
Abstract
West Nile virus (WNV) is a public health threat and has caused the death of thousands of North American birds. As such, surveillance for WNV has been ongoing, utilizing numerous biological specimens and testing methods. Nonvascular (i.e., fully grown) feathers would provide a simple method of collection from either dead or live birds of all ages and molt cycles, with presumably less biosafety risk compared with other specimen types, including feather pulp. The current study evaluates WNV detection in nonvascular feathers removed from naturally infected avian carcasses of several species groups. Feathers of corvid passeriforms had the highest sensitivity of detection (64%), followed by noncorvid passeriforms (43%), columbiforms (33%), and falconiforms (31%). Storing feathers for 1 year at −20°C or at ambient room temperature resulted in detection rates of infectious WNV of 16% and zero, respectively, but had no effect on detection rates of WNV RNA in a subset of matched feather pairs (47% for both storage temperatures). The efficacy of WNV detection in nonvascular feathers is greatly enhanced by testing multiple feathers. The advantages of using nonvascular feathers over other tissues may outweigh the relatively low detectability of WNV RNA in certain situations such as remote areas lacking resources for acquiring other types of samples or maintaining the cold chain.
Introduction
West Nile virus (WNV; family Flaviviridae, genus Flavivirus) has killed many thousands of birds since its arrival to North America. 16,17 Numerous laboratory tests are in use for surveillance of WNV in avian carcasses. Targeted tissues include brain, heart, kidney, oropharyngeal swab, and others. 4,7,13,15,17,19–21,24,25,28,34 Avian mortality surveillance is a key component for tracking the distribution of WNV activity throughout North America 31 and has been used for predicting human WNV disease risk in some localities. 6,11,14,22,30 Avian mortality surveillance is primarily a passive system reliant on public participation for reporting carcasses. 3 Most WNV-related avian deaths are detected through testing specimens from singleton carcasses, as with avian influenza surveillance. 18
Several studies of WNV-infected corvid carcasses demonstrated high detection rates of WNV in vascular feather pulp from the base of growing immature feathers. 5,35 Growing feathers are present during periods of molt, which typically occur at least once annually following the breeding season, 8 corresponding with arbovirus transmission activity in northern latitudes. However, in some cases, avian carcasses lack vascular feathers, especially where WNV transmission may occur outside of the avian breeding season. Nonvascular breast feathers from WNV-infected eastern screech-owls (Megascops asio) tested positive for WNV RNA. 26 In addition, collection of nonvascular feathers likely poses a low biosafety risk because the carcass remains intact, carcass handling is minimal, and contact with infectious fluids and wet tissues is avoided. If sufficiently sensitive for WNV detection, nonvascular feathers would provide a simple and consistent method of collection from either dead or live birds of all ages and molt cycles while posing less biosafety risk compared with other specimen types.
Nonvascular feathers have been used for detection of a variety of viruses, such as Avian influenza virus, Marek's disease virus, Beak and feather disease virus, and Avian polyomavirus. 1,10,23,37,38 Therefore, it was hypothesized that nonvascular feathers may be useful for WNV detection in avian mortality surveillance. The current study evaluates WNV detection in nonvascular feathers removed from naturally and experimentally infected bird carcasses of several species groups. Storage conditions and feather processing techniques are also compared. The relative detection probabilities were also examined in the feather vane versus calamus (i.e., tip) versus whole feather. Recommendations for WNV surveillance programs are provided.
Sensitivity of West Nile virus (WNV) RNA detection in nonvascular feathers stored frozen from WNV-infected carcasses by species and species group. *
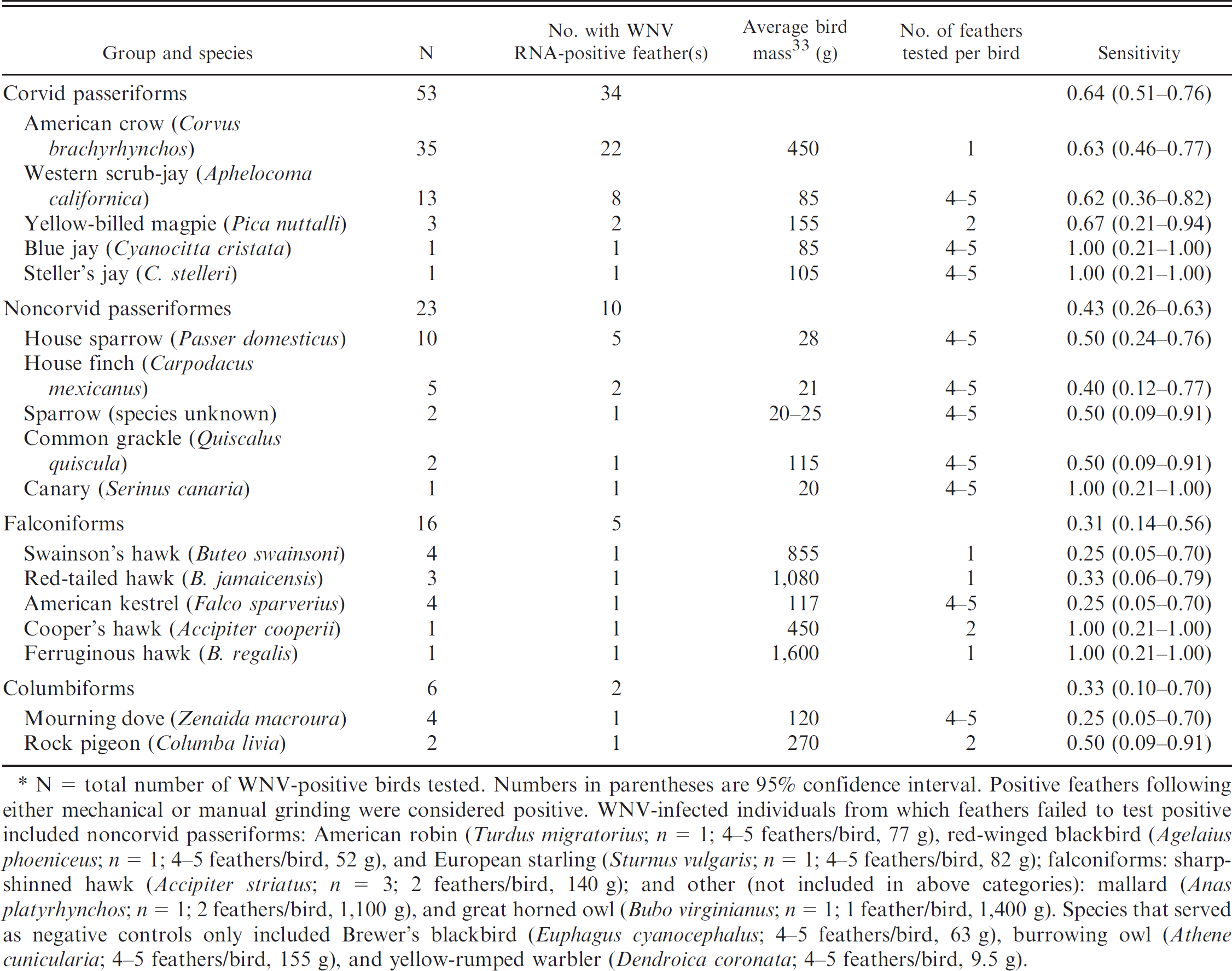
N = total number of WNV-positive birds tested. Numbers in parentheses are 95% confidence interval. Positive feathers following either mechanical or manual grinding were considered positive. WNV-infected individuals from which feathers failed to test positive included noncorvid passeriforms: American robin (Turdus migratorius; n = 1; 4–5 feathers/bird, 77 g), red-winged blackbird (Agelaius phoeniceus; n = 1; 4–5 feathers/bird, 52 g), and European starling (Sturnus vulgaris; n = 1; 4–5 feathers/bird, 82 g); falconiforms: sharp-shinned hawk (Accipiter striatus; n = 3; 2 feathers/bird, 140 g); and other (not included in above categories): mallard (Anas platyrhynchos; n = 1; 2 feathers/bird, 1,100 g), and great horned owl (Bubo virginianus; n = 1; 1 feather/bird, 1,400 g). Species that served as negative controls only included Brewer's blackbird (Euphagus cyanocephalus; 4–5 feathers/bird, 63 g), burrowing owl (Athene cunicularia; 4–5 feathers/bird, 155 g), and yellow-rumped warbler (Dendroica coronata; 4–5 feathers/bird, 9.5 g).
Materials and methods
Avian carcass collection and sampling
Avian carcasses from Colorado and California were sampled by oropharyngeal swab, removal of nonvascular (mature) breast feathers, and harvest of either heart (Colorado) or kidney (California) tissue. Carcasses collected for study were deemed dead <48 hr and were in good condition (e.g., intact, prior to visible decay and appearance of insect larvae), then frozen at −20°C for 1–3 days (Colorado) or shipped overnight on cold packs (California) prior to testing.
Feathers were removed from the carcass and stored in sterile cryovials (Colorado) or Whirl-Pak a bags (California). After feather collection, oropharyngeal swab samples were collected from Colorado birds and California crows by swabbing the oropharyngeal cavity with a cotton-tipped applicator and placing the swab into 1 ml of 15–20% fetal bovine serum prepared in BA-1 diluent (Hank's M-199 salts, 1% bovine serum albumin, 350 mg/l sodium bicarbonate, 100 units/ml penicillin, 100 mg/l streptomycin, 1 mg/l amphotericin B in 0.05 M Tris, pH 7.6). Necropsies were performed on all carcasses from Colorado and noncrow carcasses from California, at which time tissue was removed aseptically with a sterile surgical blade and either placed into cryovials with 1 ml BA-1 diluent and a ball-bearing (BB) pellet (heart) or with 1 ml lysis buffer (kidney). Heart samples (0.5 cm 3 ) were ground in a mixer mill b at 25 cycles/sec for 5 min, clarified by centrifugation (12,000–g for 3 min), and stored at −80°C until testing. Tissues (heart or kidney) and/or oropharyngeal swabs from all carcasses tested positive for WNV by either plaque assay (virus isolation) or reverse transcription polymerase chain reaction (RT-PCR), except for those of 9 birds that served as negative controls.
Feather collection and storage
Clusters of nonvascular feathers were carefully removed from the breast (i.e., ventral feather tract). The number of feathers collected per bird varied depending on the size of the bird as well as relative feather size/weight (Table 1). Mass of feathers tested per bird ranged from 0.001 g to 0.009 g.
For Colorado birds, half of the feathers collected from each carcass were held at room temperature (approximately 25°C), while the other half were held at freezer temperature (–20°C) for 1 year prior to testing. For California birds, immediately following collection, feathers were frozen at −80°C for several months until thawed for homogenization and testing.
Feather grinding techniques
Matched sets of feathers from bird carcasses collected in California (n = 68) were homogenized using 2 different techniques for each carcass. One technique was grinding between 1 and 5 whole feathers from a given carcass (Table 1) in a ground-glass tissue grinder c in 2 ml BA–1 diluent with inert alundum crystals added to facilitate homogenization (hereafter, “manual grinding”). The second technique consisted of grinding an equal number of whole feathers in a mixer mill b with 1 copper-coated stainless steel BB per vial as previously described 26 (hereafter, “mechanical grinding”). Colorado feathers were homogenized using only the former technique. Feather homogenates were clarified by centrifugation for 3 min at 16,000 x g, transferred to sterile cryovials, and stored at 4°C until testing by TaqMan RT-PCR and virus isolation within several days.
Inhibition of WNV detection in feathers
To evaluate the possibility of test inhibition as a result of components of the feather vane that may be liberated during grinding, we compared WNV RNA detection rates in matched pairs of feathers or feather components from each of 28 naturally infected American crow carcasses. First, matched pairs of breast feather calami (i.e., tips, cut from the rest of the feather at the proximal edge of the feather vane) were ground by either mechanical or manual grinding. Mechanically ground calami was then compared with mechanically ground whole feather, again in matched pairs, as well as manually ground calami with manually ground whole feather. In all cases, homogenates were clarified by centrifugation and tested by TaqMan RT-PCR.
Reproducibility of WNV detection in feathers
To assess the reproducibility of WNV RNA detection in feathers and determine the sensitivity of single versus multiple feathers tested from a single carcass, sets of 10 whole breast feathers were mechanically ground from each of 33 carcasses of naturally infected birds. Each feather was individually ground and tested by TaqMan RT-PCR.
Distribution of WNV in feathers
To determine the distribution of WNV RNA and infectious virions within the feather, the calamus and vane (including the rachis) were tested separately for each of 6 individual feathers removed immediately following euthanasia of experimentally inoculated crows (3 fish crows [Corvus ossifragus] and 3 American crows [C. brachyrhynchos]) at 6 days postinoculation (experimental details of these inoculations will be reported in a separate publication). Calami and vanes were each mechanically ground and clarified by centrifugation as described above, and supernatants were tested by TaqMan RT-PCR and virus isolation.
Plaque assay
For Colorado samples, virus isolation was performed on heart tissue and oropharyngeal swabs by Vero cell plaque assay. 29 In addition, plaque assays were performed on homogenates of whole feathers and feather components. All viral plaques were harvested and identified by a commercial WNV antigen detection assay. d
Reverse transcription polymerase chain reaction
TaqMan RT-PCR methods for detection of WNV RNA were performed on oropharyngeal swabs, hearts, kidneys, and feathers. RT-PCR methods followed those of Lanciotti et al. 21 except for the use of viral RNA mini kits e for RNA extraction and a real-time detection system f for complementary DNA (cDNA) amplification. A critical threshold value of <38.0 cycles obtained for 2 distinct sets of WNV-specific primers was considered positive for target sequence amplification. Positive RNA extraction and RT-PCR controls contained viral titers of approximately 106.5 and 104.5 plaque forming units (PFU) of WNV strain NY99–4132; tap water was used as a negative control for RNA extraction and a negative control for RT-PCR contained all reaction components except for RNA template.
Statistical analyses and determination of sensitivity and test agreement
Sensitivity and k values of WNV RNA detection testing of whole nonvascular feathers were derived by comparison of feather TaqMan RT-PCR with oropharyngeal swab, heart, or kidney TaqMan RT-PCR (referred to hereafter as “standard diagnostic methods”). Sensitivity was defined as the proportion of carcasses that were positive by standard diagnostic methods and also yielded WNV-positive feathers. Test agreement was measured using the kappa statistic, k, defined as the proportion of positive and negative carcasses determined by standard diagnostic methods that matched the feather test result. The feather test sensitivity and k for each species group were compared with the corresponding values of all other species combined using a two-tailed Fisher's exact test and Bonferroni-adjusted α = 0.0125 (for 4 comparisons representing each species group). Two-tailed Fisher's exact test was also used to compare WNV detection rates between grinding methods (i.e., manual vs. mechanical) and feather components (i.e., whole feather vs. calami). Mean viral load measures were compared with a T-test of unequal sample size and unequal variance, following log transformation of viral titers.
Results
Sensitivity of WNV detection in nonvascular feathers
Feathers were collected from 109 carcasses, with 100 WNV-positive and 9 negative. Feathers from 50 individuals of 17 species tested positive. Among feathers stored frozen (–l20°C or −80°C), sensitivity of detection by RT-PCR was greatest for corvid passeriforms (64%), followed by noncorvid passeriforms (43%), columbiforms (33%), and falconiforms (31%; Table 1). Among these 4 groups, only corvid feathers were significantly more sensitive for WNV detection than other species groups (Fisher's exact test, P = 0.001).
Storage temperature
The effect of storage temperature on WNV detectability was tested by examining sets of matched feathers held at ambient room temperature and −20°C from a subset of the WNV-positive carcasses (n = 32). WNV RNA was detected in 47% of feathers stored at room temperature and in feathers stored at −20°C, with k = 0.94. Infectious virions were detected in 16% of the feather sets stored at −20°C, with a range of viral titers from 101.0–3.6 PFU/ml of feather homogenate, and 0% of corresponding feather sets stored at ambient room temperature during the same period.
Feather grinding techniques
Manual grinding was compared with mechanical grinding for matched sets of whole feathers derived from individual carcasses using TaqMan RT-PCR and virus isolation (n = 68). In addition, these grinding methods were compared for calami from American crows (n = 28) using TaqMan RT-PCR. Mechanical grinding of whole feathers led to significantly higher detection rates (56%) than manual grinding (35%) using TaqMan RT-PCR but not virus isolation (Fisher's Exact test, P = 0.02 and 0.83, respectively), with k = 0.74 and 0.91, respectively. However, among feathers positive for infectious virions, mean viral loads were greater in mechanically ground feathers (104.6 PFU/ml feather homogenate, maximum 105.7 PFU) than manually ground feathers (103.0 PFU/ml feather homogenate, maximum 103.9 PFU; T-test, P = 0.09).
Inhibition of WNV detection in feathers
Because manual grinding provided better homogenization of the feathers, it was suspected that this grinding method liberated inhibitors of WNV detection and evaluated this hypothesis with 3 comparisons assessed from feathers harvested from 28 WNV-positive American crow carcasses: 1) WNV RNA detection probability in matched pairs of breast feather calami (1 pair per carcass) with one feather ground mechanically and the other manually; 2) WNV RNA detection probability in calamus versus whole feather, ground mechanically, and 3) WNV RNA detection probability in calamus versus whole feather, ground
Number of West Nile virus (WNV)-positive feathers by reverse transcription polymerase chain reaction among 33 individuals, each with 10 breast feathers individually tested. *

N = number of birds. Detection probability represents the probability that WNV will be detected in a single breast feather from a WNV-positive carcass. Numbers in parentheses are 95% confidence interval.
manually. For the first comparison, manual grinding produced a higher detection rate (32%) among calami than mechanical grinding (18%), although this difference was not significant (Fisher's Exact Test, P = 0.36). For the second comparison, 61% of mechanically ground whole feathers were positive, versus 18% of mechanically ground calami (P = 0.002). For the third comparison, manually ground calami tested positive at 32%, whereas the corresponding whole feathers tested positive at 29% (P = 1.0).
Reproducibility of WNV detection in feathers
Evaluating the reproducibility of WNV RNA detection among sets of 10 nonvascular breast feathers indicated substantial variation among carcasses, ranging from 0 to 10 feathers testing positive per WNV-positive carcass. At the species level, however, no significant differences were discerned, with the mean number of feathers testing positive (in species with minimum n = 2) ranging from 2.2 to 6.5 (Table 2). The overall probability of detection per feather among 33 carcasses tested was 0.32.
Distribution of WNV in feathers
WNV RNA was detected more frequently in the vane than the calamus in experimentally inoculated crows, whereas infectious virus was detected with greater frequency and in higher concentrations in the calamus (Table 3).
Results of West Nile virus (WNV) testing of feather calamus versus vane of experimentally inoculated crows at 6 days postinoculation. *
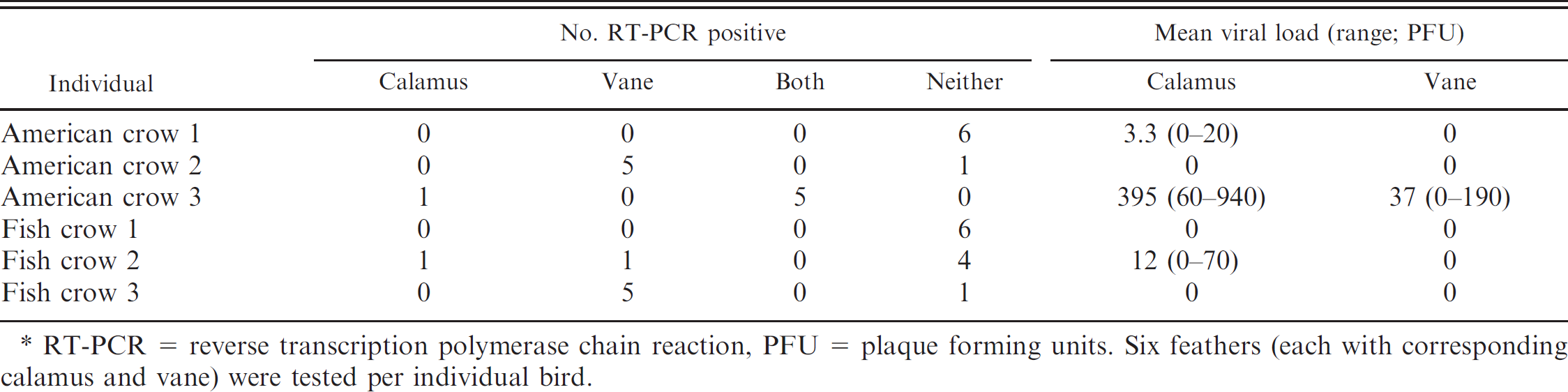
RT-PCR = reverse transcription polymerase chain reaction, PFU = plaque forming units. Six feathers (each with corresponding calamus and vane) were tested per individual bird.
Discussion
Feather testing has proven useful for diagnosis of numerous avian pathogens, and feathers may be a source of transmission. Feather follicle epithelium is a source of infectious Marek's disease virus and is important in natural transmission, 2 and feather calami are used for PCR detection of viral DNA in free-ranging birds. 23 Surveillance for Beak and feather disease virus and Avian polyomavirus also utilizes RT-PCR and PCR testing of feathers, respectively. 1,32,38 Relatively high titers of Psittacid herpesvirus 1 were found in pulp and quill of nonvascular feathers of parrots during an outbreak of Pacheco's parrot disease; the authors suggested that feather pulp may be useful for diagnosis. 10 Finally, highly pathogenic Avian influenza virus (HPAIV) H5N1 virions were visible in the feather epidermis of experimentally inoculated domestic ducks and geese 37 and transmission of HPAIV H5N1 occurred via oral ingestion of down feathers in ducks. 36
Feathers have been examined previously for WNV diagnosis, but within relatively limited parameters. One study compared WNV testing of vascular pulp from growing feather calami with cloacal swabs, kidney, and spleen tissues from carcasses of American crows and blue jays (Cyanocitta cristata). Of 65 WNV-positive birds, WNV was isolated from 100% (65/65) of feather pulp samples (from tail and wing feathers), 55% (36/65) of kidney and spleen pool samples, and 49% (32/65) of cloacal swabs. 5 The authors concluded that feather pulp was useful for WNV diagnosis; however, they recognized that duration and timing of molt varies by avian species and limits the utility of growing (i.e., vascular) feathers. A second study compared ventral tract feathers (a category that includes breast feathers) with cloacal swabs for detection of WNV in crow carcasses using antigen-capture enzyme-linked immunosorbent assay. Specificity and sensitivity for WNV detection in feathers were 96% and 71%, respectively. 9 These studies excluded noncorvid birds, which also have utility in WNV surveillance. 24,25 Another study examined nonvascular whole feathers from a small group of eastern screech-owls at specific time points following experimental WNV infection. 26 Feathers from 3 of 5 needle-inoculated owls tested positive for WNV RNA at 7 days postinoculation. Two of these owls succumbed to infection several days later, at which time feathers were still positive, while the third owl's feathers tested negative at 14 days postinoculation. Finally, WNV was detected by RT-PCR in whole feathers collected 21 days apart from a naturally infected, symptomatic red-tailed hawk (Buteo jamaicensis), while feathers tested positive between 10 to 22 days following onset of clinical signs in several other raptors, all of which survived infection in a rehabilitation clinic and were eventually released back into the wild (unpublished data). These findings raise 2 issues. First, WNV-positive feathers in carcasses may derive from prior infection unrelated to cause of death, and, second, feathers may be useful in WNV diagnosis in live birds.
In the present study, sensitivity of WNV detection in nonvascular feathers was lower than those reported in previous studies of corvid carcasses. 5,9 Various factors may explain this reduced sensitivity, including avian species differences, feather type, feather development stage (e.g., whether pulp or blood is present in the shaft), quantity of feathers tested, feather storage conditions, and components included for testing (e.g., feather calami vs. whole feathers). The developmental stage of the feather is presumed to be the key factor. However, species selection had a clear effect in the current study. Feathers from corvids were significantly more sensitive for WNV detection than feathers from noncorvid species. The effect of feather type or anatomical location of the feather was not tested in the current study; rather, feathers were exclusively from the ventral tract. However, viral tropism for certain feathers or feather clusters could exist. In addition, reproducibility of WNV detection in ventral tract feathers was approximately 30%. Therefore, sensitivity can be increased by pooling multiple feathers from a single carcass. No effect of storage condition on RNA detection was observed, while detection varied between calamus and vane depending on grinding method. This variation may be, in part, caused by detection inhibitors derived from certain feather components.
Potential inhibition of RT-PCR by certain feather components merits further study, as mitigation of such could improve the utility of feathers in diagnosis of WNV infections. For example, melanin from the shafts of brown chicken feathers was shown to inhibit PCR detection of Marek's disease virus and was mitigated by addition of bovine serum albumin to PCR reactions. 12 Data from the current study indicates a possible inhibitory effect from crow feather vanes because comparative tests among matched feathers demonstrated that 1) manual grinding yielded more positive calami than mechanical grinding, as expected because of the much more complete homogenization of tissue from manual grinding; 2) manual grinding yielded more positive calami than manual grinding of whole feather, possibly explained by release of inhibitors such as melanin from the vane; 3) mechanical grinding yielded more positive whole feathers than calami, explained by the inability of mechanical grinding to release inhibitors from the vane, and 4) mechanical grinding yielded more positive whole feathers than manual grinding, again explained by the release of inhibitors by complete homogenization of feather vane in the manual grinding process. Vane-derived inhibitors can be avoided either by testing calami only or by incomplete homogenization of vanes. Results indicate that twice as many positives were obtained by incomplete homogenization of whole feathers by the mechanical grinding protocol in the present study as compared with complete (manual) homogenization of calami.
The current study elucidates the potential use of feathers for WNV RNA detection in avian mortality surveillance programs. Although previous studies determined that vascular, growing feathers are the most sensitive tissue for WNV detection in corvids, it was demonstrated in the present study that nonvascular feathers are also useful, although with reduced sensitivity in corvids and relatively lower sensitivity in other species groups such as noncorvid passeriform birds, columbiforms, and falconiforms. Testing of nonvascular feathers would be most useful for surveillance in remote areas where maintaining the cold chain is unavailable (e.g., lack of dry ice or liquid nitrogen). To maximize the sensitivity of feather testing for WNV in American crows, the authors recommend testing multiple whole feathers per bird, as 6 feathers should yield approximately 90% sensitivity given a 32% detection probability per feather by RT-PCR. Because feather mass varies with body size and feather type or anatomical location, the optimal number of feathers for WNV detection may vary by species and feather tract. Potential inhibition from feather vane components should be mitigated as discussed above.
In summary, WNV particles, and more importantly RNA, persist in nonvascular feathers from a broad spectrum of bird species. Although nonvascular feathers may not be the most efficacious biological specimen for detecting WNV in avian carcasses, they offer numerous advantages to surveillance programs, including ease of collection, and simpler storage and shipping conditions. Additional potential advantages include longer period of detectability compared with tissues, application to live birds (morbidity surveillance and diagnosis in individual patients), and lower biosafety risk. Although infectious viral load in nonvascular feathers may be up to 105.7 PFU per feather, the virions are presumably sequestered within a keratinized sheath such that handlers are protected from the infectious fluids associated with most tissues, such as feather pulp and body cavities used for swabbing or removal of tissue for storage. However, to the authors’ knowledge, contamination of external feather surfaces has not been examined. Taken together, these advantages may warrant utilization of feathers for avian morbidity and mortality surveillance of WNV, HPAIV, and other important zoonotic pathogens.
Acknowledgements
The authors are grateful to the following for providing samples: Gail Kratz, Judy Scherpelz, Lisa Winta, Carin Avila, and Rebecca Bates of the Rocky Mountain Raptor Program, Bob Nightwalker and Jessica Plunkett of Wild-Kind (Larimer County Humane Society), and Jackie Parker of California Animal Health and Food Safety Laboratory. Richard Bowen (Colorado State University) and Susan Beckett (CDC) provided logistical support, and Maureen Dannen and Ying Fang (University of California, Davis) provided technical support. Funding for RNA detection platforms used for WNV RNA detection was partially provided by the Pacific Southwest Regional Center for Excellence (PSWRCE) U54 AI-65359.
Footnotes
a.
Nasco International Inc., Fort Atkinson, WI.
b.
Retsch GmbH, Haan, Germany.
c.
Tenbroek tissue grinder, Wheaton Science Products, Millville, NJ.
d.
VecTestr WNV/SLEV Antigen Assay, Medical Analysis Systems Inc., Camarillo, CA.
e.
Qiagen Inc., Valencia, CA.
f.
iCycler iQTM Real-time Detection System, Bio-Rad Laboratories, Hercules, CA.