
Abstract
Proventricular dilatation disease (PDD) is a neurologic disease of psittacine birds suspected to be caused by a recently identified Avian bornavirus (ABV). In the current report, data supporting the causal association of ABV with PDD are presented. Immunohistochemistry (IHC) with rabbit polyclonal antiserum raised against ABV nucleocapsid protein was used to identify cell and organ distribution of viral antigen. The ABV antigen was most consistently detected in brain, spinal cord, adrenal gland, pancreas, and kidney. Histopathologic evaluation was correlated with ABV-specific polymerase chain reaction (PCR) and immunohistochemical tests in multiple tissues from 16 psittacine birds with and without PDD. Using histopathologic diagnosis as the gold standard, the sensitivity and specificity of IHC for ABV antigens were found to be 100% and 100%, respectively. Many more tissues were positive for ABV RNA by reverse transcription PCR than were positive for pathologic changes or viral antigens by IHC, indicating the presence of subclinical or asymptomatic infection at many sites.
Keywords
Introduction
Proventricular dilatation disease (PDD) was identified in the late 1970s and early 1980s as a pathologic syndrome of unknown etiology affecting psittacine birds. The disease was first reported in macaws imported from Bolivia into North America (Woerpel RW, Rosskopf WJ Jr., Hughes E: 1984, Proventricular dilatation and wasting syndrome: myenteric ganglioneuritis and encephalomyelitis of psittacines; an update. In: Proceedings of the International Conference on Avian Medicine of the Association of Avian Veterinarians, pp. 25–28. East Northport, NY). 21 Since then, the disease has been reported in Europe, Asia, and Australia and has been identified in over 50 psittacine species 13,26 and even some nonpsittacine species, 4,22 The disease is known by several synonyms including macaw wasting disease, myenteric ganglioneuritis, infiltrative splanchnic neuropathy, proventricular dilatation of psittaciformes, neuropathic gastric dilatation of psittaciformes, and psittacine encephalomyelitis. 13 The reported case mortality rate approaches 100%, 13 as clinical identification of early or subclinical infections has not been possible. Psittacine birds affected with PDD can exhibit gastrointestinal signs (anorexia, emaciation, weight loss, lethargy, regurgitation, diarrhea, and the presence of undigested seeds in the feces), neurologic signs (ataxia, tremors, seizures, and motor or proprioceptive deficits), or a combination of both. 3,13,20
Although a variety of clinical techniques, particularly plain and contrast radiography, and fluoroscopic evaluation of gastrointestinal motility, have been used for tentative antemortem diagnosis (Bond MW, Downs D, Wolf S: 1993, Screening for psittacine proventricular dilatation syndrome. In: Proceedings of the Annual Conference of the Association of Avian Veterinarians, pp. 92–97. Nashville, TN; Taylor M, Dobson H, Hunter DB, Atkinson R: 1997, New research in psittacine gastrointestinal motility in normal and disease states. In: Proceedings of the Annual Conference of the Association of Avian Veterinarians, pp. 131–132), 7,20,25 histopathologic examination of the affected tissues remains the gold standard for definitive diagnosis of the condition. The characteristic lymphoplasmacytic infiltrates were initially described in the myenteric ganglia of the crop, proventriculus, ventriculus, or duodenum; however, lesions can occur in many tissues of the central and peripheral nervous systems. 1 Confirmation of the disease in live birds has relied on the identification of diagnostic lesions in biopsy tissue. Full thickness biopsies of the proventriculus and ventriculus are considered invasive with increased morbidity and mortality when compared with biopsy of the ingluvies (crop). However, crop biopsy is less sensitive, since only 68–76% of birds with PDD confirmed histopathologically at necropsy had lesions in this organ (Bond MW, et al.: 1993, Screening for psittacine proventricular dilatation syndrome; Doolen M: 1994, Crop biopsy—a low risk diagnosis for neuropathic gastric dilatation. In: Proceedings of the Annual Conference of the Association of Avian Veterinarians, pp. 193–196). 12 A method for accurate, noninvasive antemortem diagnosis of the disease is essential to identify affected birds and prevent spread of the disease.
A variety of viral agents including paramyxo-viruses, togaviruses, picornaviruses, and coronaviruses have been proposed as the causative agent of PDD in psittacines 10,11,13,14,17,21 ; however, further studies have failed to provide evidence that these viral agents are actually associated with the disease. A novel Avian bornavirus (ABV) has recently been proposed as a candidate etiologic agent of the disease, 18 and there is strong supporting evidence from bird inoculation studies 9 and outbreak investigations. 19 Other independent research findings have also supported this association. 16,23
Confirmation that PDD is in fact caused by ABV would allow the use of molecular biologic techniques for early antemortem diagnosis and would help prevent the spread of the disease between susceptible birds. It would also pave the way for further research regarding the treatment and prevention of the disease. The purpose of the present study was to investigate a causal association of ABV with PDD by assessing the association between detection of ABV using reverse transcription polymerase chain reaction (ABV-specific RT-PCR) and immunohistochemistry (IHC) versus the diagnosis of PDD in psittacine birds based on histopathology (the current gold standard).
Materials and methods
Tissues and cloacal swab samples from 16 psittacine birds were used for the study (Table. 1). Twelve birds belonged to a research colony that had been created 3 years previously to study fecal virus shedding in psittacine birds with PDD. All birds had a history of exposure to other birds with PDD; however, the actual PDD status of each bird (based on histopathologic evidence) was not known at the time of the study. Three out of these 12 birds (birds 2, 7, and 12) were reported to have shown neurologic signs temporarily within the past year, and 2 of the birds (birds 1 and 3) showed neurologic signs at the time of euthanasia. The clinical signs noted on observation of these birds at rest included ataxia, difficulty perching, tremors, and head tilt. The birds were housed in the same room and managed similarly. All the birds were maintaining their body weight on a pelleted ration and showed no clinical signs of gastrointestinal disease over the course of the study. All birds were housed alone in their respective cages throughout their time in the research colony, with the exception of birds 4 and 5 and birds 6 and 7, which had been kept as pairs. Four additional birds were individual pet psittacine birds submitted for necropsy to the Department of Pathobiology, Ontario Veterinary College, University of Guelph (Guelph, Ontario, Canada). Birds 13 and 14 had no clinical or histopathologic evidence of PDD (and had other known causes of death). They were selected for inclusion in the study as positive and negative controls. Birds 15 and 16 had been clinically identified as having PDD, which was histopathologically confirmed at necropsy.
To assess the shedding frequency of the virus using ABV-specific RT-PCR primers directed against the matrix (M) and nucleocapsid protein (N) genes, 18 fresh fecal samples were collected individually from 5 birds in the research colony (birds 1–5) that had been previously identified as ABV RT-PCR positive based upon fecal sampling. Samples were collected 5 times daily (8:00 AM, 10:00 AM, 12:00 PM, 2:00 PM, and 4:00 PM) for 5 consecutive days. The samples were collected in an RNA stabilization solution, a kept at 5° for 24 hr, further mixed using a vortex, and stored at —7°C. The 5 samples from each individual bird, on each day, were pooled together and were evaluated with the ABV RT-PCR assay. In the week prior to depopulation of the research colony, cloacal and choanal swabs were collected from all 12 birds (birds 1–12) and processed, stored, and assayed as described above.
Specimen collection
The 12 birds from the research colony were euthanized to provide tissues for ABV identification. The birds were anesthetized with isoflurane b and were subsequently euthanized with intravenous potassium chloride. c All procedures involving animals were approved by the University of Guelph Animal Care Committee in accordance with the requirements of the Animals for Research Act of Ontario, revised 1990, and the recommendations of the Canadian Council for Animal Care.
A complete necropsy was performed on the 12 birds, and tissue samples were collected as described below. Two samples each of cerebellum, cerebrum, brain stem, spinal cord, sciatic nerve, brachial nerve, vagus, crop, esophagus, proventriculus, ventriculus, duodenum, small intestine, colon, cloaca, spleen, kidney, heart, liver, lungs, pancreas, gonad, skeletal muscle, and skin, and 1 sample each of adrenal gland, thyroid, parathyroid, and eye were immediately frozen and stored at —70°C for evaluation by ABVRT-PCR. An additional sample from each of the above tissues was placed in 10% neutral buffered formalin for histopathologic examination and further immunohistochemical staining. Available frozen tissues and their formalin-fixed, paraffin-embedded counterparts from the other 4 birds (birds 13–16) were used for ABV RT-PCR, and for immunohistochemical and histopathologic examination, respectively (Tables 2–4). Many, but not all, relevant tissues were available for both forms of evaluation.
Study population of psittacine birds with and without proventricular dilatation disease.*
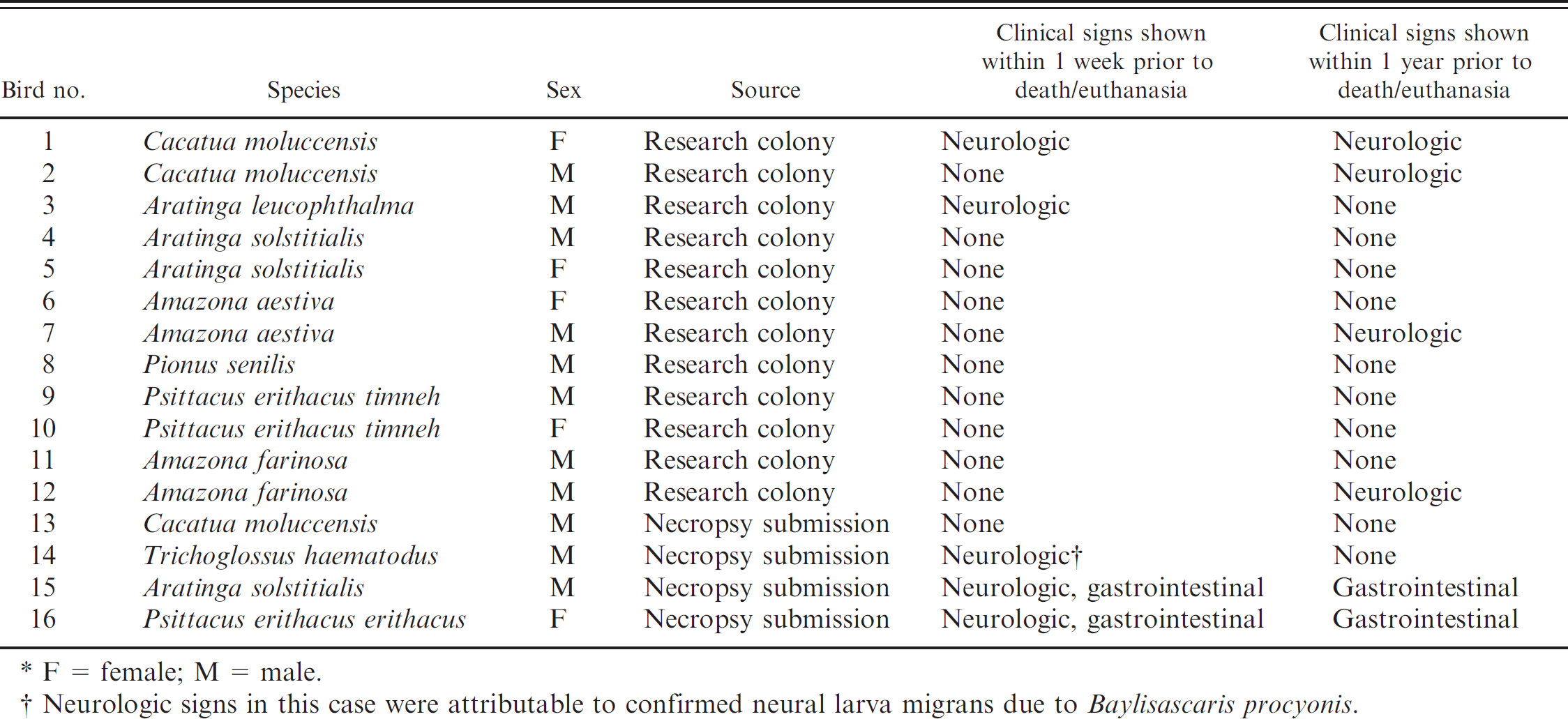
F = female; M = male.
Neurologic signs in this case were attributable to confirmed neural larva migrans due to Baylisascaris procyonis.
Histopathologic analysis
Fixed necropsy tissues were routinely processed, paraffin-embedded, sectioned at 4-μm, and stained with hematoxylin and eosin (HE). The tissues were evaluated blindly (by case) for the presence of lymphoplasmacytic infiltrates, particularly in the nervous tissue, as well as for the presence or absence of other pathologic lesions. Evaluations were conducted initially by one author (RR), and then confirmed by another author, a pathologist (DS). Presence of characteristic lymphoplasmacytic infiltrates in the myenteric ganglia of the crop, proventriculus, or ventriculus, and in the ganglia adjacent to the adrenal gland and the heart was used as the primary criterion for diagnosing a bird as PDD positive. Other lesions suggestive of PDD, including perivascular cuffing of blood vessels in the brain and spinal cord, lymphoplasmacytic adrenalitis, and peripheral neuritis were noted and used as secondary criteria to aid the histopathologic diagnosis.
Detailed description of the cloning, recombinant expression, purification, and generation of polyclonal antisera against the ABV nucleocapsid (ABVN) protein has been previously described. 9 Briefly, the open reading frame (ORF) of the ABV2 nucleocapsid protein (ABVN) gene was recovered through RT-PCR of total RNA derived from crop tissue containing the fully sequenced ABV2_bil isolate (GenBank accession no. EU781967). The resulting ABVN RT-PCR product was subcloned into a pMAL vector, generating an expression construct to produce a recombinant version of ABVNp flanked by a maltose-binding protein tag (MBP) and Tobacco etch virus (TEV) protease cleavage site (tev) at its N-terminus and 6xHIS (His6) amino acid tag at its C-terminus. This expression construct was transformed into pRIL+BL21(DE3) Escherichia coli. Expression of recombinant protein was induced with addition of 250 μM of isopropyl beta-D-thiogalactoside (IPTG) for 4 hr at 37°C, and resulting MBV-tev-ABVN-His6 was purified from cell lysates via Ni2+-nitrilotriacetic acid and amylose column chromatography. The N-terminal MBP tag was removed via cleavage with TEV protease, and the cleaved ABVN-His6 protein was purified by ion exchange chromatography, followed by size fractionation on a 24-ml gel filtration column, d and concentrated via centrifugation with a 15. kD centrifugal filter unit. e Two and one-half milligrams of this purified ABVN-His6 protein was utilized for polyclonal antisera generation in rabbits. f This antisera has been shown to recognize the ABVN protein by Western blot.
Distribution of histopathologic lesions attributable to proventricular dilatation disease in the study population.*
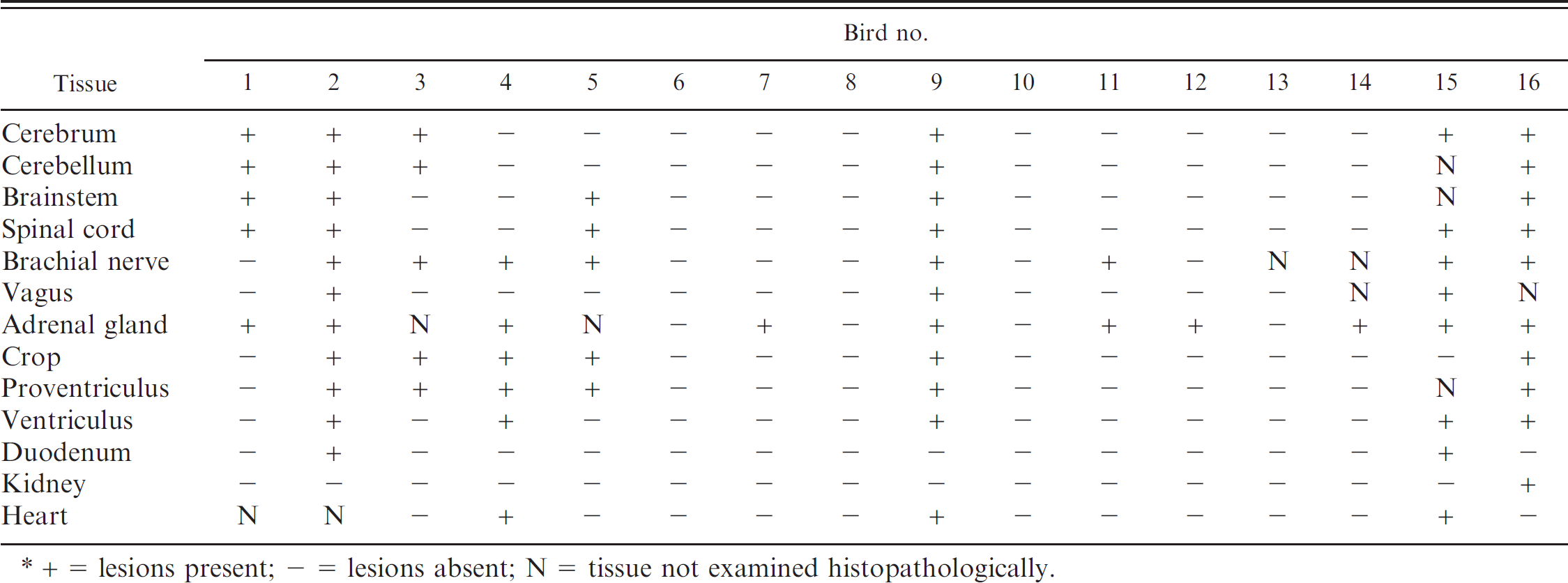
+ = lesions present; - = lesions absent; N = tissue not examined histopathologically.
Tissue distribution of Avian bornavirus nucleocapsid protein antigen by immunohistochemistry.*
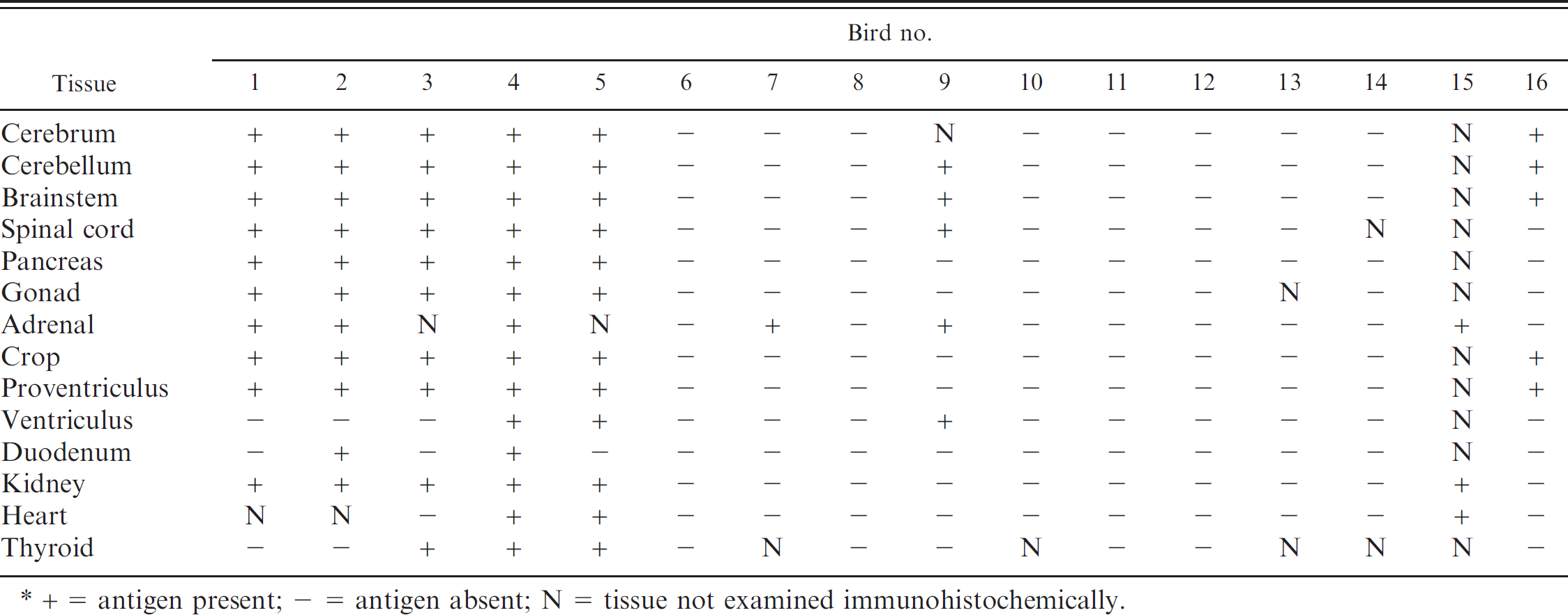
+ = antigen present; - = antigen absent; N = tissue not examined immunohistochemically.
Results of reverse transcription polymerase chain reaction (RT-PCR) testing for Avian bornavirus matrix (ABVM) and nucleocapsid protein (ABVN) genes on cloacal swabs, choanal swabs, and tissues from 16 psittacine birds with and without proventricular dilatation disease.*
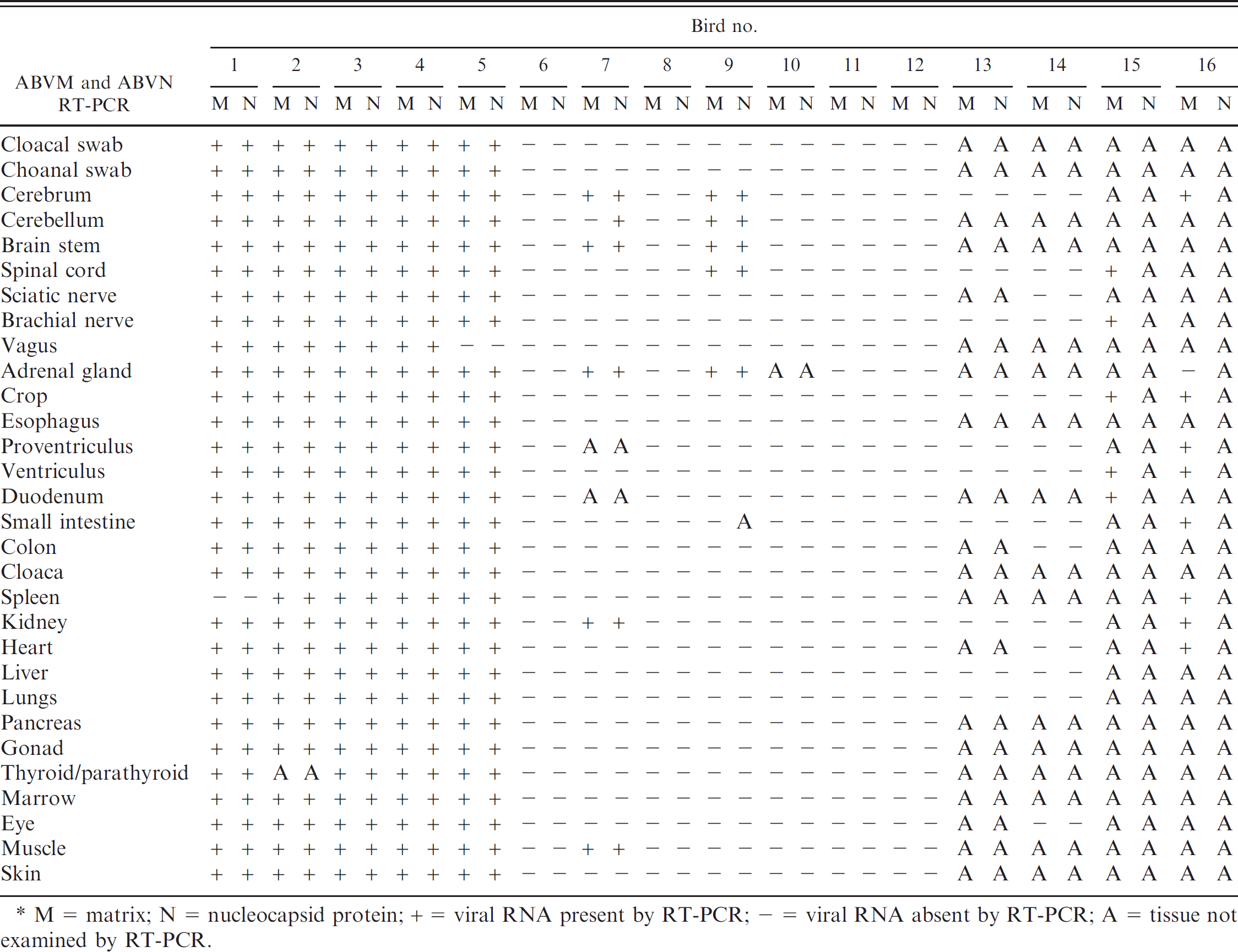
M = matrix; N = nucleocapsid protein; + = viral RNA present by RT-PCR; - = viral RNA absent by RT-PCR; A = tissue not examined by RT-PCR.
Immunohistochemical analysis
Immunohistochemistry was performed on 4-μm tissue sections mounted on charged slides and included all paraffin-embedded tissue blocks examined by light microscopy using HE stains. All processing was done on an automated staining instrument, g using rabbit polyclonal antiserum raised against ABVN protein. All incubations were completed at room temperature. Following manual deparaffinization and rehydration, tissue sections were treated with 3% hydrogen peroxide to quench endogenous peroxidase activity. Antigen retrieval was accomplished by incubation with proteinase K h for 6 min. Sections were then incubated with rabbit anti-ABVN antiserum (1:2,000) for 30 min, followed by 30-min incubation with a goat anti-mouse/rabbit polymer i visualization system. Nova Red j was used as the chromogen. For negative reagent controls, duplicate sections of each control and test tissue were subjected to the same immunohistochemical procedure with substitution of pre-immune rabbit serum at similar protein concentration for the anti-ABVN antisera. Slides were blinded by case and interpreted by one author (RR).
Since mammalian Borna disease virus (BDV) is known to replicate intranuclearly, 5 the criterion for immunohistochemical case-positive classification included strong and diffuse intranuclear staining in the parenchymal cells of 1 or more tissues. Birds were classified as IHC-negative when intranuclear staining with the anti-ABVN antisera was absent.
Nucleic acid isolation and RT-PCR
Choanal and cloacal swabs, and thawed frozen tissue samples collected at necropsy, were evaluated for the presence of ABV by RT-PCR detection of ABV matrix (ABVM) and ABVN genes. Total nucleic acids were extracted using a total RNA isolation kit k from 50-μl aliquots of 10% tissue suspensions, or from 50-μl aliquots of inoculated virus transport media 1 when cloacal and choanal swabs were used. Primers for ABV RT-PCR were those previously described. 18 The RT-PCR was carried out in 25-μl reactions using a one-step RT-PCR kit. m A case was classified as ABV-PCR positive if 1 or more tissues tested positive with ABVM or ABVN RT-PCR. A case was classified as negative if all tissues tested with ABVM and ABVN RT-PCR were found to be negative.
Sequencing and sequence analysis
Partial nucleotide sequences of ABVM and ABVN genes were determined at the Guelph Molecular Supercentre, University of Guelph. Sequences were assembled with the SeqMan module of Lasergene, software. n The sequences reported in the present study have been deposited in GenBank under accession numbers GQ496343, GQ496344, GQ496345, GQ496346, GQ496347, GQ496348, GQ496349, GQ496350, GQ496351, GQ496352, GQ496353, GQ496354, GQ496355, GQ496356, GQ496357, GQ496358, GQ496359, and GQ496360.
Sequence comparisons were carried out using a 345-base pair (bp) fragment of the N gene from nt 659 to nt 1003 and a 270-bp fragment of the M gene from nt 1972 to nt 2241, relative to complete genomic sequence of ABV isolate bil, GenBank accession no. EU781967. Multiple alignments were carried out with the MegAlign module of Lasergene software using the default settings of its ClustalW algorithm. Phylogenetic trees were also generated with the MegAlign module of Lasergene software using its default settings.
Statistical analyses
Statistical analyses were performed by 2 authors (RR and DP) using Stata statistical computer software.° Test agreement was assessed for individual tissue type and for the diagnosis of each bird based on the results of all tissues tested. Agreement between tests was evaluated using Cohen's kappa statistic after assessing the presence of any test bias using the Exact McNemar's test to determine any significant differences in the proportions positive to the tests being compared. 8 Histopathologic diagnosis of each bird was used as the gold standard, and the epidemiologic specificity and sensitivity of the different tests were derived using the Stata software.
Results
Histopathologic analysis
Among the 16 birds in the study population, 8 were histopathologically diagnosed with PDD, and 8 birds did not have lesions attributable to PDD (Table. 5). The lesions observed included lympho-plasmacytic myenteric ganglioneuritis in the crop (6/8), proventriculus (6/7), ventriculus (5/8), and duodenum (2/8); lymphoplasmacytic peripheral neuritis in the brachial (7/8) and vagus (3/7) nerves; lymphoplasmacytic adrenalitis with or without lymphoplasmacytic infiltration in the adjacent nerves (6/6); lymphoplasmacytic infiltration in the ganglia adjacent to the heart (3/6) and kidney (1/8); and lymphoplasmacytic perivascular cuffing in various parts of the central nervous system including cerebrum (6/8), cerebellum (5/7), brainstem (5/7), and spinal cord (6/8). Peripheral neuritis in the brachial nerve (1/6) and lymphoplasmacytic adrenalitis (4/8) were also observed in birds that were histopathologically diagnosed as negative for PDD. The tissue distribution of histopathologic lesions in positive and negative birds is shown in Table 2. Seven of the 8 positive birds had pathognomonic lesions in the myenteric ganglia of the gastrointestinal system. The other positive bird (bird 1) had characteristic lymphoplasmacytic infiltrates in the ganglia adjacent to the adrenal gland as well as suggestive lesions in the brain, spinal cord, and adrenal gland.
Histopathology, immunohistochemistry (IHC), and reverse transcription polymerase chain reaction (RT-PCR) test results by bird.*
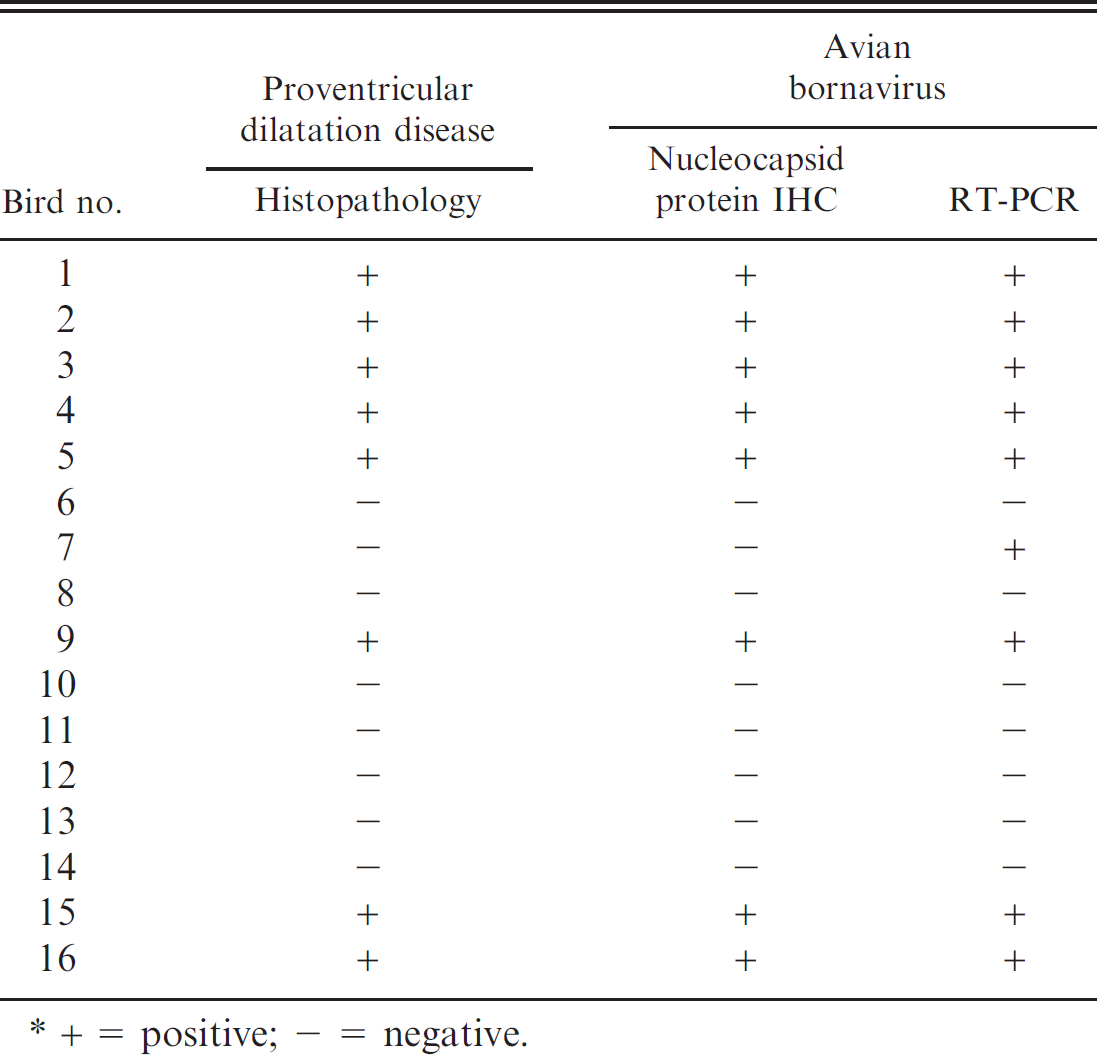
+ = positive; - = negative.
An additional finding was the presence of lymphoplasmacytic infiltrates in the renal interstitium of the majority (7/8) of histopathologically positive birds (birds 1–4, 9, 15, and 16), as well as in 2 out of 8 histopathologically negative birds (birds 7 and 12).
Immunohistochemical analysis
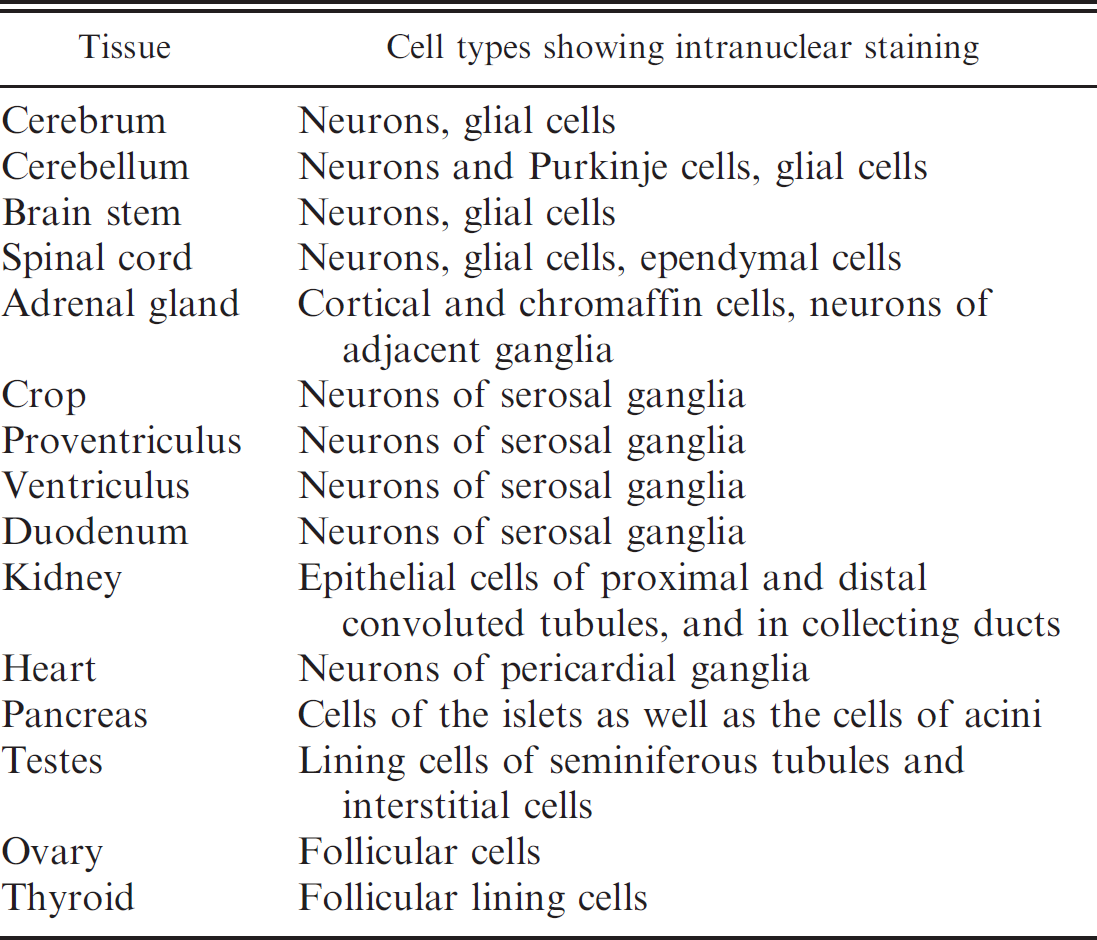
Distinct intranuclear staining for ABVN antigen was detected in various tissues from 8 birds, all of whom had histopathologically confirmed PDD. In contrast, no specific intranuclear staining for ABVN was observed in tissues from the remaining 8 PDD-negative birds (Table. 5). Table 3 lists the tissues examined and the tissue distribution of ABVN antigen in all the birds; Table 6 documents the observed staining characteristics and cell distribution of the antigen in those tissues.
In those birds positive for ABV by IHC, staining for ABVN antigen in brain and spinal cord was present in more than 50% of the neurons per high power field (40x). ABVN antigen was also demonstrated in nuclei of neurons within ganglia throughout the body, including those in the crop, proventriculus, ventriculus, duodenum, heart, and adrenal gland (Fig. 1).
Moderately intense, diffuse, intracytoplasmic staining accompanied intranuclear staining in some neurons and renal tubular epithelial cells and was interpreted as specific staining. Stromal and intracytoplasmic staining of varying, but generally lesser, intensity and unaccompanied by nuclear staining was present in multiple tissues and cell types in both IHC-positive and IHC-negative birds and was interpreted as spurious immunoreactivity not associated with ABVN antigen. This staining pattern was most prominent in the proventricular glands; however, intravascular leukocytes, vascular endothelial cells, cell membranes of adipocytes and myocytes, and collagen in multiple tissues demonstrated this non-ABVN-associated pattern. No nonspecific or background staining was evident in negative reagent control slides from any of the 16 birds tested.
Polymerase chain reaction
Out of the 16 birds tested in a blinded fashion, 9 birds (birds 1–5, 7, 9, 15, and 16) had positive signal for ABVM or ABVN RT-PCR on 1 or more tissue samples and were classified as ABV RT-PCR positive (Tables 4, 5); all but 1 of these birds (bird 7) were classified histopathologically as having PDD. Seven birds were negative for ABV RT-PCR on all tissue samples tested; none of these birds were positive for PDD histopathologically (Tables 4, 5).
In general, in infected birds, more tissues were positive by RT-PCR than by IHC; this is not an entirely unexpected result, given the known higher sensitivity of PCR-based amplification methods: bird 1 (27/28), bird 2 (27/27), bird 3 (28/28), bird 4 (28/28), bird 5 (27/28), bird 15 (5/5), and bird 16 (8/9). Of the remaining ABV RT-PCR-positive birds, ABV was detected in fewer, primarily neural, tissues (Table 4; birds 7 and 9).
Cloacal and choanal swabs were ABVM and ABVN RT-PCR positive in 5 out of 12 birds tested (birds 1-5). These 5 birds were histopathologically confirmed PDD-positive cases that were positive by ABV RT-PCR in multiple tissues (Table 4). The swabs were negative in 2 birds that were RT-PCR positive on a few tissues (birds 7 and 9) and in the 5 birds whose tissues were negative by ABV RT-PCR.
Cellular distribution of intranuclear immunohistochemical staining for Avian bornavirus nucleocapsid protein antigen in immunohistochemistry-positive birds.
Sequencing
The ABV RT-PCR products recovered from the birds were sequenced and compared with the publicly available ABV sequences present in GenBank (Fig. 2). Based on these results, it appears that 2 different isolates of ABV were detected. ABVM and ABVN sequences recovered from tissues of birds 1–3, 9, and 16 were virtually identical to each other (95.1–100% average pairwise nucleotide identity) and to the ABV4 sequences present in GenBank (93.9–100% average pairwise nucleotide identity). In contrast, the ABVM and ABVN sequences recovered from the tissues of birds 4, 5, 7, and 15 were nearly identical to each other (100% average pairwise nucleotide identity) and to the ABV2 sequences present in GenBank (95.1–100% average pairwise nucleotide identity).
Statistical associations between tests
Test comparisons were initially made comparing birds classed as positive and negative based on a parallel interpretation of results, where multiple tissues were examined from each bird with each test (i.e., only 1 tissue had to test positive with a particular test for a bird to be classified as positive for that test). Based on Exact McNemar's tests, there were no significant differences in the proportions positive to each of the tests (histopathologic diagnosis, IHC, and RT-PCR) compared; therefore Cohen's kappa statistic could be used to assess the degree of agreement beyond what would have been expected by chance. Cohen's kappa statistic of agreement between histopathologic diagnosis and IHC was 100% (observed agreement = 100%; expected agreement = 50%; P-value < 0.0001), and was 100% (observed agreement = 100%; expected agreement = 50%; P-value = 0.0001) between ABVM and ABVN RT-PCR. Cohen's kappa statistic of agreement between histopathologic diagnosis and ABV RT-PCR (ABVM and ABVN RT-PCR considered together) was 87.5% (observed agreement = 93.75%; expected agreement = 50%; P-value = 0.0002). Cohen's kappa statistic of agreement between ABV RT-PCR and IHC was 87.5% (observed agreement = 93.75%; expected agreement = 50%; P-value = 0.0002).
When the results were compared according to the type of tissue tested, there was a statistically significant high level of agreement among histopathologic lesions, IHC, and ABV RT-PCR for 2 groups of tissues: those of the gastrointestinal system (crop, esophagus, proventriculus, ventriculus, duodenum, small intestine, colon, and cloaca considered together) and those of the nervous system (cerebrum, cerebellum, brainstem, spinal cord, brachial nerve, sciatic nerve, and vagus considered together). Narrowing test performance analysis to ABVN IHC and ABV RT-PCR yielded a high level of agreement for all the tissues (Table 7).
Histopathologic diagnosis as the gold standard
If histopathologic diagnosis of PDD is used as the gold standard, the sensitivity and specificity of IHC were 100% (95% confidence interval [CI]: 63.1%, 100.0%) and 100% (95% CI: 63.1%, 100.0%), respectively. Using the same criteria, the sensitivity and specificity of RT-PCR (ABVM and ABVN RT-PCR considered together) were 100% (95% CI: 63.1%, 100.0%) and 87.5% (95% CI: 47.3%, 99.7%), respectively. With the same criteria, the sensitivity and specificity of ABVM RT-PCR were 100% (95% CI: 63.1%, 100.0%) and 87.5% (95% CI: 47.3%, 99.7%), respectively, and those of ABVN RT-PCR were 100% (95% CI: 54.1%, 100.0%) and 87.5% (95% CI: 47.3%, 99.7%), respectively.
Assessment of the shedding frequency of the virus using RT-PCR
Finally, prior to euthanasia, the authors also investigated whether live birds could shed ABV in their stool, and if so, whether such shedding was continuous or intermittent. As shown in Table 8, ABV shedding was frequently intermittent, with only 2 of 5 serially tested birds excreting ABV virus continuously without interruption. (Bird 1, which tested negative on all 5 days, had tested positive on a previous cloacal swab.) The median number of positive ABV RT-PCR tests on fecal samples over 5 sampling days was 3, with a range of 0-5 positive tests.
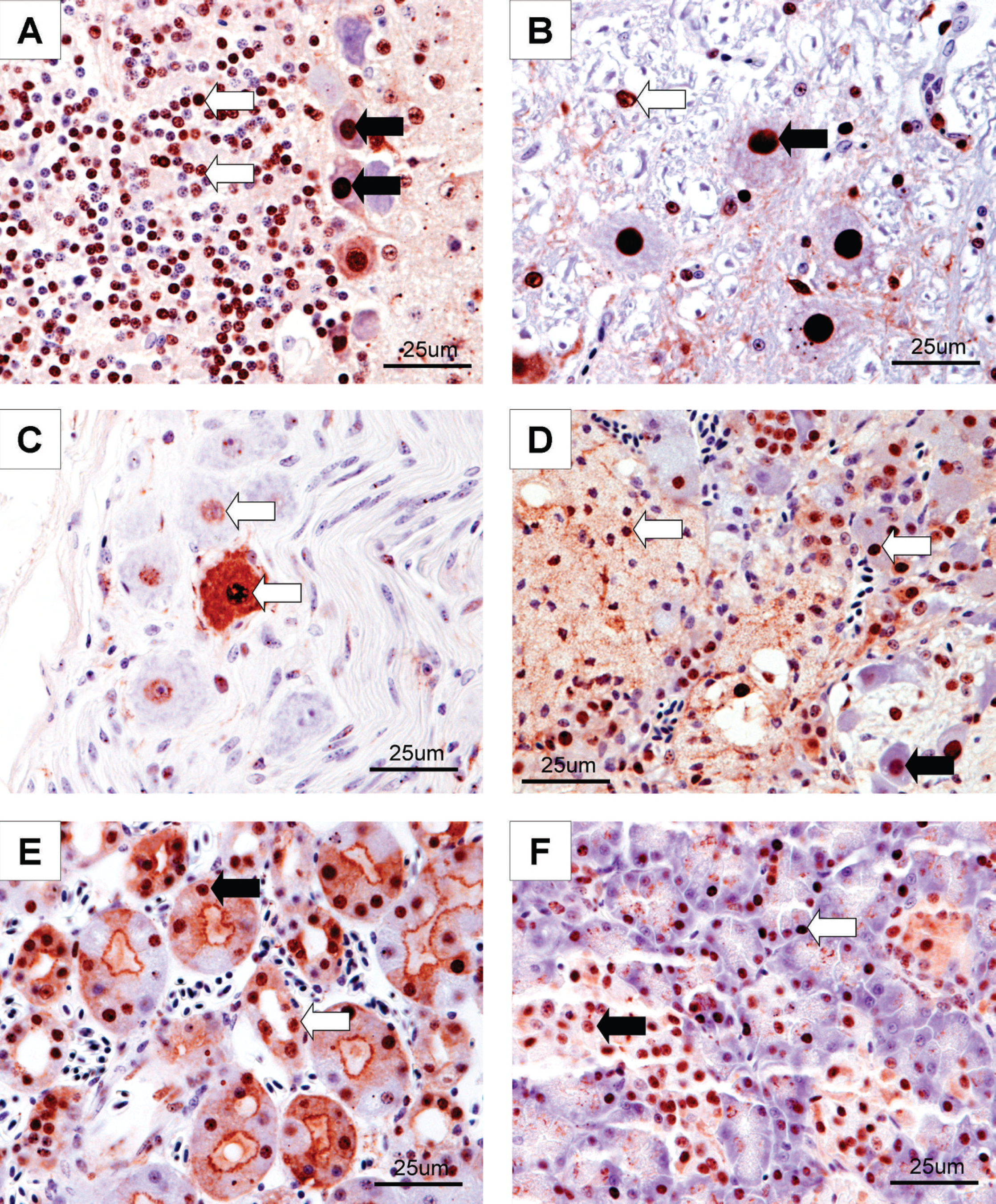
Immunohistochemical staining for Avian bornavirus (ABV) antigen (40X) in psittacine birds with proventricular dilatation disease. Intranuclear brown staining shows the location of the ABV antigen in these tissues.

Phylogenetic comparison of Avian bornavirus (ABV) matrix (
Discussion
The striking agreement between the histopathologic diagnosis of PDD and the detection of ABV using specific RT-PCR and immunohistochemical tests on tissues strongly supports the association of ABV with PDD and is consistent with (though not conclusive for) an etiologic role for ABV in that disease. The presence of characteristic histopathologic lesions is the established gold standard for diagnosis of PDD, and the authors of the present study have adhered to that criterion. The range of histopathologic lesions seen in affected birds was similar to and consistent with those reported previously in the veterinary literature (Hughes PE: 1984, The pathology of myenteric ganglioneuritis, psittacine encephalomyelitis, proventricular dilatation of psittacines, and macaw wasting syndrome. In: Proceedings of the 33rd Western Poultry Disease Conference, pp. 85–87. Davis, CA). 1,13
Statistical association between histopathologic diagnosis of proventricular dilatation disease (PDD) and identification of Avian bornavirus by immunohistochemistry (IHC) and reverse transcription polymerase chain reaction (RT-PCR) for various tissue types in psittacine birds with and without PDD.*
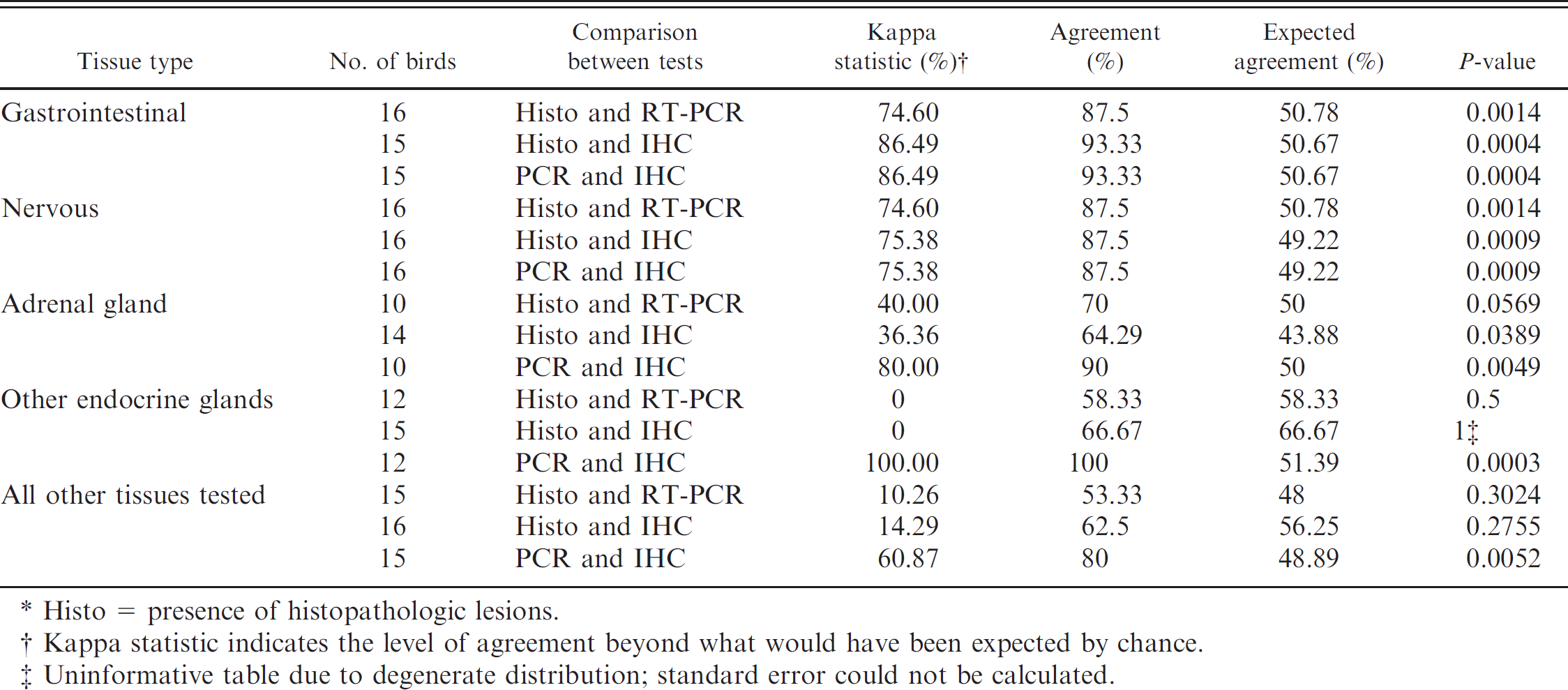
Histo = presence of histopathologic lesions.
Kappa statistic indicates the level of agreement beyond what would have been expected by chance.
Uninformative table due to degenerate distribution; standard error could not be calculated.
Using RT-PCR, ABV RNA was detected in 100% (8/8) of the birds with histopathologically confirmed PDD, and in 1 additional bird that did not have histopathologic lesions. This is in comparison to previous publications where 71% of birds (5/7) with histopathologic evidence of PDD were ABV RT-PCR positive on samples of crop, proventriculus, and ventriculus, 18 and 46.15% (6/13) birds with clinical signs consistent with PDD tested positive using ABV RT-PCR on crop biopsies, blood samples, and brain tissues. 23 The ABV RT-PCR assay used in the present study and the 2 previously published studies 18,23 were based on the same published sequences. 18 The higher success rate of ABV detection in the present study can be attributed to the sampling of multiple tissues from each bird (thereby increasing the sensitivity of testing), and to strict case determination criteria. The first study 18 used histopathologically confirmed cases of PDD, but ABV RT-PCR was performed on fewer tissues than were examined in the current study. The second study 23 again used fewer tissues per bird, and also included birds with clinical signs consistent with PDD but without histopathologic confirmation of the disease.
Seven out of the 9 birds that were ABV RT-PCR positive for ABV tested uniformly positive for all or most of the tissues evaluated. In contrast, in 2 birds (birds 7 and 9), only a small number of tissues were positive; namely, brain, spinal cord, and adrenal gland (bird 9) and brain, spinal cord, adrenal gland, kidney, and skeletal muscle (bird 7). It is not known why these birds showed a different extent of viral infection; they were not the only representatives of their species and, therefore, other unknown underlying determinants of disease progression may have been involved. The tissues that were positive in all 9 RT-PCR-positive birds, and thus the obvious targets for diagnostic necropsy testing, were cerebrum, cerebellum, and brainstem, spinal cord, and adrenal gland. These tissues were not specifically examined in the 2 previous reports outlined above, 18,23 again highlighting the fact that tissue choice may be important in detection sensitivity. Central nervous system tissue is frequently not selected for histopathologic assessment for PDD in diagnostic cases, where sections of the crop, proventriculus, and ventriculus are considered most essential. 13
Assessment of the frequency of shedding of Avian bornavirus (ABV) in urofeces by 5 psittacine birds using ABV reverse transcription polymerase chain reaction.*
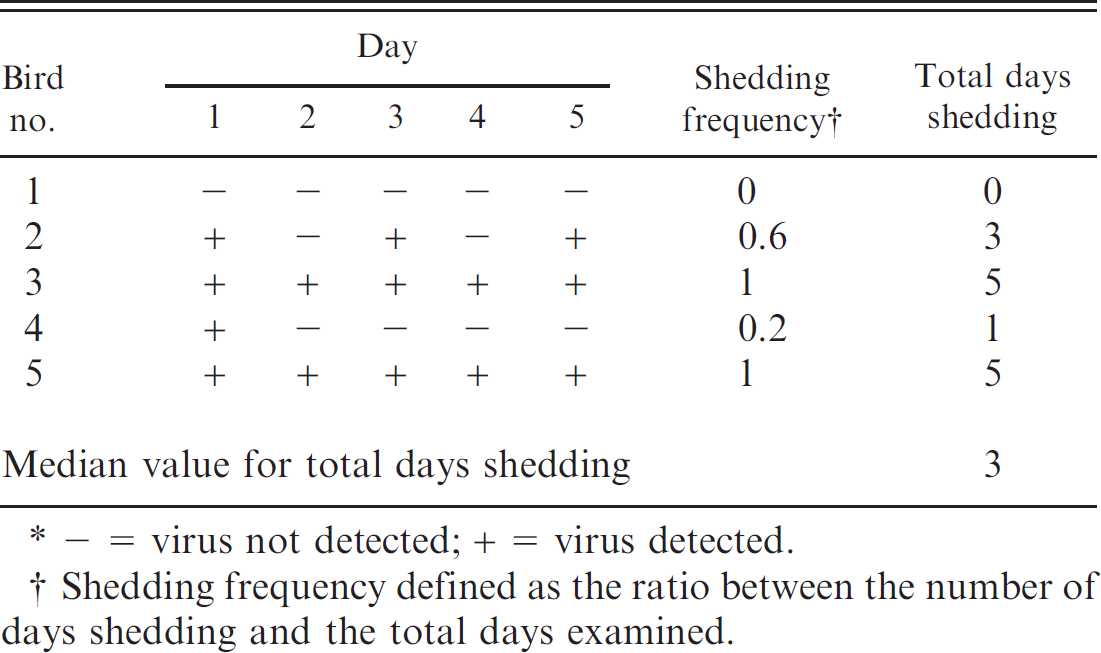
− = virus not detected; + = virus detected.
Shedding frequency defined as the ratio between the number of days shedding and the total days examined.
The use of RT-PCR to identify ABV RNA in a variety of secretions and excretions appears to have potential as a screening test for ABV-infected birds in psittacine populations. However, caution must be taken, since shedding was intermittent in the urofeces of birds that tested positive at necropsy. Hence, sampling each bird on multiple occasions would appear to be necessary to decrease the number of false-negative results. Further research on shedding frequency will be necessary to develop guidelines for diagnostic sample collection, particularly in acute clinical cases, and to provide sufficient data to determine sensitivity and specificity of a testing program. All 5 birds with numerous RT-PCR-positive tissues were also RT-PCR positive on both choanal and cloacal swabs taken shortly before euthanasia. The 2 birds with less extensive ABV tissue distribution were negative on both samples. Whether this reflects a true difference in viral extent and shedding, or simply the effect of single sampling, is not known.
Genotypes 2 and 4 of ABV were detected among the RT-PCR-positive birds. Viral sequences derived from birds 1 and 2 were nearly identical, which might be explained by the fact that these birds came from the same aviary. The viral sequences derived from the tissues of birds 4 and 5 were identical; these birds came from the same second source and had been kept together in the same cage. Bird 7 (from a third source) was housed in a cage adjacent to birds 4 and 5, and exhibited nearly identical viral sequences to these birds. But in contrast, bird 6, housed in the same cage as bird 7, was negative for ABV and PDD. As the birds were donated to the colony based on exposure to PDD, their actual status on admission was not known. Although in the clinical experience of the authors, close contact or fecal spread appears necessary for disease transmission. Careful sanitation was practiced, but birds were not held in isolation units; hence the possibility of viral transmission occurring in the research colony exists.
Successful development and performance of an immunohistochemical procedure to identify ABVN antigen in psittacine tissues using a rabbit polyclonal antibody raised against the ABV nucleocapsid protein was demonstrated in the current study. There was a statistically high level of agreement (kappa = 100%) between the diagnosis of cases by IHC and by histopathology.
Intranuclear staining for ABVN antigen was demonstrated in birds identified as ABV IHC positive, particularly in neurons and glial cells of various parts of the brain (20/20 samples) and spinal cord (6/7 samples), in the myenteric ganglia of the gastrointestinal system and in ganglia adjacent to the adrenal gland and heart. Similar IHC findings were reported in a recently published study describing experimental infection of cockatiels with ABV. 9 These tissues may represent sites of multiplication of the virus since BDV is known to replicate intranuclearly. 5 Intranuclear viral antigen was also identified in other tissues examined including pancreas, adrenal gland, gonads, thyroid gland, and in the cells of the proximal and distal convoluted tubules of the kidney and in the collecting ducts. These findings, and RT-PCR positivity for viral RNA at these and other sites, suggest that ABV has a broad tissue tropism. Only intranuclear staining was considered as truly indicative of viral infection in the present study. Cytoplasmic staining of myocytes, stromal elements, and epithelial cells of the gastrointestinal tract and lamina propria of the intestine was seen in both positive and negative cases (based on histopathology and RT-PCR) and hence is likely nonspecific staining caused by the primary antibody. Based on the current study, tissues of the central nervous system (including cerebrum, cerebellum, brain stem, and spinal cord), adrenal gland, kidney, and pancreas are preferred for immunohistochemical evaluation due to distinct intranuclear antigen localization of ABV antigen and minimal nonspecific staining seen in these tissues. Tissues of the gastrointestinal tract, including biopsy samples of crop, may also be useful for diagnosis; however, until techniques are developed to reduce nonspecific staining, the interpretation of mucosal and stromal staining must be made with caution.
This broad distribution of nuclear staining for viral antigen is in contrast with what is known about BDV. Borna disease virus has been reported to target neurons and glial cells in the central nervous system, particularly the limbic system, in most species of mammals that act as end-hosts, as well as in some laboratory animals, and virus-specific intranuclear inclusion bodies called Joest-Degen bodies are seen in the neurons of BDV-affected animals. 6,24 Inclusion bodies were not noted in any of the avian tissues examined in the present study. In bicolored shrews (Crocidura leucodon), which were proposed as reservoir hosts of BDV, the cardiomyocytes were also shown by RT-PCR to contain viral antigen. However, on IHC, the labeling was both intranuclear and intracytoplasmic, and the characteristic intranuclear inclusion bodies could not be demonstrated. 15
A previous study 23 in parrots using antiserum directed against the P protein of BDV had reported staining in 1 out of 2 birds tested. However, that study 23 described immunohistochemical staining in the nucleus and cytoplasm of cells of the brain, cardiomyocytes, hepatocytes, and in the epithelial cells and lamina propria of the intestine, and there was no discussion regarding staining of negative reagent or tissue controls to determine the degree of nonspecific cytoplasmic staining that may have occurred. Although the use of heterologous antisera and the presence of cytoplasmic staining limit interpretation of those results, the previous study's conclusion that ABV may display broader tissue tropism than BDV is consistent with the present findings.
The mechanism(s) of natural spread of ABV from one bird to another remains to be proven, but results from the current study show that virus is present in urofeces and in cloacal and choanal swabs of ABV-positive birds. A fecal-oral spread fits well with the clinical scenarios seen by the authors when dealing with disease outbreaks in the pet bird community. Intermittent shedding of virus, as was shown by the limited shedding trial in the present study, as well as low environmental stability of the mammalian BDV and possibly of ABV, may help explain the slow rate of transmission seen in situations where close attention is paid to biosecurity and fecal containment. The 12 birds in the study group in the present study lived in the same room for up to 3 years, yet 5 had no evidence of viral presence based on IHC and RT-PCR evaluation of multiple necropsy tissues. One of these 5, a blue-fronted Amazon (bird 6), in fact shared a cage with bird 7, which was positive for ABV on several tissues by PCR. Differing species susceptibility to infection and disease may also be important.
It was interesting to observe that the choanal swabs also tested positive in the 5 positive birds with a widespread distribution of virus. In mammals, it has been suggested that the primary route of infection of BDV could be through the olfactory neuroepithelium, which also acts as the primary virus replication site. 6 The positive choanal swabs could reflect the birds becoming re-infected or recontaminated from their own environment, or it might be inferred that the oral or nasal cavity could be a source of viral shedding. BDV has been shown to travel exclusively through neuronal conduit by centrifugal and centripetal migration along peripheral nerves to and from the central nervous system, 2 which would facilitate infection of a variety of organ systems and make possible a number of routes of viral transmission. It is quite possible that a similar mechanism of viral transport and shedding through multiple routes occurs with ABV; however, further research is needed to draw any such conclusions.
Using histopathologic diagnosis of PDD as the gold standard, the sensitivity and specificity of ABV-specific IHC has been established by the current study to be 100% and 100%, respectively, and that of ABV-specific RT-PCR to be 100% and 87.5%, respectively. The comparatively lower specificity of RT-PCR is based on a positive result for 1 bird (bird 7) that was categorized as negative for the disease on histopathology and IHC. Looking in more detail at the results for bird 7, the ABV-specific RT-PCR was positive by both M and N probes on cerebrum, brain stem, adrenal, kidney, and skeletal muscle (5/29 tissues examined), and by N probe alone on cerebellum. Bird 7 had also been reported as showing neurologic clinical signs in the past. Hence, it was posited that this represents a subclinical or latent form of infection, rather than a false-positive result. The single cloacal swab from this bird was negative for virus by RT-PCR, suggesting that the bird was not shedding virus at the time of sampling. Bird 6, housed in the same cage with bird 7, was negative for PDD (histopathologically) and ABV (by IHC and by RT-PCR) suggesting that there had not been significant shedding of virus into the environment. However, based on the shedding frequency trial in the present study, 1 sample is insufficient to make a confident assessment on whether shedding of virus is occurring.
Although histopathology has been used as the gold standard for diagnosis of PDD, it is well recognized that false-negative results can occur based on the uneven distribution of lesions, particularly in clinical biopsy material (Bond MW, et al.: 1993, Screening for psittacine proventricular dilatation syndrome). 13 Using the hypotheses that infection by ABV causes PDD, and that RT-PCR is a more sensitive method of identifying infection than histopathology, it is suggested that ABV-specific RT-PCR might be a better technique to use for the screening of psittacine birds at risk for PDD, especially for detection of early infections. Data from the current study suggest that the use of histologic criteria alone would lead to a significant underestimation of the prevalence of ABV infection in a psittacine population, and that performing IHC and/or RT-PCR on crop biopsy specimens would help to increase the sensitivity of diagnosis of PDD, allowing earlier identification of infected birds. It must be borne in mind, however, that evaluating only crop tissue by either of these techniques can still fail to identify ABV-positive cases.
The RT-PCR data in the current study reinforce the hypothesis that the distribution of virus is much broader than the clinical signs or histopathologic lesions would suggest. In addition, birds with histologic evidence of PDD can be free of evident clinical symptoms in many histologically involved sites. For example, many of the birds in the present study with histopathologic lesions in the crop or proventriculus had no clinical symptoms of gastrointestinal tract dysmotility or wasting. It thus appears that while the presence of classical clinical signs is a strong indicator for the presence of PDD, the absence of clinical signs, even over a period of up to 3 years (as seen in the current study with birds from the research colony), cannot be used to indicate freedom from disease and/or infection. The possibilities of latent infection and subclinical PDD have long been suspected, and their existence is strongly supported by these findings.
Five out of the 7 birds (birds 1-3, 15, and 16) that had shown neurologic signs at some point in their clinical history were histopathologically diagnosed with PDD and were positive for ABV by both RT-PCR and IHC. All 5 birds had histopathologic lesions in the central nervous system, and 4 out of 4 examined had intranuclear staining in the neurons in brain and spinal cord on IHC. The 2 other birds (birds 7 and 12) that had shown neurologic signs did not have histologic lesions in the central nervous system in sections examined and were negative for ABV by IHC. Bird 7 was, however, RT-PCR positive in sections of the brain as well as adrenal gland. Focal lesions elsewhere in the central nervous system cannot be excluded in these cases. PDD remains a cause of neurologic clinical signs in psittacine birds, but other etiologies, including bacterial and other viral encephalitides, toxicity, neoplasia, and aberrant parasite migration, must always be considered for these cases.
An unexpected finding was the presence of a lymphoplasmacytic interstitial infiltrate in the kidneys of 7 out of 8 birds diagnosed histopathologically with PDD, as compared with 2 out of 8 birds without the disease. Among the 8 birds with histopathologic evidence of PDD, 6 out of 7 were positive for ABV RT-PCR in their kidney samples (birds 1-5 and 16). Five of these birds (birds 1-5) also had widespread positive ABV IHC staining in the proximal and distal tubule nuclei, and all 5 had RT-PCR-positive cloacal swabs. This could reflect viral residence in the kidney resulting in an interstitial nephritis, as well as the possibility of renal excretion of virus. Bird 7, although negative for PDD on histopathology and for ABV on IHC, was ABV RT-PCR positive on various tissues including kidney, but the cloacal swab was negative by RT-PCR. Segmental distribution of histopathologic lesions and of the antigen, intermittent viral shedding, or simply another cause of the lympho-plasmacytic infiltrate could help explain the inconsistent findings in this bird.
In conclusion, the findings strongly support a causal association between infection with Avian bornavirus and PDD in psittacine birds. An excellent statistical association was found between histopathologic diagnosis of the disease and ABV-specific RT-PCR and IHC. The current study described a protocol for immunohistochemical analysis of ABV in psittacine tissues, and by using RT-PCR and IHC, has also demonstrated the cell and tissue tropism of ABV infection in psittacine birds. The sensitivity and specificity of ABV-specific IHC and RT-PCR for diagnosis of PDD in tissues of psittacine birds have also been discussed in the present study. However, further research including infectivity and transmission studies are warranted to fulfill Koch's postulates and conclusively establish ABV as the sole causative agent of PDD, and to determine the exact pathogenesis of ABV infection, including the actual mode and routes of viral shedding in different species of psittacine birds.
Acknowledgements
The authors are indebted to Linda Groocock for taking excellent care of our research parrots over the years and gratefully acknowledge the technical assistance received from Janet Swinton and Susan Lapos with the ABV RT-PCR and ABV IHC, respectively. The authors also thank Barb Jefferson for her help with processing and storage of specimens early on in this study. Research support was provided for this work by the Ontario Veterinary College Pet Trust Fund, the Canadian Parrot Symposium, HARI, STOP PDD, and many dedicated parrot owners and associations of parrot owners and fanciers. Amy Kistler, Don Ganem, and Joseph DeRisi were supported by the Howard Hughes Medical Institute.
Footnotes
a.
RNA later®, Ambion Inc., Austin, TX.
b.
Forane®, Baxter International Inc., Deerfield, IL.
c.
Hospira Inc., Lake Forest, IL.
d.
Superdex™ 200, GE Healthcare Technologies, Piscataway, NJ.
e.
Amicon Ultra-15, Millipore, Billerica, MA.
f.
Pacific Immunology, Ramona, CA.
g.
Dako Autostainer, Dako Canada Inc., Mississauga, Ontario, Canada.
h.
Dako Canada Inc., Mississauga, Ontario, Canada.
i.
UltraVision ONE, Lab Vision Products/Thermo Fisher Scientific, Fremont, CA.
j.
Vector Laboratories Inc., Burlington, Ontario, Canada.
k.
MagMAX™-96 kit, Applied Biosystems Inc., Foster City, CA.
l.
Multitrans System, Starplex Scientific Inc., Etobicoke, Ontario, Canada.
m.
One-Step RT-PCR Kit, Qiagen Inc., Mississauga, Ontario, Canada.
n.
SeqMan module of Lasergene®, DNASTAR Inc., Madison, WI.
o.
Stata Intercooled 9.0, Stata Corp., College Station, TX.