
Abstract
Between 2001 and 2007, several outbreaks of disease associated with Border disease virus (BDV) infection were reported in the central Pyrenees (northeast Spain) and were associated with a major reduction in chamois (Rupicapra pyrenaica) populations. At the same time, wild boars (Sus scrofa) from the same area were found to be seropositive to this pestivirus, without showing clinical signs. The present study examines the susceptibility of domestic swine and the course of the infection with a BDV strain isolated from naturally infected chamois. Twenty pigs were inoculated with 1 × 107 TCID50 (50% tissue culture infective dose) by oronasal route, and 16 control pigs received Eagles sterile Minimal Essential Medium. Serologic (enzyme-linked immunosorbent assay and virus neutralization test) and reverse transcription polymerase chain reaction assays were performed on serum samples obtained at 0, 3, 7, 10, 14, 21, and 31 days postinoculation (dpi). All infected pigs were viremic from 3 to 14 dpi. After 14 dpi, all infected animals developed an antibody response against the homologous virus. Clinical signs or histologic lesions were not observed in inoculated pigs. The present work demonstrates the susceptibility of domestic swine to a BDV strain of chamois origin.
Keywords
Introduction
Pestiviruses (family Flaviviridae) are single-stranded, positive-sense RNA viruses classified traditionally according to the species of origin. Under this classification, genus Pestivirus includes 4 species: Bovine viral diarrhea virus 1 and 2 (BVDV-1 and BVDV-2, respectively), which affect cattle; Border disease virus (BDV), which infects small ruminants; and Classical swine fever virus (CSFV), which affects pigs and wild boar (Sus scrofa). 3 Three tentative pestivirus species isolated from giraffe, reindeer, 1,2 and fetal calf serum 25 have additionally been proposed.
Interspecies transmission of pestiviruses among several domestic species has been reported previously. Bovine viral diarrhea virus 1 and 2 virus infects not only cattle but also sheep, pigs, and goats 10,20 Classical swine fever virus experimentally infects cattle and goats 13,26 ; and BDV has been isolated from several domestic species such as sheep, goat, cattle, and swine (Rosell R, Pujols J, Muñoz I, et al.: 2008, Antigenic and molecular characterization of border disease virus isolated from pigs. The 7th European Society of Veterinary Virology Pestivirus Symposium, September 16–19, Uppsala, Sweden). 3,5,19 Cross-species transmission of pestiviruses between wildlife and domestic livestock, and the ecology of transmission, is a subject of recent concern. While some investigators theorize that an independent cycle of pestivirus infections occurs in wildlife, 8,9 others describe transmission of pestiviruses from wild animals to susceptible domestic animals on pastures. 15,18 Moreover, pestivirus isolates have been reported in several wild ruminants. 9,11,30
Clinically, BDV infection of sheep is characterized by reproductive disease. 17 Acute infection of immunocompetent animals is usually subclinical, and viremia is transient and difficult to detect. Severe border disease epizootics are unusual but, under some conditions, a severe hemorrhagic syndrome has been reported in sheep. 4,28
Pyrenean chamois (Rupicapra pyrenaica) is a small mountain ruminant belonging to the family Bovidae and subfamily Caprinae. During 2001 and 2002, an outbreak of a disease associated with BDV infection 9,11 occurred in the central Pyrenees, which resulted in a 42% reduction in the chamois population in the affected area. 15 Classical signs of illness associated with this virus in chamois outbreaks were cachectic condition of the animals, presence of pallid mucous membranes due to a low red blood cell count in comparison with reference values, alopecia with skin hyperpigmentation, and several degrees of neurologic alterations such as depression, weakness, and difficulty in moving. Since then, additional outbreaks of severe BDV disease in Pyrenean chamois have been reported in the Catalan Pyrenees. 16
During the same period, wild boars from the same area were found to be seropositive to CSFV as determined by enzyme-linked immunosorbent assay (ELISA; Rosell R, Cabezón O, Mentaberre G, et al.: 2008, Serum antibodies directed against pestivirus in wild boar (Sus scrofa) in Catalonia (NE Spain). The 7th European Society of Veterinary Virology Pestivirus Symposium, September 16–19, Uppsala, Sweden). The antibodies were determined to be due to BDV infections by a comparative virus neutralization (VN) test. If pigs were infected with chamois BDV, the presence of cross-reactive antibodies to CSFV detected by the CSFV ELISA would pose a diagnostic challenge for the control of classical swine fever. Therefore, the objective of the present study was to assess the susceptibility of the domestic pig to infection with a BDV strain isolated from naturally infected chamois, as well as to monitor the dynamics of the BDV infection and antibody response in experimentally inoculated pigs for a 1-month period after inoculation.
Material and methods
Animals
Thirty-six 1-month-old female Large White X Landrace pigs were housed in the level-3 biosafety facilities of the experimental station of Centre de Recerca en Sanitat Animal (Universitat Autònoma de Barcelona, Spain). Sera from all pigs were tested for the presence of pestiviruses and antibodies using a commercial ELISA, a VN test, and reverse transcription polymerase chain reaction (RT-PCR) 29,31 test. All analyzed samples were negative.
Virus and inoculum preparation
The BDV strain used to infect the animals was isolated from the spleen of a diseased chamois found in the Pyrenees in 2005 that showed classic signs of illness associated with BDV infection. 15 The spleen sample was homogenized in Eagles sterile Minimal Essential Medium (EMEM; 1 g of tissue + 9 ml EMEM) 6% supplemented with penicillin 10,000 U/ml, streptomycin 10,000 μ/ml, and nystatin antibiotics 10,000 U/ml, and tested for pestiviral RNA by RT-PCR. A BDV strain was isolated in the pestivirus-free sheep fetal thymus (SFT-R) cell line. The virus was titrated directly in the SFT-R pestivirus-free monolayer cell line in a 96-well tissue culture plate. Viral replication was monitored by immunoperoxidase monolayer assay (IPMA) 33 with a swine polyclonal pestivirus antibody. Virus titration was performed as described previously, 21 yielding a titer of 1 × 107 TCID50 (50% tissue culture infective dose).
To assess the phylogenetic homology between BDV and the BDV strain isolated in the same and previous outbreaks, amplified DNA from spleen homogenate and virus isolates were purified b and sequenced. Purified amplicons were analyzed with a commercial kit c and a genetic analyzer. c The 5′ UTR (untranslated region) sequence of the virus used in the experimental infection was deposited in GenBank under the name CADI-6 (accession no. AM905923). The phylogenetic tree was calculated by the neighbor-joining method 24 using an automatic root location. To test the reliability of the branches in the tree, a bootstrap analysis of 1,000 replicates was performed by creating a series of bootstrap samples. The viral sequence from the chamois isolate was located in the same BDV-4 genotype described in previous outbreaks in chamois populations (Fig. 1).
Experimental design
After an acclimatization period of 7 days, pigs were randomly assigned to 2 groups reared in separate isolation units. Twenty pigs were inoculated by oronasal natural route with 1 × 107 TCID50 of BDV in 2 ml of EMEM. The inoculum was administered by a combination of nasal catheter (0.5 ml in each nostril), and 1 ml was administered orally. Sixteen pigs were inoculated with 2 ml of sterile EMEM by the same route to serve as negative controls.
Pigs were observed daily for clinical signs. Rectal temperature was monitored 3 times per week, and weight was measured weekly during the experiment. Whole blood was collected at 0, 3, 7, 10, 14, 21, and 31 days postinoculation (dpi). Blood samples were collected from the cranial cava vein, placed in serum collection tubes, and centrifuged at 103 rcf for 15 min. Nasal, oral, and rectal swabs were also collected the same days as blood samples, and resuspended in 2 ml of sterile phosphate buffered saline (PBS) supplemented with antibiotic. Sera samples and swabs were stored at −80°C until tested. Necropsies were performed at 0 dpi (2 control pigs [CP]), 3 dpi (2 CP, 3 inoculated pigs [IP]), 7 dpi (2 CP, 3 IP), 14 dpi (3 CP, 3 IP), 21 dpi (2 CP, 3 IP), and 31 dpi (5 CP, 8 IP). At necropsy, urine swabs and tissue samples (spleen, kidney, brain, lymph node, skin, thyroid gland, intestine, liver, lung, bone marrow, and testicle) were collected. Animal care and study procedures were conducted in accordance with the guidelines of the Good Experimental Practices, under the approval of the Ethical and Animal Welfare Committee of the Universitat Autònoma of Barcelona.
Virologic examination
Viral RNA was extracted directly from 150 μl of sera and swab samples diluted in PBS by a commercial kit d according to the manufacturer's procedures. Reverse transcription PCR was performed to detect pestiviral RNA (5′ UTR region) using previously described pan-pestivirus primers 324 and 326 29,31 and a commercial kit. e In order to verify the homology between the virus detected in infected pigs and the inoculum, RNA present in a positive serum sample was amplified, purified, and sequenced as described above.
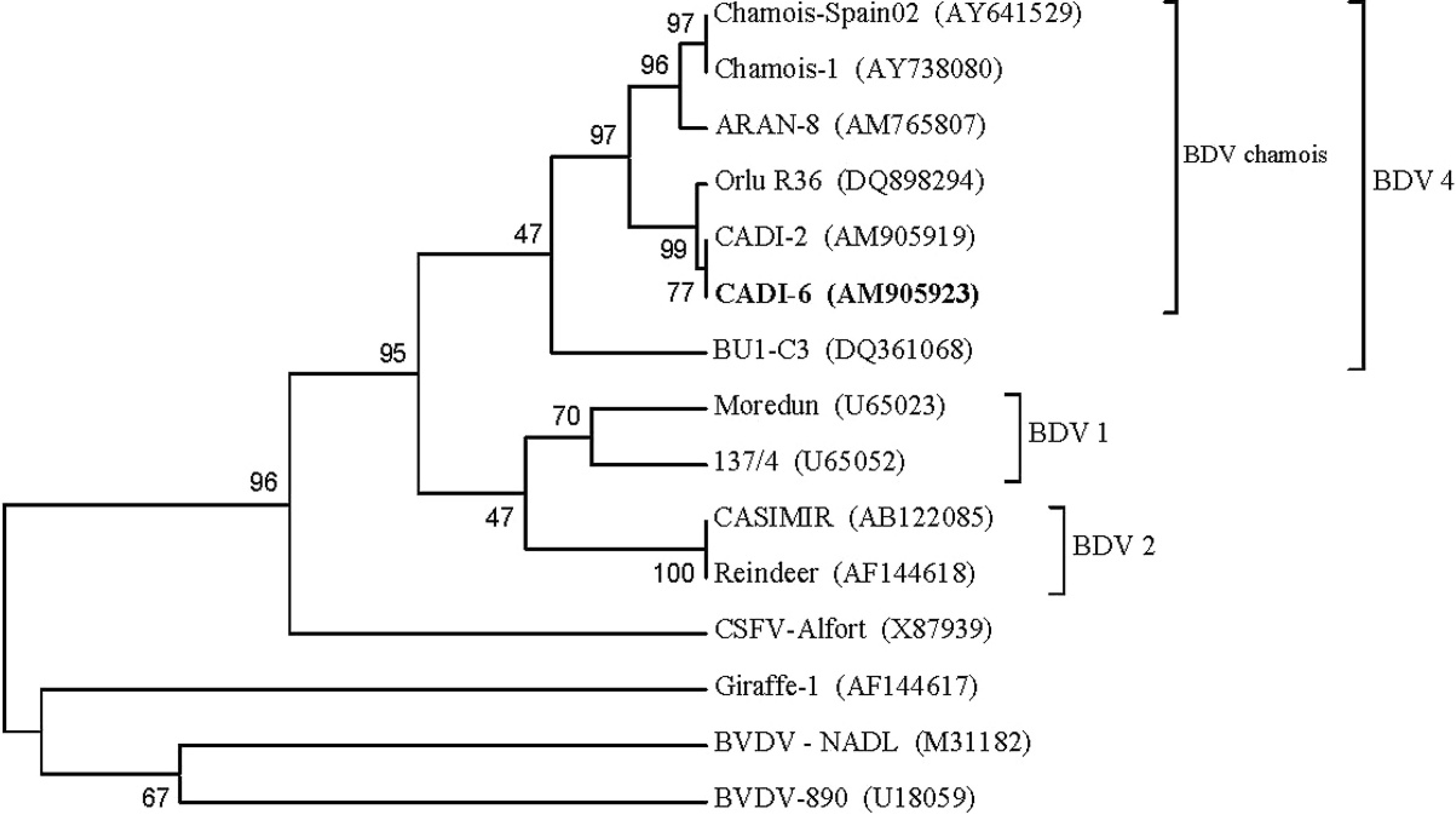
Unrooted neighbor-joining phylogenetic tree based on the 5′-untranslated region sequence among pestiviruses. Chamois strain CADI-6 AM905923 used to experimentally infect swine was enclosed in a differentiated group into Border disease virus (BDV) serotype 4. The numbers on the branches indicate the bootstrap values (in percent; 1,000 replicates). GenBank accession numbers are indicated in brackets.
Serologic tests
Three commercial ELISA tests were used to detect anti-pestivirus antibodies in sera from all animals and sample collection days. The 3 ELISA tests were performed following the procedure described by the manufacturer. The POURQUIER® BVD/MD-BD P80 antibody blocking ELISA kit a (hereafter, Pourquier ELISA) detected antibodies against the p80 protein of BVDV and BDV. The inhibition percentage in individual sera samples was calculated as follows: inhibition percentage = ODS/ODN × 100, where OD is the optical density of the sample (S) and of the negative (N) control. Samples with inhibition percentage ≥50%, 40–50%, and ≤40% were considered negative, weakly positive, and positive for BDV antibodies, respectively. The reaction of the Pourquier ELISA was considered valid when the negative control had a minimum mean OD450 (optical density at 450 nm) value of 0.800 and the percentage of competition of the positive control was less than 20%.
The HerdChek® CSFV antibody ELISA f (hereafter, HerdChek ELISA) and the Ceditest® CSFV 2.0 ELISA g (hereafter, Ceditest ELISA) detected antibodies against the E2 protein of CSFV. The inhibition percentage in individual sera samples in the HerdChek ELISA was calculated as follows: blocking percentage = ((NCX − sample A450)/NCX) × 100, where NCX is the negative control mean. Samples with blocking percentage ≥40%, 30–40%, and ≤30% were positive, weakly positive, and negative, respectively. The reaction of the test was considered valid when the negative control had a minimum mean OD450 value of 0.50 and the percentage of competition of the positive control was more than 50%. The inhibition percentage of the sera samples in the Ceditest® ELISA was calculated as inhibition percentage = 100 − [(ABS S/ABS N) × 100], where ABS is absorbance. Samples with blocking percentage <40% and ≥40% were considered negative and positive, respectively. The reaction of the test was considered valid when the negative control ABS mean was >1.0 and the positive control inhibition percentage was >80%.
A VN test was used for the detection and quantification of anti-BDV antibodies in serum of inoculated and control pigs. The VN test was performed against 4 pestiviruses: the homologous BDV (CADI-6), and the reference pestiviruses CSFV-Alfort, BDV-137/4, and BVDV-NADL. Virus neutralization test was performed following the procedure described in the Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 33 Titers of 1:10 or higher were considered as positive. Viral replication was monitored by IPMA 33 with a polyclonal pestivirus antibody.
Histopathological studies
All tissue samples collected during necropsy (spleen, kidney, brain, lymph node, skin, thyroid gland, intestine, liver, lung, bone marrow, and testicle) were fixed by immersion in formalin and subsequently embedded in paraffin. Tissue sections were cut at 4 μm, stained with hematoxylin and eosin, and examined for the potential presence of lesions.
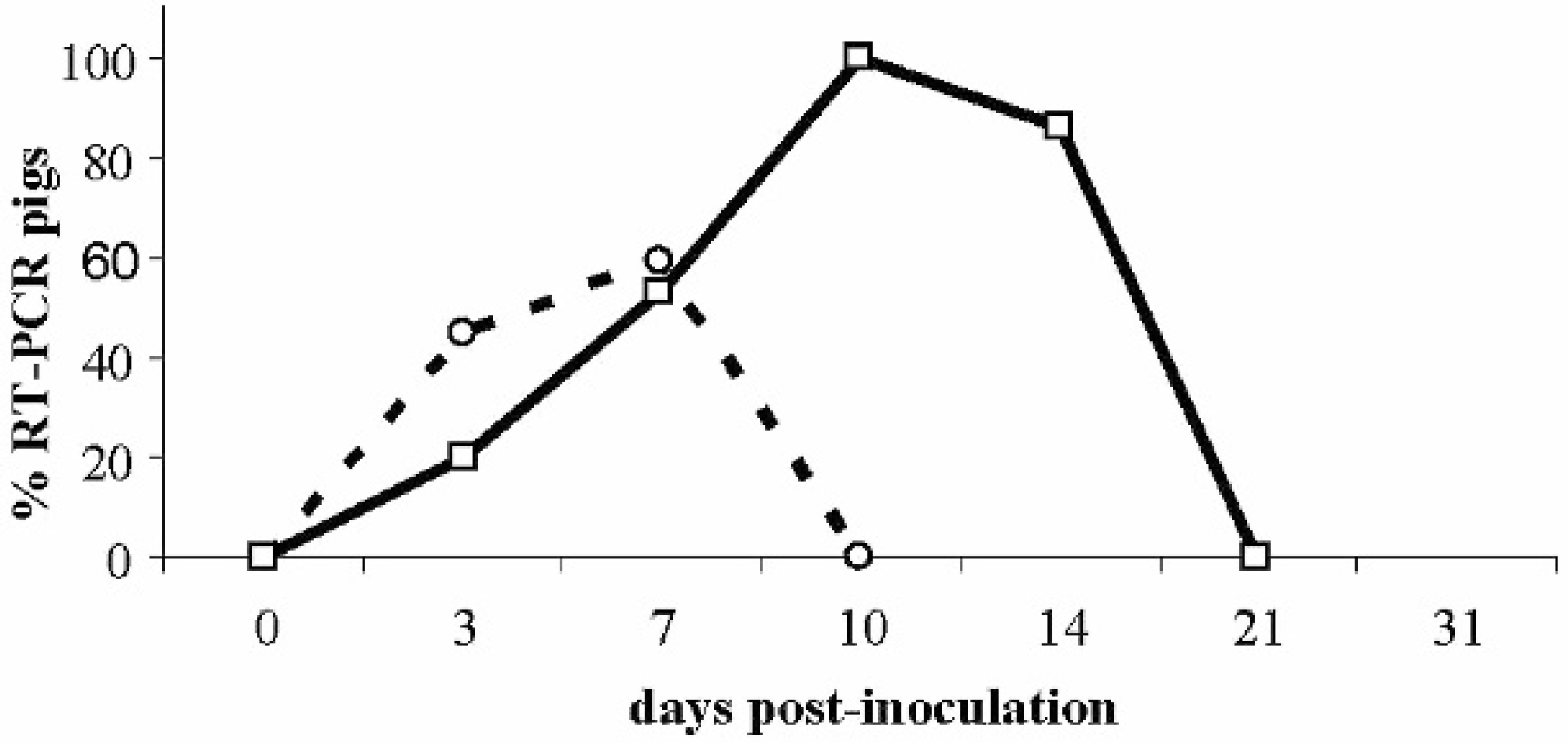
Infection dynamics of inoculated piglets. Percentage reverse transcription polymerase chain reaction (RT-PCR) pigs positive in oral or nasal swabs (◯) and in serum (□).
Statistical analysis
Differences in mean temperatures between inoculated and control groups were analyzed by analysis of variance using proc mixed. h P values less than 0.05 were considered significant.
Results
No clinical signs were observed during the entire experimental period. There were no statistical differences in the mean temperatures between inoculated and control pigs during the infection period (P > 0.05).
At postmortem examination, gross or histopathologic lesions were not found in any inoculated or control pig. Prevalence of viremic pigs at different dpi was 20% (4/20) at 3 dpi, 29.41% at 7 dpi (5/17), 100% at 10 dpi (14/14), and 85.71% at 14 dpi (12/14; Fig. 2). The mean duration of viremia was 7.9 days (±3.36 standard deviation [SD]).
Border disease virus was detected in oral and/or nasal swabs from 3 dpi in 9 out of 20 pigs (45%) to 7 dpi in 10 out of 17 pigs (58.82%) and was not detected in rectal or urine swabs from any inoculated pig. Sera and all swab types from control pigs at all sampling points were negative for BDV by RT-PCR. The sequence of a BDV strain isolated from 1 inoculated pig at 10 dpi had 100% identity with the BDV strain used for inoculation.
Antibodies to BDV were detected in 1 out of 14 pigs (7.14%) at 14 dpi, 3 out of 11 pigs (27.27%) at 21 dpi, and 7 out of 8 pigs (87.5%) at 31 dpi using the Pourquier ELISA. The other 2 ELISA tests to detect antibodies to CSFV detected CSFV antibody-positive serum samples from only 1 pig at 21 and 31 dpi (Table 1). Neutralizing antibodies to BDV were detected in serum samples from almost all pigs at 14 dpi and in all pigs at 21 dpi. Maximum titers were observed at 31 dpi (minimum 80, maximum 640; Table 1). Higher titers of neutralizing antibodies were observed against the challenging BDV strain compared with other pestivirus strains (BDV-137/4 and BVDV-NADL) at 21 and 31 dpi. Neutralizing antibodies to CSFV-Alfort were not detected in inoculated animals at any point during the experiment.
Discussion
To the authors' knowledge, this is the first experimental infection of swine with BDV isolated from a wild ungulate. Many experimental infections with pesti-viruses, mainly CSFV and BVDV, have been performed in swine, but there are few studies on experimental infections with BDV viruses with information on the clinical signs and pathogenesis (Depner KR, Schirrmeier H: 2002, Inoculation of pigs with BDV or BVDV-2 and subsequent challenge with CSFV. In: Abstract of the 5th European Society of Veterinary Virology Pestivirus Symposium, Cambridge, England). 13
Virus neutralization titers for sera of inoculated pigs.*
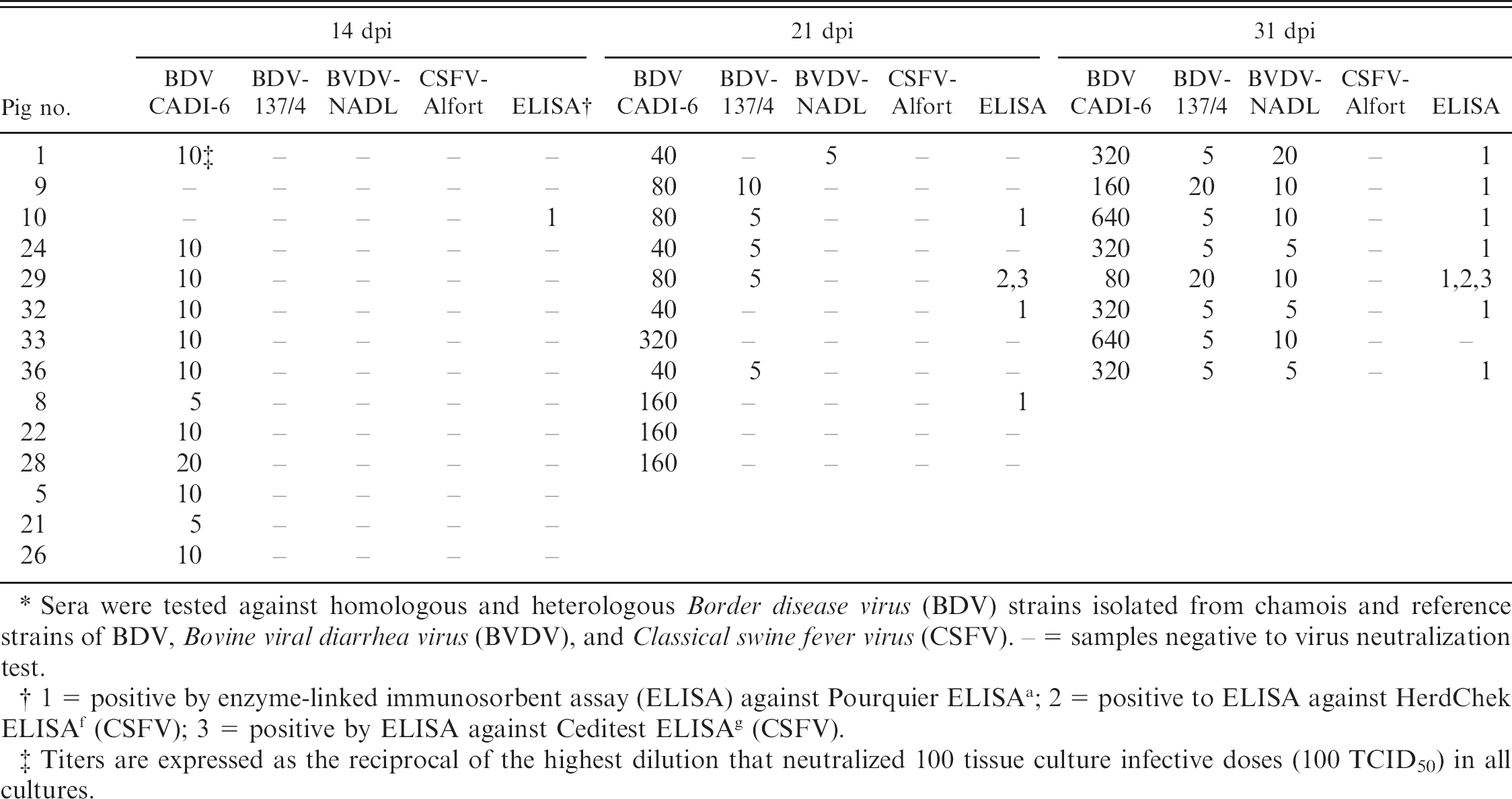
Sera were tested against homologous and heterologous Border disease virus (BDV) strains isolated from chamois and reference strains of BDV, Bovine viral diarrhea virus (BVDV), and Classical swine fever virus (CSFV). − = samples negative to virus neutralization test.
1 = positive by enzyme-linked immunosorbent assay (ELISA) against Pourquier ELISA a ; 2 = positive to ELISA against HerdChek ELISA f (CSFV); 3 = positive by ELISA against Ceditest ELISA g (CSFV).
Titers are expressed as the reciprocal of the highest dilution that neutralized 100 tissue culture infective doses (100 TCID50) in all cultures.
The pathogenesis of pestivirus infection in pigs is highly variable depending on the strains involved. Classical swine fever virus infection is associated with clinical signs and disease. 33 However, BVDV infection in pigs leads to mild or unapparent disease. 6,7,27,32 Pigs inoculated by the oronasal route with the CADI-6 BDV isolate did not develop clinical disease, and statistical differences between mean rectal temperatures of control and inoculated groups was not observed. The results support previous data that BDV is unlikely to induce clinical disease in pigs. 12
Classical swine fever virus, BVDV, and BDV are shed by infected animals in oronasal and lacrimal secretions, urine, feces, and semen. 33 The presence of virus in oral and nasal swabs between 3 and 7 dpi in inoculated pigs of the present study is likely to represent an active secretion of virus through these 2 routes, as well as viral replication at the inoculation region.
The 4 species of pestiviruses infect swine; however, the period of viremia varies depending on the species and strain of the virus. 14,33 In CSFV infections, viremia usually starts between 2 and 15 dpi, 33 while in BVDV infections, viremia is detected within the first week of the infection. 6–8,27,32 The results of experimental BDV infections in pigs indicated low or undetectable virus in the blood (Depner KR, Schirrmeier H: 2002, Inoculation of pigs). In the present study, BDV-inoculated pigs were positive for the presence of virus in sera from 3 to 14 dpi, which is a longer period of viremia than that previously reported. The reasons for this relatively long-term viremia are unknown but might be due to the high inoculating dose and/or a replication characteristic of the specific BDV used in the challenge.
The serologic data indicate that all inoculated pigs were infected with BDV and that an antibody response was developed against the homologous virus. The humoral immune response to pestiviruses has been previously reported to be highly variable in swine with respect to onset relative to infection. Some authors reported detectable BVDV antibody levels 5 weeks after infection in sows, while others showed evidence of seroconversion within 3 weeks of the inoculation of pigs with a BDV strain. 12,32 In the present study, swine seroconverted by 14 dpi, and antibodies were maintained until the end of the study (31 dpi), with titers as high as 640 in some pigs by ELISA and VN tests. The detection of antibodies by VN test coincided with the end of RT-PCR-positive serum samples. It is important to note that higher titers to the BDV challenge were found compared with the other pestivirus used in the VN test. There was some cross-reactivity with other BDV and BVDV strains detected by the VN test; however, no CSFV-specific antibodies were detected in the serum of any of the infected pigs.
The 3 commercial ELISA tests were much less sensitive than the VN test in detecting antibodies. The detection of antibody-positive pigs with the Pourquier ELISA increased from 14 dpi (7.7%) through the end of the study at 31 dpi (87.5%). One pig out of 14 that was detected as seropositive by the Pourquier ELISA at 14 dpi was seronegative by the VN test. The most likely explanation for this situation is that this serum was a false-positive sample detected by the ELISA. Of interest, a single pig was antibody positive at 21 and 31 dpi by both the HerdChek and Ceditest ELISAs. This animal was positive by the Pourquier ELISA at 31 dpi. Detection of BDV antibodies in swine serum by the HerdChek and Ceditest ELISAs could present a diagnostic challenge for CSFV serodiagnosis. The current study suggests that the VN test is the best reference laboratory test to discriminate antibodies in the various species affected by pestiviruses. Therefore, currently circulating pestivirus strains should be included in routine VN tests.
In conclusion, the present study confirms that the BDV strain isolated from naturally infected chamois can also infect the domestic pig. The infected animals developed a relatively long viremia with seroconversion 2 weeks after inoculation. The results support the horizontal transmission of BDV between chamois and wild boar suspected to have occurred during the BDV epizootic in chamois (Rosell R, et al.: 2008, Serum antibodies directed against pestivirus in wild boar). The current study agrees with previous studies that indicate transmission of BDV from ruminants to swine occurs when both species are in close contact. 12,22 Further, the findings suggest that this BDV strain could be transmitted from wild boar to domestic pigs. Taking into account that the wild boar is a reservoir of CFSV, 23 the potential transmission of BDV from the wild to the domestic pig could result in a diagnostic challenge due to the cross-reactivity of the BDV antibodies for CSFV ELISA based tests.
Acknowledgements
The authors would like to give special thanks to the rangers and staff of the Cadí National Hunting Reserve for the collection of samples and the capture of the diseased chamois. Research was supported by the Direcció del Medi Natural del Department de Medi Ambient de la Generalitat de Catalunya. The study was supported by Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), reference project FAU 2006-00007-C02-02.
Footnotes
a.
Institut Pourquier SAS, Montpellier, France.
b.
Minelute Gel Extraction Kit, Qiagen Inc., Valencia, CA.
c.
BigDye® Terminator v.3.1 Cycle Sequencing Kit, 3130xl Genetic Analyzer; Applied Biosystems, Foster City, CA.
d.
NucleoSpin® Viral RNA Isolation, Macherey Nagel GmbH & Co. KG, Düren, Germany.
e.
OneStep PCR kit, Qiagen Inc., Valencia, CA.
f.
IDEXX Laboratories Inc., Westbrook, ME.
g.
Cedi-Diagnostics B.V., Lelystad, The Netherlands.
h.
SAS Inc., Cary, NC.