
Abstract
Pyrenean chamois (Rupicapra pyrenaica) populations of the central and eastern Pyrenees have been affected by severe outbreaks associated with Border disease virus (BDV) since 2001. Eight Pyrenean chamois (7 males and 1 female) from 1 to 8 years of age with clinical signs consistent with BDV infection were studied. At necropsy, whole blood, tissue samples (skin, brain, prescapular lymph node, thyroid gland, lung, liver, spleen, kidney, small intestine, bone marrow, and testicle), urine, and nasal, oral, and rectal swabs were obtained. The fetus from a pregnant female was also studied. Reverse transcription polymerase chain reaction (RT-PCR) was used to detect the virus in all samples, and virus isolation was performed. Sera and tissue samples were positive to RT-PCR, and the virus was isolated from all chamois. The nasal, oral, and rectal swabs and urine samples were RT-PCR positive in 100%, 85.71%, 71.43%, and 100% of chamois, respectively, confirming the excretion of the virus via these 4 routes. In addition, sera were tested for BDV antibodies using enzyme-linked immunosorbent assay and seroneutralization techniques, with negative results. Sequence analysis of the 5′ untranslated region in 7 of the chamois confirmed that the virus is grouped into the BDV-4 genotype, the same BDV previously described in Pyrenean chamois. To the authors' knowledge, this is the first study of naturally infected Pyrenean chamois, providing evidence that infected animals shed BDV through nasal, oral, fecal, and urinary excretion routes.
Keywords
Border disease virus (BDV; family Flaviviridae, genus Pestivirus) is the cause of border disease (BD) in sheep and goats. The disease is characterized by reproductive manifestations such as barren ewes, abortions, stillbirths, and weak lambs that can be persistently infected (PI). Infection between days 50 and 60 of gestation may result in the birth of immunotolerant PI lambs that shed the virus throughout life and constitute the most important source of BDV in ruminant populations. 17 In these PI lambs, BDV is widespread in all organs. 10 Severe outbreaks of BD in small ruminant populations are unusual, but several epizootics have been described in sheep. 3,4,16
The Pyrenean chamois (Rupicapra pyrenaica) is a high-mountain small ruminant distributed in the Apennines (Italy), the Cantabrian Mountains (Spain), and the Pyrenees (Southern France, Andorra, and Northern Spain). In 2001 and 2002, an outbreak of disease associated with BDV infection 1,5,6 was detected in the central Pyrenees, resulting in a 42% reduction in the chamois population in the affected area. 8 Since that date, further severe outbreaks of BDV infection in Pyrenean chamois have been reported 9 in the Catalan Pyrenees.
Although BDV infection has been studied widely in domestic animals, there are no studies on virus excretion and virus distribution in organs from diseased chamois and other wild animals. The aim of the current study was to investigate the distribution and quantification of virus in the organs of affected chamois and to determine the routes of excretion (nasal, oral, fecal, and urinary) of the virus.
In 2006, 8 Pyrenean chamois (7 males and 1 pregnant female) from 1 to 8 years of age with clinical signs consistent with BDV infection were studied. All chamois were emaciated and in terminal stages of illness. All animals showed areas of alopecia with epidermal hyperpigmentation, depression, weakness, and difficulty in moving. The chamois were captured in the Cadí National Hunting Reserve (Eastern Pyrenees, Catalonia, Spain) during a severe outbreak of the disease. 9 The animals were sent to the wildlife facilities of the Veterinary Faculty at the Universitat Autònoma de Barcelona (Bellaterra, Barcelona, Spain) and were euthanized in accordance with animal care guidelines. Complete necropsies were performed on all animals.
Before euthanasia, a 20-ml blood sample was taken from the jugular vein of each animal and placed in a serum collection tube. Nasal, oral, and rectal swabs were also collected. At necropsy, urine and tissue samples (skin, brain, prescapular lymph node, thyroid gland, lung, liver, spleen, kidney, small intestine, bone marrow, and testicle) were obtained. A sample from the spleen was taken from the fetus of a pregnant chamois, which was estimated to be in the last 7 weeks of gestation. Separate sterile instruments were used for each animal and during sampling in order to avoid cross-contamination. Nasal, oral, and rectal swabs were homogenized in 2 ml of sterile phosphate buffered saline supplemented with antibiotic. Blood samples from serum collection tubes were centrifuged at 103 × g for 15 min and sera were obtained. Tissue samples were homogenized in sterile Eagle Minimal Essential Medium (EMEM; 1 g of tissue + 9 ml EMEM) 6% supplemented with antibiotics (penicillin: 10,000 U/ml; streptomycin: 10,000 μg/ml; nystatin antibiotics: 10,000 U/ml). All samples were stored at −20°C until they were tested.
Amplification of viral RNA was conducted by reverse transcription polymerase chain reaction (RT-PCR). Viral RNA was extracted directly from tissue homogenates, swabs, and sera using a commercial kit. a Reverse transcription PCR was performed to detect pestiviral RNA (5′-untranslated region [UTR] region) using previously described primers 324 and 326 17,19 and a commercial kit. b
In order to quantify the virus from the samples of the studied animals, oral, nasal, and rectal swab samples; sera; urine; and tissue homogenates were titrated directly on sheep fetal thymoid pestivirus-free monolayer cell line in a 96-well cell culture plate. Viral replication was monitored using the immunoperoxidase monolayer assay with an in-house polyclonal antibody. 20 Virus titrations were performed as described previously. 14
Serum samples from all 8 animals were tested for the presence of Pestivirus antibodies against p125/p80 protein with a commercial sandwich enzyme-linked immunosorbent assay test (ELISA), c following the manufacturer's instructions. A virus neutralization test (VNT) 20 was also performed on the sera. Titers of 1:10 or higher were considered to be positive for anti-BDV neutralizing antibodies.
Analysis of the sequence of the 243–base pair 5′UTR fragment generated by RT-PCR was performed on all of the chamois studied. Amplified DNA from 1 sample from each animal was purified d and sequenced. Purified amplicons were analyzed with a commercial sequencing kit e and an automated electrophoresis system. e The phylogenetic tree was made by the neighbor-joining method 15 using an automatic root location. To test the reliability of the branches in the tree, a bootstrap analysis of 1,000 replicates was performed by creating a series of bootstrap samples. Pestivirus genome was detected by RT-PCR in sera (8/8; 100%); oral (6/7; 85.7%), nasal (7/7; 100%), and rectal (5/7; 71.4%) swabs; urine (7/7; 100%); and in all of the tissue samples of all studied chamois, including the spleen from the fetus (Table 1).
The results of isolation and titration of virus in different samples are listed in Table 1. Briefly, BDV was isolated from sera (8/8; 100%); nasal (4/7; 57.1%), oral (2/7; 28.6%), and rectal (1/7; 14.3%) swabs; skin (6/7; 85.7%); and from almost all of the tissue samples tested (100%), except for the small intestine (2/6; 33.3%). Infectious BDV could not be isolated from urine samples and fetal spleen. The highest concentration of infectious virus was present in 6 out of 8 serum samples with titers higher than 6.5 50% tissue culture infective dose/ml. Both ELISA and VNT for Pestivirus antibodies produced negative results.
Genetic analysis of the 5′UTR of the viral genome showed that the BDV isolates obtained belong to the BDV-4 genotype and are closely related to the BDV isolates previously reported 1,6,8,12 in sick Pyrenean chamois (Fig. 1). The sequences from the present report were deposited in the GenBank under the following names and accession numbers: CADI-1 (AM905918), CADI-7 (AM905924), CADI-8 (AM905925), CADI-9 (AM905926), CADI-10 (AM905927), CADI-11 (AM905928), CADI-12 (AM905929), and CADI-13 (FN397676).
Pestivirus infection, mostly associated with Bovine viral diarrhea virus, has been reported 18 in many species of wild ruminants. Border disease virus infection with a fatal outcome has only been described in a bison from a German zoo and in free-ranging Pyrenean chamois. 1,2,6,8,9
In sheep with persistent BDV infection, the virus has been demonstrated to exist in almost every organ, indicating a generalized infection. Continuous excretion occurs mainly via nasal secretions and saliva, making the oronasal route the principal route of natural spread of Pestivirus among adult individuals. 7,13 In the diseased chamois of the present study, the presence and isolation of the virus in sera—in all tissues and in all excretion routes—concur with findings of previous sheep studies, confirming that these chamois were in a viremic state of the infection.
The presence of BDV in oral, nasal, and rectal swabs and in urine samples was successfully demonstrated by RT-PCR. However, virus was not isolated in several of those samples, and it was never isolated from the urine samples. These differences might be explained by the fact that the virus titer in these samples was low or below the detection level by virus isolation and by the likelihood that the inactivation of virus probably occurs more rapidly in the urine samples. When virus was isolated in swabs, the titers observed were lower than the titers observed in the organs (Table 1). Nevertheless, detection of BDV by RT-PCR in all swabs and urine samples confirms that diseased chamois excrete significant levels of the virus and that it is most likely that a high horizontal transmission rate of the virus occurs in nature. This is likely one of the factors that could explain the high number of chamois affected in the latest epizootics. 8,9
Reverse transcription polymerase chain reaction (RT-PCR) study and infectious Border disease virus (BDV) titers in sera, swabs, and main organs of 8 Pyrenean chamois (Rupicapra pyrenaica) with Pestivirus infection. *
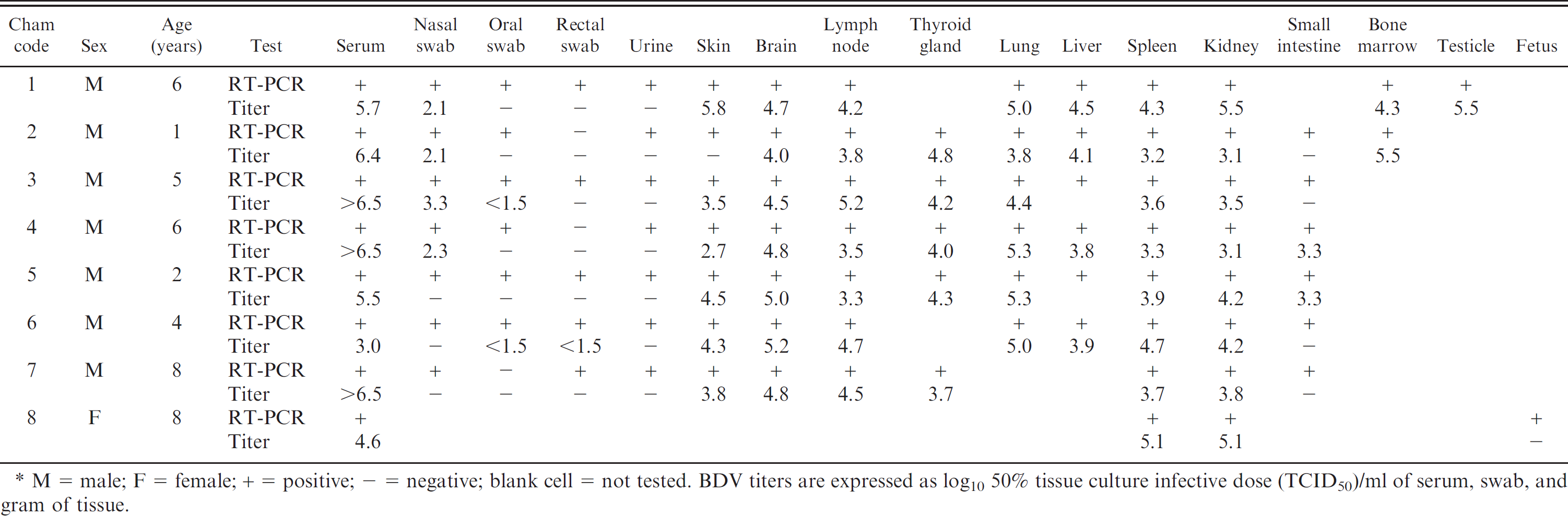
M = male; F = female; + = positive;–= negative; blank cell = not tested. BDV titers are expressed as log10 50% tissue culture infective dose (TCID50)/ml of serum, swab, and gram of tissue.
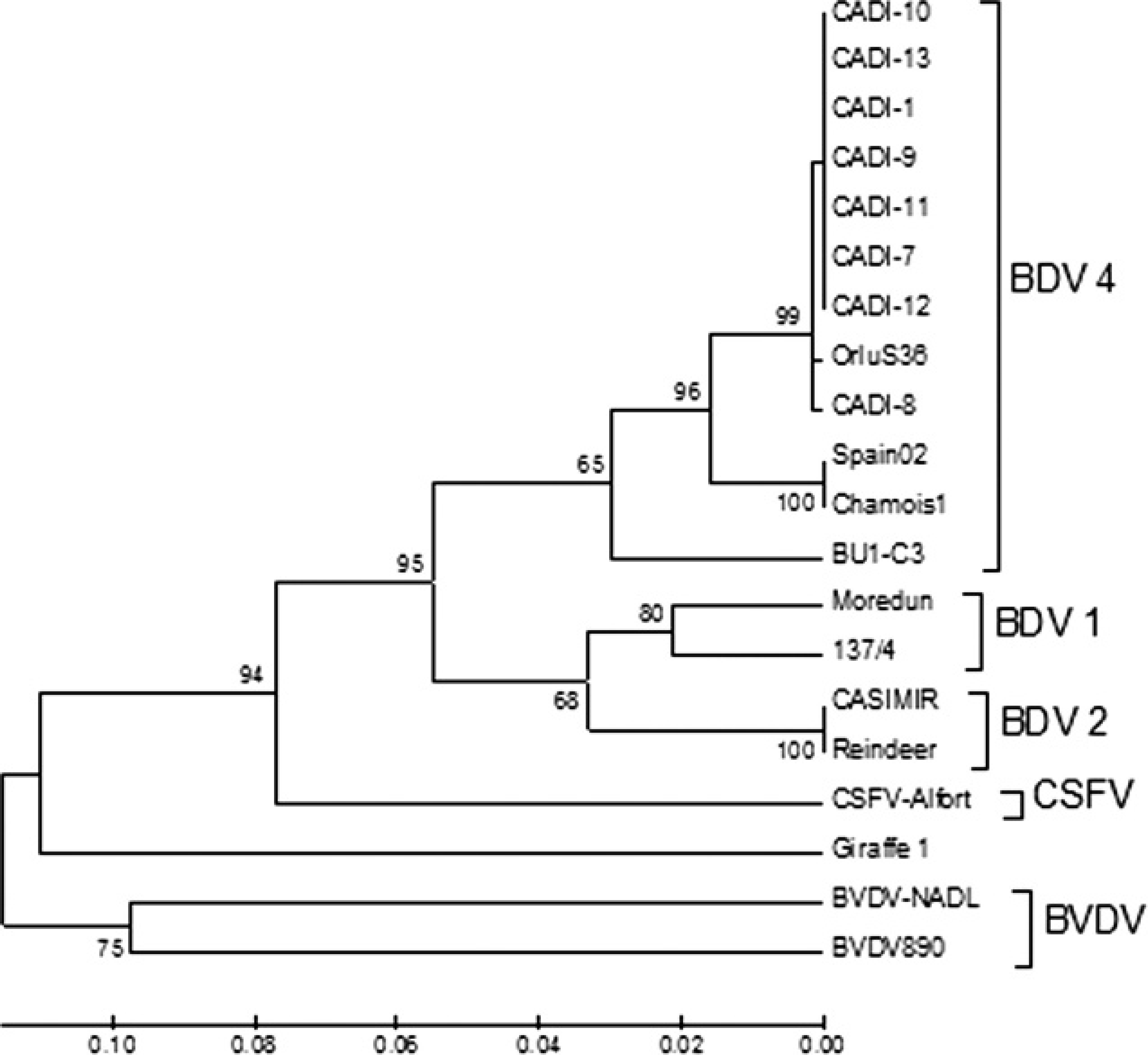
Unrooted neighbor-joining phylogenetic tree based on the 5′ untranslated region (UTR) sequence among pestiviruses. Chamois strains were enclosed in a differentiated group into Border disease virus genotype 4 (BDV-4). The numbers on the branches indicate the bootstrap values (in percent; 1,000 replicates). Sequences of strains were taken from GenBank with the following accession numbers: Chamois-Spain02 (AY641529), Chamois-1 (AY738080), OrluS36 (DQ898293), BU1-C3 (DQ361068), CASIMIR (AB122085), Reindeer (AF144618), 137/4 (U65052), Moredun (U65023), Alfort (X87939), Giraffe1 (AF144617), NADL (M31182), 890 (U18059), CADI-1 (AM905918), CADI-7 (AM905924), CADI-8 (AM905925), CADI-9 (AM905926), CADI-10 (AM905927), CADI-11 (AM905928), CADI-12 (AM905929), and CADI-13 (FN397676).
The detection of BDV in the fetus of a viremic female chamois indicates that vertical transmission of BDV occurs in this species, as in pestivirus-infected domestic ruminants. However, the possible effects of this congenital infection are not known. Confirmation of a persistently infected status requires identification of the virus in 2 separate samples within a minimum of 21 days. 10 The chamois of the present study could not be confirmed as PI because none of the animals could be retested on a second occasion as a result of their clinical condition. The high level of viremia, shedding of BDV, and the lack of antibodies in the chamois would indicate that either they were in the acute phase of the disease or they were PI animals. Unfortunately, it was not possible to determine the time of infection of these animals as a result of their free-ranging life. However, the epidemiological pattern of the outbreaks observed (characterized by a short course and mortality as high as 85%) 8,9 seems to indicate that the outcome of the infection is fatal and that there is a high horizontal transmission rate.
In sheep, acute infections of lambs and nonpregnant mature animals produce no obvious pathological changes. Only slight fever and mild leukopenia were associated with a short-lived viremia, after which virus-neutralizing antibodies appeared in serum. 11 Although high mortality in adult sheep is rare, highly pathogenic BDVs have been reported to cause high mortality in antibody-free populations. 4 Thus, it is reasonable to presume that the same thing could have happened in the outbreaks associated with BDV in the chamois populations of the Central Pyrenees.
Acknowledgements
The authors would like to give special thanks to the rangers and staff of the Cadí National Hunting Reserve for the capture of the diseased chamois. This research was supported by the Direcció del Medi Natural del Department de Medi Ambient de la Generalitat de Catalunya. Roser Velarde is supported by the Spanish Ministerio de Ciencia e Innovación (Subprogram Personal Técnico de Apoyo) and the European Social Fund.
Footnotes
a.
NucleoSpin® Viral RNA Isolation, Macherey Nagel GmbH & Co. KG, Düren, Germany.
b.
OneStep PCR kit, Qiagen Inc., Valencia, CA.
c.
Institut Pourquier SAS, Montpellier, France.
d.
MinElute Gel Extraction Kit, Qiagen Inc., Valencia, CA.
e.
BigDye® Terminator v.3.1 Cycle Sequencing Kit, 3130xl Genetic Analyzer; Applied Biosystems, Foster City, CA.