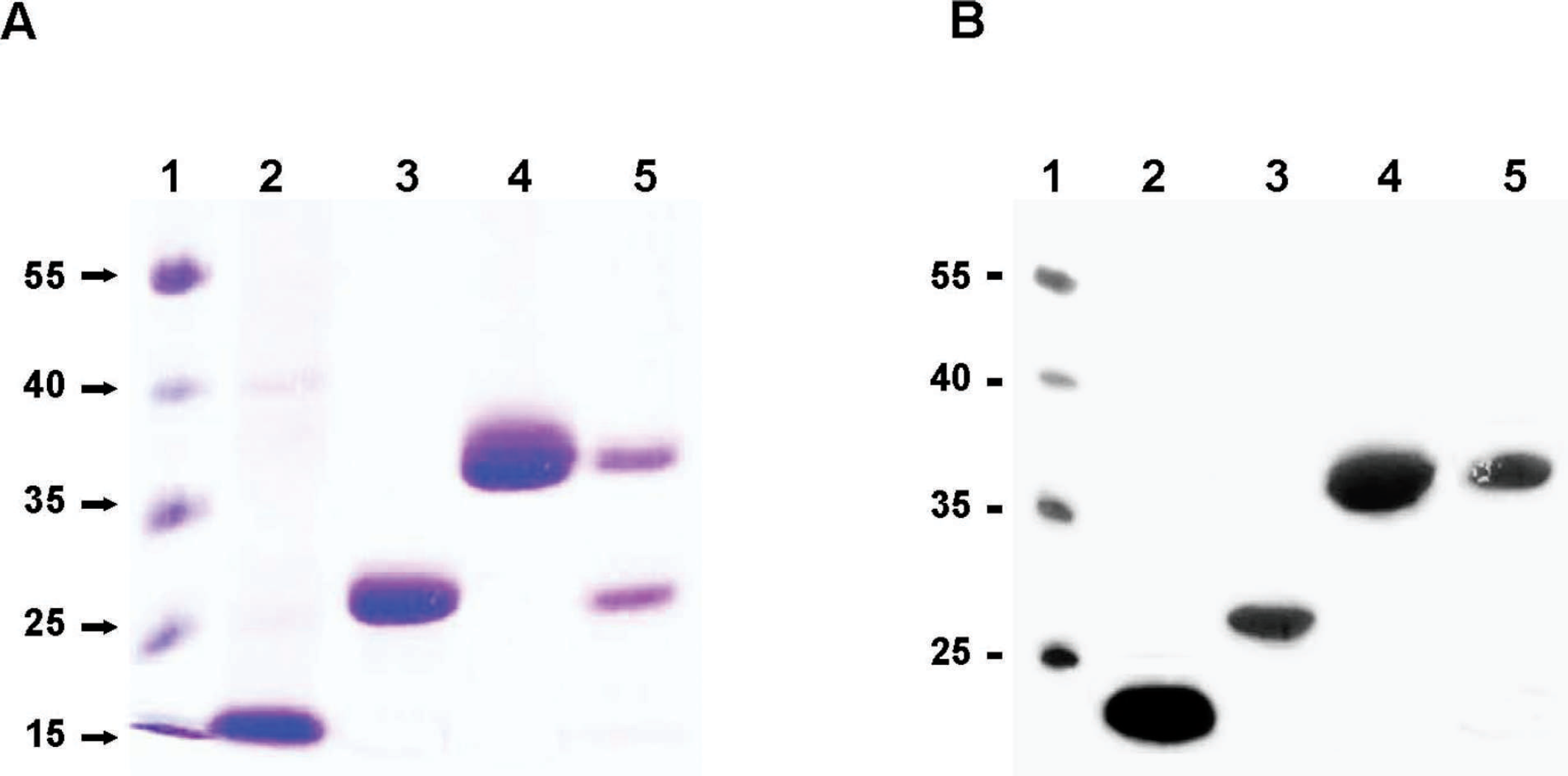
Sodium dodecy1 sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis and Western blot analysis of recombinant Brucella abortus proteins. A, SDS-PAGE analysis of recombinant B. abortus proteins. Lane 1: molecular weight marker; lane 2: Omp16; lane 3: PP26; lane 4: CP39; and lane 5: EryC. Recombinant B. abortus proteins in SDS-PAGE gels were stained with Coomassie Brilliant Blue 250R. B, Western blot analysis of the purified proteins was performed using B. abortus–positive serum samples.
Determination of seroreactivity of recombinant Omp16, PP26, CP39, and EryC using indirect ELISA
The seroreactivity of recombinant Omp16, PP26, CP39, and EryC was determined using an indirect ELISA. A total of 300 serum samples (100 TAT-positive and 200 TAT-negative) were used as sources of B. abortus antibody. The 300 serum samples were prediagnosed using TAT, the standard test of bovine brucellosis in Korea. The mean absorbance values for TAT-negative serum samples were measured to be 0.15, 0.14, 0.05, and 0.12 when recombinant Omp16, PP26, CP39, and EryC were used as antigens, respectively. The mean absorbance value of B. abortus whole cell lysates for TAT-negative serum samples was measured to be 0.20 (Fig. 2). The mean absorbance value of recombinant CP39 for TAT-negative serum samples was significantly lower than those of other proteins and of B. abortus whole cell lysates (P> <p 0.001). The mean absorbance value of recombinant EryC for TAT-negative serum samples was significantly lower than that of B. abortus whole cell lysates (P < 0.05). The mean absorbance values for TAT-positive serum samples were measured to be 0.34, 1.06, 0.36, 0.69, and 0.83 when recombinant Omp16, PP26, CP39, and EryC and B. abortus whole cell lysates were used as antigens, respectively. The mean absorbance value of recombinant PP26 for TAT-positive serum samples was significantly higher than those of the other proteins (P < 0.001) and that of B. abortus whole cell lysates (P < 0.05). The mean absorbance value of recombinant EryC for TAT-positive serum samples was higher than that of Omp16 (P < 0.001) or CP39 (P < 0.001). The sensitivities for recombinant Omp16, PP26, CP39, and EryC and for B. abortus whole cell lysates against TAT-positive serum samples were calculated to be 10%, 51%, 64%, 43%, and 53%, respectively (Table. 2). Although the sensitivity of recombinant CP39 was highest, the overall absorbance values of positive or negative samples were relatively lower than those obtained when other proteins were used as antigens. The sensitivity of the recombinant PP26 was comparable to that of B. abortus whole cell lysates. The sensitivity of recombinant EryC was considerably comparable to that of PP26 or B. abortus whole cell lysates. The specificities of recombinant Omp16, PP26, CP39, and EryC were calculated to be 97.7%, 97.7%, 97.7%, and 97.8%, respectively, whereas the specificity of B. abortus whole cell lysates was calculated to be 94.6%. These results indicate that recombinant PP26 and EryC react strongly toward bovine brucellosis serum samples and could serve as single diagnostic protein reagents for bovine brucellosis.
Discussion
Most serologic tests for bovine brucellosis are based on the detection of antibodies reacting to the B. abortus S-LPS antigen. Although the S-LPS antigen is highly immunogenic, it shares epitopes with other bacterial LPS antigens, possibly leading to false-positive reactions. The identification of new B. abortus–specific protein antigens as diagnostic reagents is needed to improve the quality of diagnosis of bovine brucellosis. Although the outer membrane proteins, 4,5 periplasmic proteins, 18,30,33 or cytoplasmic proteins 9,13 of B. abortus have been explored as a diagnostic reagent for bovine brucellosis, the serologic test using them is far from ideal.
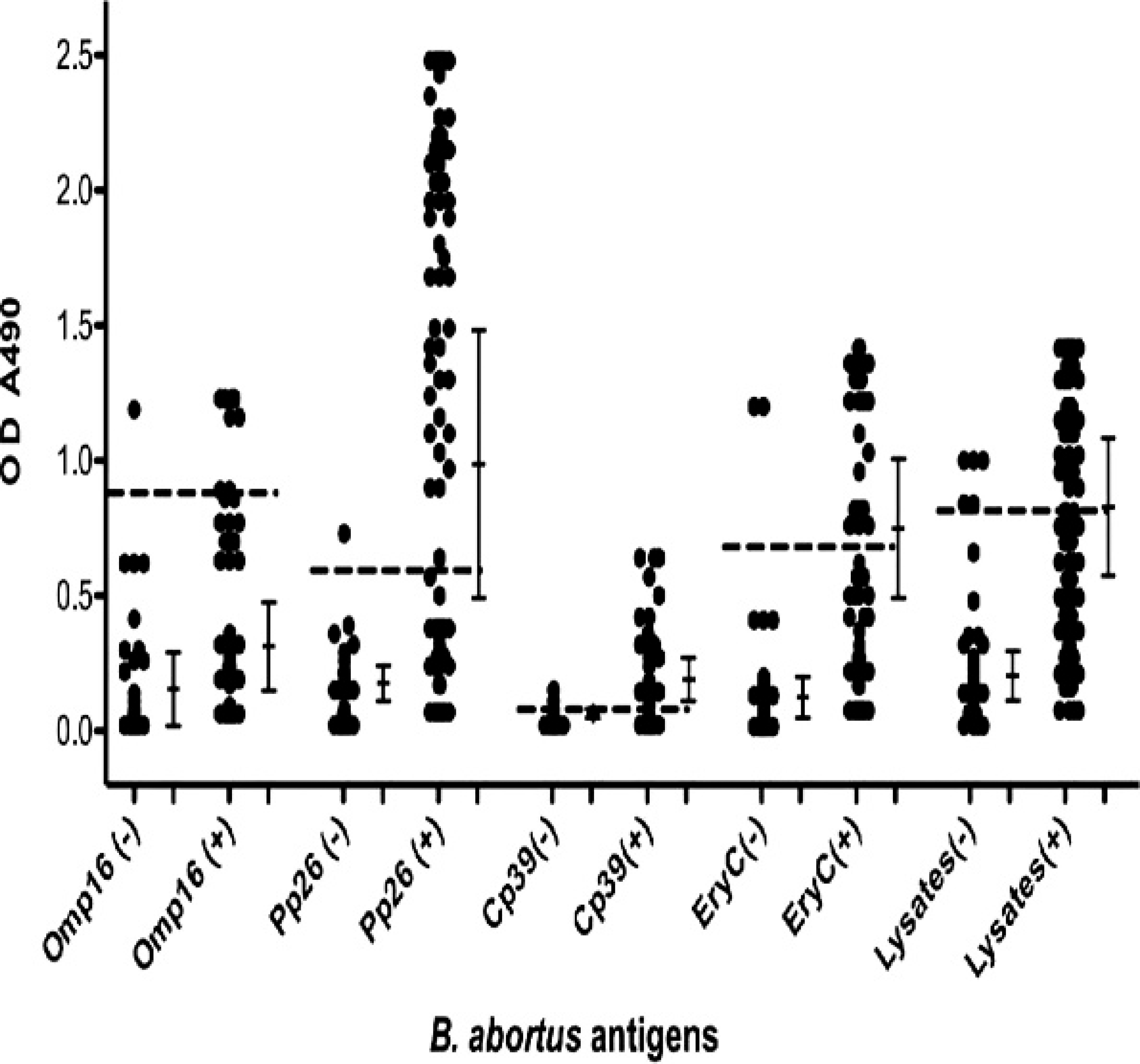
Seroreactivity of recombinant Omp16, PP26, CP39, and EryC, and of Brucella abortus whole cell lysates, for bovine brucellosis. Tube agglutination test (TAT)-positive serum samples (n – 100) were used as positive controls for bovine brucellosis, and TAT-negative serum samples (n = 43 or 93) were used as negative controls to evaluate the seroreactivity of recombinant B. abortus proteins used in the current study using an indirect enzyme-linked immunosorbent assay with 1.5 μg/ml Omp16, 1.0 μg/ml PP26, 2.0 μg/ml CP39, or 1.5 μg/ml EryC, or with 2 μg/ml B. abortus whole cell lysates. Vertical bars represent mean ± SD and dotted lines represent cutoff values (0.83, 0.54, 0.14, 0.66, and 0.80 for Omp16, PP26, CP39, and EryC, and for B. abortus whole cell lysates, respectively). OD = optical density.
In the present study, the seroreactivity of a novel B. abortus protein EryC, using an indirect ELISA with TAT-positive and TAT-negative serum samples, was determined. Among B. abortus proteins previously reported, Omp16 reacted strongly to B. abortus– positive cattle serum shown by Western blot analysis, but the sensitivity was relatively low (10%). The results obtained herein are consistent with previous reports that B. abortus Omp16 was immunogenic, but its sensitivity was low due to cross-reactivity with phylogenetically related other bacteria. 7,40 Recombinant CP39 generated low absorbance values in an indirect ELISA. These results are consistent with a report performed previously, 9 which demonstrated that the seroreactivity of 39-kDa cytoplasmic protein was relatively weak. Although recombinant CP39 had the highest sensitivity (64%), it may not be a good diagnostic reagent for bovine brucellosis because its seroreactivity was indistinguishable between TAT-positive and TAT-negative serum samples. Recombinant PP26, a periplasmic protein, reacted most strongly to both B. abortus–positive serum samples shown by Western blot analysis and to TAT-positive serum samples by an indirect ELISA among the protein antigens used in the current study. The highest mean absorbance value was observed in the ELISA using recombinant PP26 and TAT-positive serum samples, and the sensitivity of recombinant PP26 was comparable to that of B. abortus whole cell lysates. These results are consistent with a previous report, indicating that the 26-kDa periplasmic protein had a high agreement with the conventional test for bovine brucellosis. 21
The sensitivity and specificity of the recombinant Brucella abortus proteins and B. abortus whole cell lysates for seropositive or seronegative sera determined by tube agglutination test standard assay.
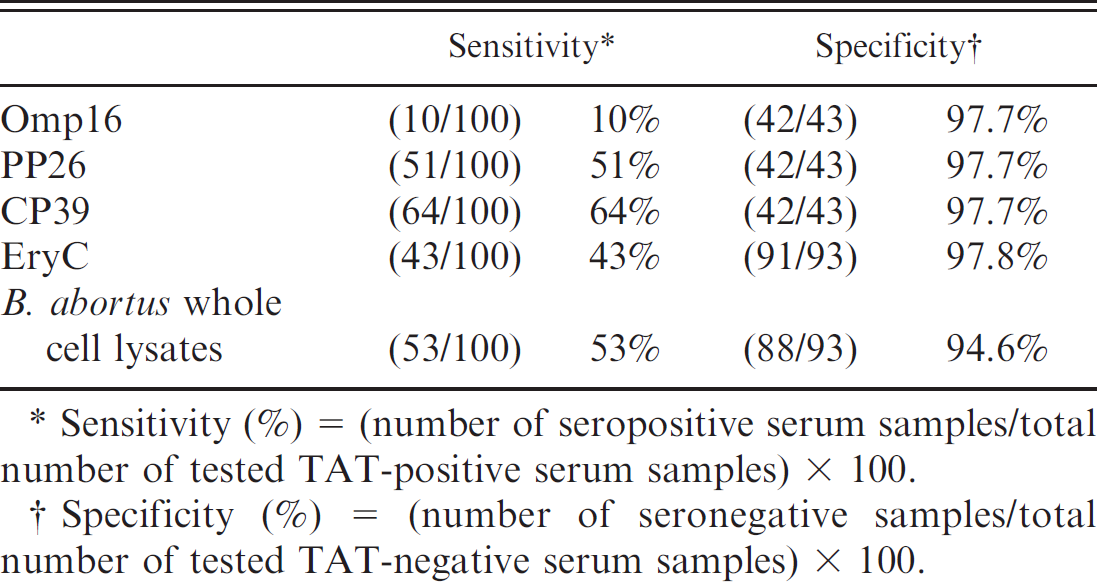
Sensitivity (%) = (number of seropositive serum samples/total number of tested TAT-positive serum samples) × 100.
Specificity (%) = (number of seronegative samples/total number of tested TAT-negative serum samples) × 100.
EryC,
The sensitivity of B. abortus antigens was more than 90% for bovine brucellosis in previous reports, 3,10,24 but in the current study, the sensitivity of B. abortus whole cell lysates for TAT-positive serum samples was approximately 50%. False-positive reactions may be present in TAT-negative samples that were identified using S-LPS as a diagnostic antigen, thus B. abortus–uninfected cattle might be diagnosed as TAT positive. Other serologic tests would be required to confirm this issue. In Korea, bovine brucellosis is prevalent. Therefore, B. abortus–infected cattle might also be diagnosed as TAT negative, which could increase the cutoff value due to high seroreactivity. The present study suggests that a novel protein of B. abortus, EryC, as well as PP26 may serve as diagnostic reagents for bovine brucellosis, as these proteins had a high sensitivity and specificity for bovine brucellosis. In addition, EryC may be used to discriminate B. abortus–infected cattle from B. abortus S19–vaccinated cattle.
Acknowledgements
This work was supported in part by a grant from the Ministry of Agriculture, Forestry and Fisheries-Special Grants Research Program (MAFF-SGRP) in Korea, by a grant from the Korean Health 21 R&D Project, the Ministry of Health and Welfare, Korea (A010381), and by a grant from the Brain Korea Project for the Medical Science in Yonsei University.
Footnotes
a.
Invitrogen, Carlsbad, CA.
b.
Difco, Detroit, MI.
c.
Sigma Aldrich, St. Louis, MO.
d.
National Center for Biotechnology Information of the National Library of Medicine, Bethesda, MD.
e.
Novagen, Darmstadt, Germany.
f.
Qiagen Inc., Valencia, CA.
g.
Clontech Laboratories Inc., Mountain View, CA.
h.
KPL Inc., Gaithersburg, MD.
i.
Amersham Pharmacia Biotech, Stockholm, Sweden.
j.
Costar Corp., Cambridge, MA.
k.
Molecular Devices®, BioTek Instruments Inc., Hyland Park, VA.
l.
GraphPad Software, San Diego, CA.