
Abstract
The capacity of a commercially available gamma interferon (IFNγ) assay to detect infected sheep early in the pathogenesis of Johne's disease enables the removal of such animals from the flock before bacterial shedding and pasture contamination. However, nonspecific IFNγ responses in the assay have meant that to achieve high-test specificity, there has been a reduction in sensitivity. Although the optimal conditions for the use of the assay in cattle have been well documented, there have been few studies optimizing the assay for use in sheep. The current study details the effect of anticoagulant, duration of incubation, cell concentration, blood storage temperature, time of stimulation of cells with antigen relative to time of sample collection, and temperatures during transit on IFNγ synthesis. Maximal IFNγ synthesis occurred with incubation periods of 48 hr in blood collected into heparinized tubes. Decreasing the leukocyte population by diluting the total peripheral blood leukocyte concentration was associated with a decreasing IFNγ response. Conversely, concentrating the peripheral blood concentration 2-fold resulted in an increase in the IFNγ production. In field studies, immediate incubation of blood samples with antigen at 37°C resulted in larger IFNγ responses; however, significantly lower IFNγ values were obtained if the samples were transported at ambient temperature. The results of this study indicate that optimization of the IFNγ assay may enable increased synthesis of IFNγ during the stimulation phase of the assay and that future work may determine whether this translates to increased sensitivity of the assay in detecting early infections in sheep.
Introduction
The Bovigam sandwich assay a is a commercially available, biphasic, whole blood assay using an enzyme-linked immunosorbent assay (ELISA) to measure gamma interferon (IFNγ) production. The assay was originally developed for use in cattle in the diagnosis of tuberculosis, 1 and, with a modification to the antigen used in the whole blood stimulation phase of the assay, has subsequently been used in the diagnosis of bovine Johne's disease. 2 The assay detects the cell-mediated immune responses that, in diseases such as Johne's disease, typically occur earlier than the humoral responses that are often associated with onset of clinical disease. The optimal conditions for use of the Bovigam sandwich assay in cattle have been well documented. 3 It is recommended that bovine blood samples be collected in blood tubes in which heparin salts are used as the anticoagulant and that 16 hr is the optimum incubation time for the whole blood stimulation phase of the assay. 3 It is also recommended that the blood is stimulated within 8 hr of blood collection to obtain maximum IFNγ synthesis. 3,4 This is a situation that may be difficult to achieve, particularly with samples collected from farm animals in remote field settings.
Ovine IFNγ is also detected by the Bovigam sandwich assay 1,5 and, in recent years, the assay has been evaluated as a diagnostic test for use in ovine Johne's disease (OJD) control programs. 6,7 The assay's capacity to detect infected animals early in the disease process enables identification of potentially infected sheep before any bacterial fecal shedding. Irrespective of whether these animals progress to the clinical stage, removal of such animals from the flock has the potential to decrease contamination of pasture and, subsequently, to reduce infection of other sheep. However, because of nonspecific IFNγ responses, raised cutoff points have been required to achieve high specificity (≥98%), 7 which has resulted in a reduction of sensitivity to less than 50%, limiting its application for early detection of infection or certification of disease freedom. Optimization of the IFNγ production in ovine blood samples may improve the diagnostic sensitivity of the assay. The current study, therefore, aimed to improve the utility of the Bovigam IFNγ assay a in sheep by investigating the effect of anticoagulant, duration of incubation, cell concentration, blood storage temperature, time of stimulation of cells with antigen relative to time of sample collection, and temperature during transit on IFNγ production in whole ovine blood.
Materials and methods
Sheep
All experiments using housed or field animals in the present study were carried out with the approval of the University of Sydney Animal Ethics Committee. For the preliminary investigations, 2 Merino sheep approximately 7 months old were used. Only 2 animals were used in the study to maximize the number of incubation condition variables studied. The sheep had been vaccinated with an adjuvanted, killed Mycobacterium avium subspecies paratuberculosis (MAP) vaccine b at least 14 days before the experiment according to the manufacturer's instructions. The animals were fed lucerne chaff and grain once daily with water supplied ad libitum. For the experiments involving naturally infected sheep, Merinos, 3 years of age or older, were obtained from a farm with endemic OJD in the Central Tablelands of New South Wales, Australia, where vaccination is not currently used against the disease. The animals were screened for OJD by agar gel immuno-diffusion (AGID) test. Sixteen AGID-positive sheep were transported to a containment facility at the University of Sydney, Camden Campus, where the animals were housed and fed lucerne chaff and grain once daily with water supplied ad libitum. For the large-scale field trial, 50 Merino wethers, aged approximately 7 months, were selected from a property in Trunkey (near Bathurst) in New South Wales, Australia. These sheep had been vaccinated b according to the manufacturer's instructions 5 months before blood collection. The choice as to whether vaccinated or naturally infected sheep were used for each of the experiments depended upon availability at the time of the experiment.
MAP antigen and blood collection
Mechanically disrupted MAP antigen was derived from strain 316V. c Blood was collected from the jugular vein into commercially available 10-ml blood collection tubes, d containing 144 United States Pharmacopeia (USP) units of spray-dried lithium–heparin for all experiments, except for those in which the effect of other anticoagulants was analyzed. In these experiments, blood was collected into commercially available, 10-ml, blood collection tubes d containing either potassium oxalate/sodium fluoride (100/sp, 1,000/ca) or potassium ethylenediamine tetra-acetic acid (EDTA; 15% solution, 0.117 ml, 17.55 mg [100/sp, 1,000/ca]).
Whole blood culture
Previous studies in cattle described the nonspecific release of IFNγ in some samples when whole blood was diluted in tissue culture media.
3
This complication was not observed with ovine blood samples in the current study, so blood was diluted 1:2 in media to facilitate adequate mixing with antigen or mitogen. The culture medium used for all experiments consisted of Roswell Park Memorial Institute (RPMI)-1640 medium,
e
containing 5% fetal bovine serum (FBS),
f
Gamma interferon enzyme-linked immunosorbent assay
The whole blood culture method was followed by the Bovigam sandwich ELISA as previously described 1 and performed according to the manufacturer's instructions. All samples were assayed in duplicate or triplicate, and optical densities (ODs) were measured on an ELISA plate reader h at 450 nm. Samples were reassayed if duplicate or triplicate values differed from their means by more than 10%.
Effect of concentration of peripheral blood leukocytes and total and differential peripheral blood leukocyte counts on IFNγ production
Concentration of peripheral blood leukocytes. For the experiments investigating the effect of concentrating leukocytes (i.e., doubling the total leukocyte count in whole blood) on the production of IFNγ, 10 ml of blood was collected into each of 2 commercially available blood collection tubes d from 4 naturally infected Merino sheep. One tube was spun in a centrifuge at 1,455 × g for 15 min. The buffy coat was then aspirated from this tube, added to the second blood tube, and mixed thoroughly on a blood mixer for 10 min before commencing the assay as described above.
Total and differential leukocyte counts. Total and differential white blood cell counts were performed manually on blood collected from 16 naturally infected Merino sheep. Total white blood cell counts were performed on cells suspended in Türk solution using standardized methodology, whereas differential cell counts were performed on blood smears stained with a commercially available Romanowsky stain i according to the manufacturer's instructions.
Field trial investigations into the effect of varying transport temperature and time of adding antigen on IFNγ production
Blood collection, transport conditions, whole blood assay, and ELISA. Experiments were conducted on blood obtained from 50 vaccinated Merino sheep aged approximately 7 months and selected from a property in Trunkey (near Bathurst) in New South Wales, Australia. Blood was initially collected into commercially available 10-ml blood tubes d containing lithium heparin anticoagulant, and then mixed and immediately dispensed in 1-ml aliquots into 1 of 9 polypropylene-capped tubes. g Three of the tubes were pretreated with 1 ml of media alone, 3 tubes were pretreated with 1 ml of media containing MAP antigen, and 3 tubes were pretreated with 1 ml of media containing PWM. The tubes were then stored at 4°C overnight and transported to the property at ambient temperature. The triplicate tubes were then treated in 1 of 3 ways:
Group 1.—For each sheep, 3 polypropylene-capped tubes containing 1 ml of blood in 1 ml of media alone, 1 ml of media plus antigen, and 1 ml of media plus mitogen, respectively, were transported to the laboratory at ambient temperature, which was estimated to vary between 10°C and 20°C. Once at the laboratory, the blood was placed on a laboratory bench overnight at approximately 20°C, then incubated at 37°C for 48 hr before harvesting the supernatant.
Group 2.—For each sheep, 3 polypropylene-capped tubes containing 1 ml of blood in 1 ml of media alone, 1 ml of media plus antigen, and 1 ml of media plus mitogen were transported to the laboratory in a portable incubator j set at 37°C. Once at the laboratory, the blood was incubated at 37°C overnight and for a further 48 hr before harvesting the supernatant.
Group 3.—For each sheep, 3 polypropylene-capped tubes containing 1 ml of blood in 1 ml of media were transported to the laboratory at ambient temperature, which was estimated to vary between 10°C and 20°C. Once at the laboratory, the blood was placed on a laboratory bench overnight at approximately 20°C. The following morning, either MAP antigen or PWM was added to 2 of the 3 tubes, whereas the third tube remained as a control tube without additives. All tubes were then placed in the laboratory incubator at 37°C for 48 hr before harvesting the supernatant. Once harvested, all supernatants from all 3 groups were stored at −20°C before being assayed in the Bovigam sandwich ELISA according to manufacturer's instructions.
Statistical analyses
All statistical analyses were performed using GenStat version 10, k Minitab version 14, l or SAS release 9.1. m
Effect of concentration of peripheral blood leukocytes on IFN γ production. Effect of concentration of peripheral blood leukocytes on IFNγ response in blood samples obtained from 4 naturally infected Merino sheep was evaluated using a paired t-test.
Effect of total and differential peripheral blood leukocyte counts on IFN γ production. Linear regression analyses were performed to evaluate the effect of total and differential leukocyte counts on IFNγ production, measured by OD values of samples containing MAP antigen and by adjusted OD values (OD value MAP – OD value media). These outcome variables based on OD values were regressed on explanatory variables representing the blood cell counts or percentages, namely white blood cell count, lymphocyte percentage, total lymphocytes, neutrophil percentage, total neutrophils, monocyte percentage, total monocytes, eosinophil percentage, and total eosinophils. All OD values were logarithmically transformed before analyses to normalize their distributions.
In addition, logistic regression analyses were performed to evaluate the effect of blood cell counts or percentages on probability of classification of a sample as positive. Samples were designated as positive or negative using 2 stringency criteria. For the more stringent level, the sample was considered positive if the absorbance of the stimulated sample (either PWM or MAP antigen) was 0.100 absor-bance units greater than the absorbance achieved for the nonstimulated control well (media only) for that animal and if the absorbance of the stimulated sample was twice that of the unstimulated sample (i.e., OD MAP/OD media only ≥2 and OD MAP ≥ OD media only +0.1). This classification criterion was reported by researchers using the Bovigam sandwich assay for detection of IFNγ in cattle. 8 For the less-stringent criterion, the sample was considered positive if the absorbance of the stimulated sample (either PWM or MAP antigen) was at least 0.05 absorbance units greater than the absorbance achieved for the nonstimulated control well (media only) for that animal and if the absorbance of the stimulated sample was at least 1.3 times that of the unstimulated sample (i.e., OD MAP/OD media only ≥1.3 and OD MAP ≥ OD media only +0.05). This criterion has also been reported for the detection of IFNγ in sheep. 9,10
In initial linear and logistic regression analyses, all explanatory variables were tested individually to determine their unconditional association with the outcome. Subsequently, explanatory variables associated with the outcome at univariable P-value of ≤0.25 were selected for inclusion in multivariable logistic regression models. Multivariable models were fitted using the manual, forward, stepwise approach employing the SAS LOGISTIC procedure, retaining variables with P-value < 0.05.
Effect of varying transport temperature and time of adding antigen on IFN γ production. Optical density values for samples containing media alone, MAP antigen, and PWM were compared between 3 treatment groups using analysis of variance. In addition, adjusted OD values, calculated by subtracting OD values of the samples containing media alone from the samples containing MAP antigen, were compared between 3 treatments. All OD values were logarithmically transformed before analysis of group differences to normalize their distributions. In a second set of analyses, OD values were categorized into positive and negative, and probability of classification of samples as positive was compared between 3 treatment groups using logistic regression analyses.
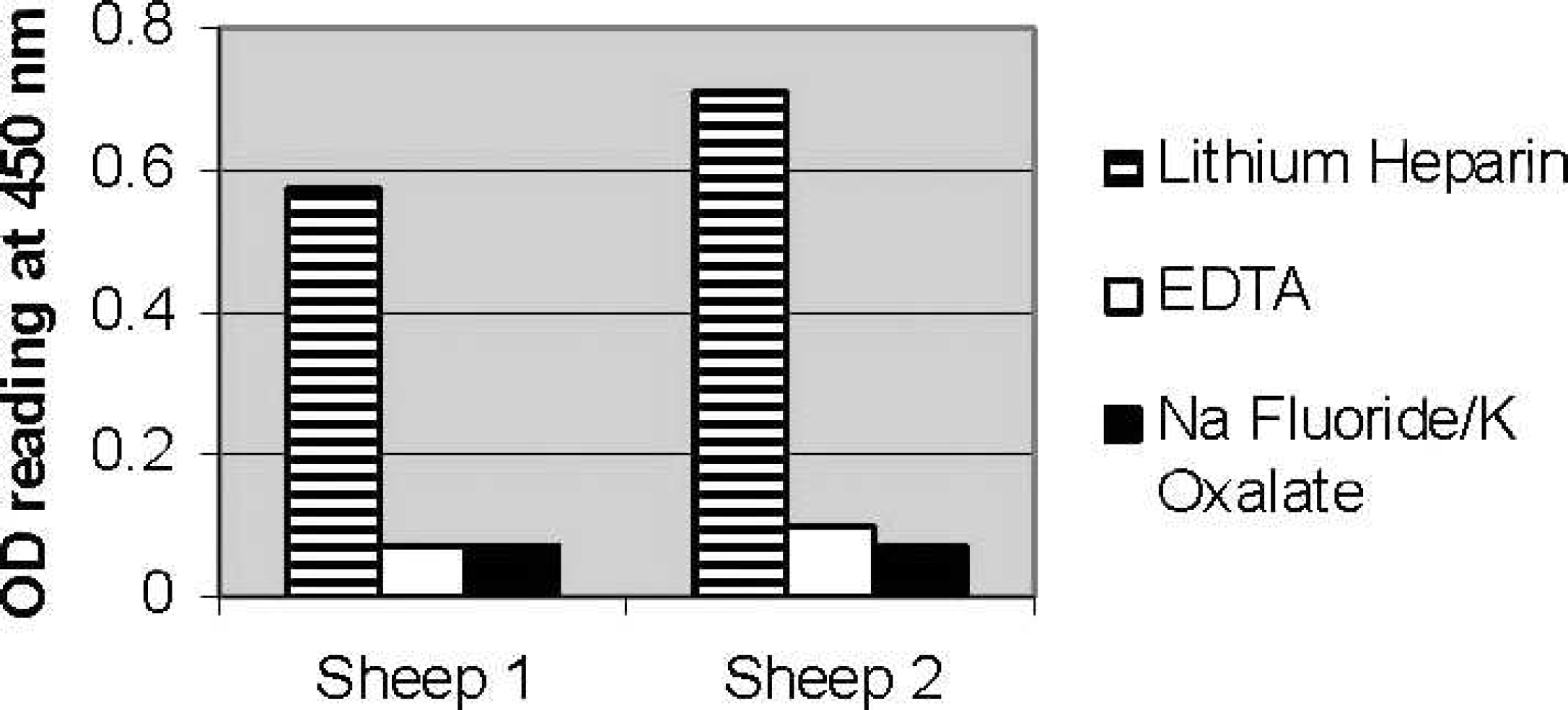
Effect of anticoagulant on gamma interferon production during incubation of whole blood with Mycobacterium avium subspecies paratuberculosis antigen (at a final concentration of 5 μg/ml) and measured as optical density (OD) at 450 nm in the Bovigam sandwich assay in 2 vaccinated Merino sheep. EDTA = ethylenediamine tetra-acetic acid.
Results
Effect of anticoagulant
The effect of anticoagulant on IFNγ production was evaluated in 2 vaccinated animals. Blood collected into tubes containing lithium heparin resulted in the optimal response with collection of blood into tubes containing potassium EDTA, and potassium oxalate/sodium fluoride, resulting in a complete abrogation of the IFNγ response (Fig. 1).
Effect of varying duration of incubation
The effect of varying duration of incubation on IFNγ production was evaluated in 2 vaccinated Merino sheep following stimulation in the whole blood assay with MAP antigen (final concentration of 5 μg/ml) and Con A mitogen (final concentration of 10 μg/ml). Blood was incubated for 24, 48, or 72 hr before harvesting the supernatant. In both sheep and with both MAP antigen and mitogen, optimum IFNγ production was obtained following 48 hr incubation (Fig. 2). The responses to MAP antigen were greater than those to ConA in both sheep.
Effect of blood storage temperature and duration
To account for field sampling and time to assay, the effect on IFNγ production of varying the temperature at which blood was stored for 4, 8, or 24 hr before incubation in the whole blood assay was evaluated in 2 vaccinated animals. Adding the MAP antigen at the time of blood collection consistently resulted in lower IFNγ production than the levels obtained when the antigen was added just before incubation irrespective of the time or temperature at which the blood was held before incubation (Table 1). For both sheep, regardless of when antigen was added, there were clear indications that the longer the delay to incubation, the less IFNγ was produced.
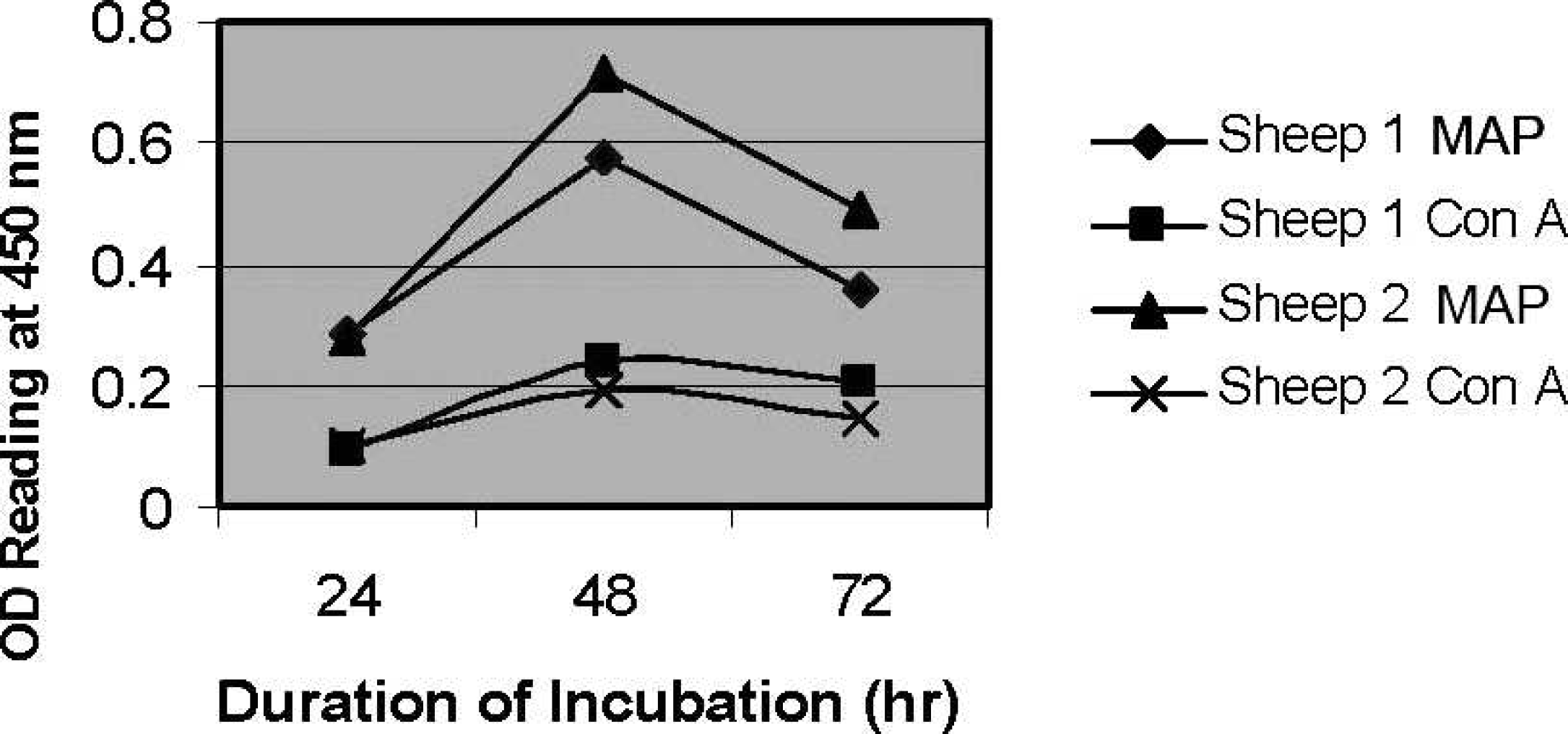
Effect of varying whole blood incubation time at 37°C on gamma interferon production in peripheral blood obtained from 2 vaccinated Merino sheep in response to stimulation with Mycobacterium avium subspecies paratuberculosis (MAP) antigen (final concentration of 5 μg/ml) and concanavalin A (Con A) mitogen (final concentration of 10 μg/ml) and measured as optical density (OD) at 450 nm in the Bovigam sandwich assay.
Effect of total white blood cell concentration and differential cell count
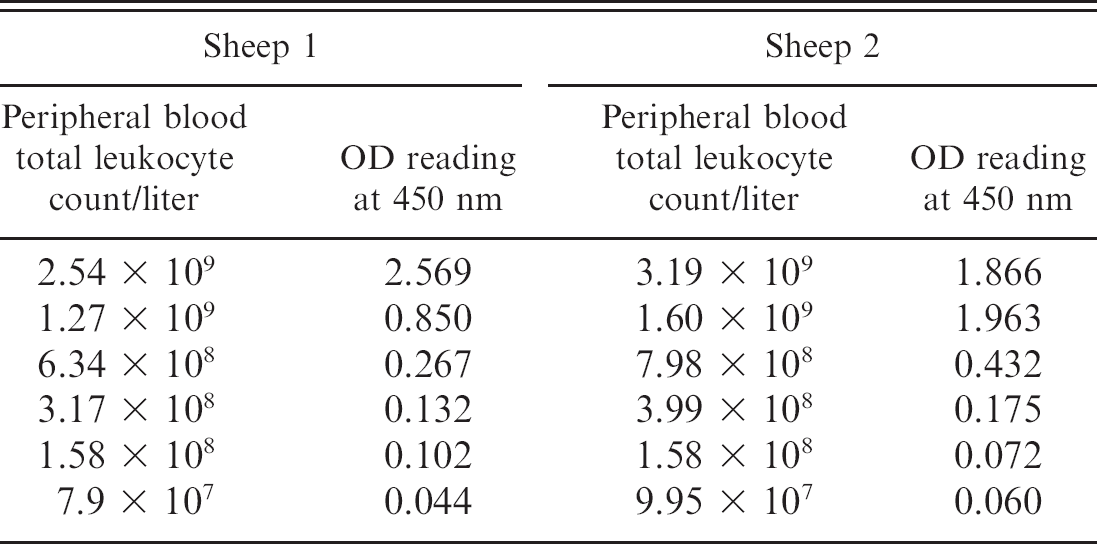
Effect of cell concentration. Decreasing cell concentration in vaccinated sheep.—Decreasing the responding leukocyte population by diluting the total peripheral blood leukocyte concentration in medium was associated with a decreasing IFNγ response (Table 2). There was a substantial reduction in IFNγ signal with 2–4-fold reductions in total white blood cell counts.
Increasing and decreasing cell concentration in naturally infected sheep.—The effect of both concentrating and titration of the peripheral blood leukocyte concentration with media was examined in 4 naturally infected Merino sheep. In all 4 sheep, concentrating the peripheral blood concentration 2-fold resulted in a significant increase in IFNγ production (P-value = 0.03). Subsequent serial doubling dilutions of the sample in media resulted in a dose-dependent decreasing IFNγ response (Fig. 3).
Effect of total and differential leukocyte counts on IFN γ production. These experiments were undertaken on blood obtained from naturally infected Merino sheep aged ≥3 years. Linear regression analyses were conducted to evaluate the effect of blood counts on IFNγ production, but none of the leukocyte variables was significantly associated with MAP OD values or MAP-adjusted OD values, indicating that they might not influence IFNγ production. Similarly, logistic regression analyses conducted for the 2 outcome variables created based on the stringent and the less-stringent criterion, respectively, suggested no effect of blood cell counts, except that there were higher odds of a positive outcome (stringent) in animals with high total lymphocyte counts.
Effect of delay in peripheral blood sample processing, and the temperature at which the sample was stored throughout that delay, on gamma interferon production in response to stimulation with Mycobacterium avium subspecies paratuberculosis antigen (at a final concentration of 5 μg/ml) measured as optical density at 450 nm (OD450nm) in the Bovigam sandwich assay in 2 vaccinated Merino sheep. *
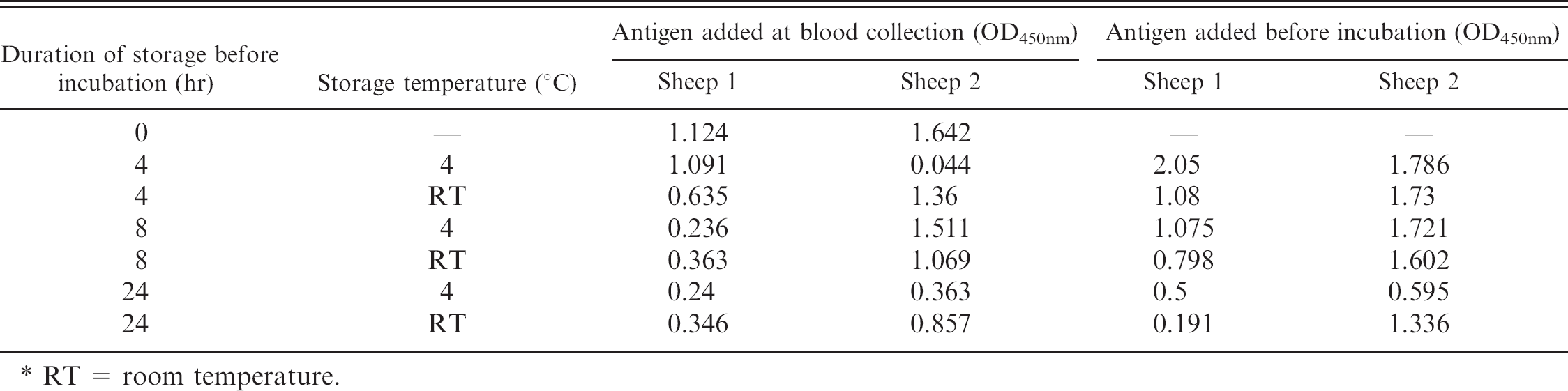
RT = room temperature.
Field trial investigations into the effect of varying transport temperature and time of adding antigen on IFNγ production
Effect of blood sample treatment on IFNγ response. The IFNγ production in 50 blood samples from vaccinated sheep to which MAP antigen was added at the time of blood collection before transport (3 hr at ambient temperature) to the laboratory was examined. The responses of this group (group 1) were significantly lower than for those samples in which the MAP antigen was added at blood collection with subsequent incubation at 37°C (group 2) or for those that were transported at ambient temperature to the laboratory with the antigen added 24 hr after blood collection (group 3). This effect was observed only with the samples stimulated with MAP antigen and not with those stimulated with PWM or media only, in which no significant differences were observed between the 3 groups (Fig. 4). A similar result was obtained when adjusted OD values were compared between 3 groups instead of raw OD values.
Effect of blood sample treatment on classification of a sample as IFNγ positive or negative. Two levels of stringency criteria were examined to determine whether adding antigen at blood collection and incubating the sample during transport would produce a positive IFN-γ result that would otherwise have been considered negative. Logistic regression analyses indicated no significant effect of treatment on the probability of classification of a sample as positive based on the more stringent criterion (criterion 1). However, based on the less-stringent criterion (criterion 2), the addition of antigen at the time of blood collection with subsequent transportation to the laboratory at ambient temperature (group 1) resulted in a significantly lower probability of a sample being positive (Fig. 5) compared with incubation at 37°C (group 2; P < 0.05). The difference between groups 1 and 3 (transportation to the laboratory at ambient temperature with the antigen addition 24 hr after blood collection) was not significant.
Effect of titration of white blood cell concentration on gamma interferon production in response to stimulation with Mycobacterium avium subspecies paratuber culosis antigen (at a final concentration of 5 μg/ml) measured as optical density (OD) at 450 nm in the Bovigam assay in peripheral blood samples obtained from 2 vaccinated Merino sheep.
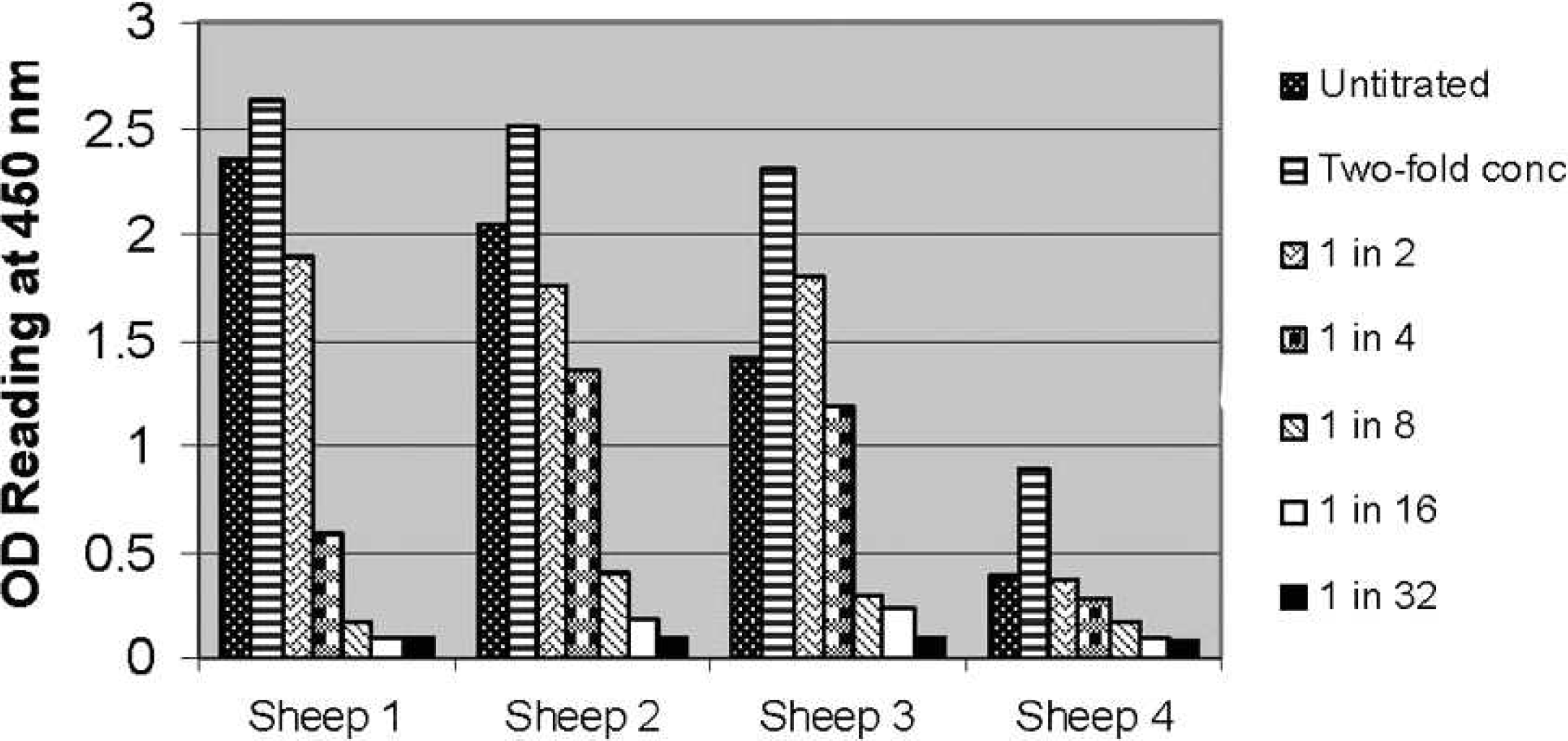
Effect of concentrating (2-fold) and diluting (down to 1 in 32) the leukocyte concentration on gamma interferon response measured as optical density (OD) at 450 nm in the Bovigam sandwich assay in peripheral blood samples obtained from 4 naturally infected Merino sheep.
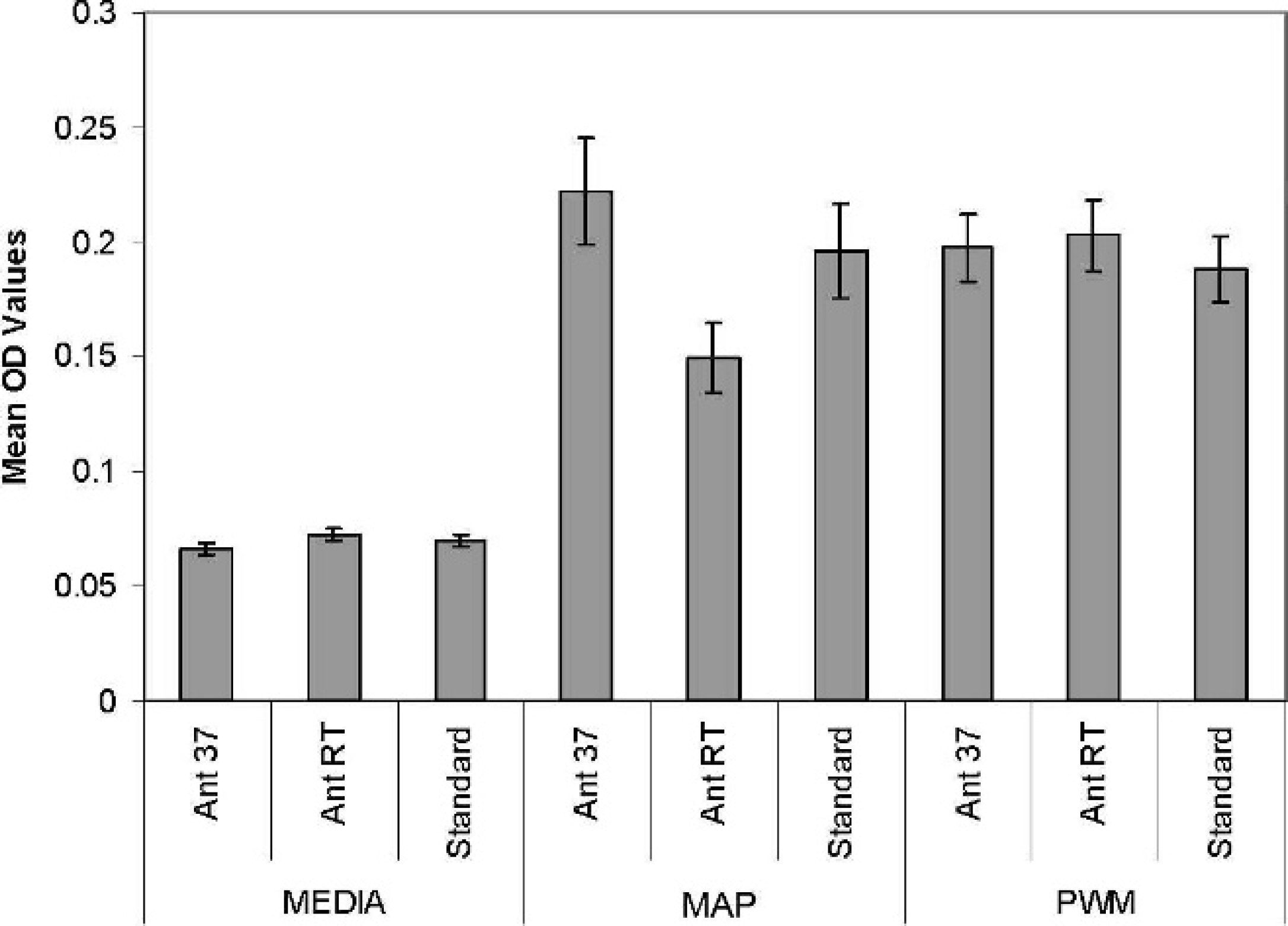
Effect of timing of addition of antigen and transport temperature on IFN-γ response measured as optical density (OD) at 450 nm in the Bovigam sandwich assay a in peripheral blood samples obtained from 50 vaccinated Merino sheep. Ant RT = Mycobacterium avium subspecies paratuberculosis (MAP) antigen added at the time of blood collection with subsequent transportation to the laboratory at ambient temperature; Ant 37 = MAP antigen added at blood collection with subsequent incubation at 37°C; standard = transportation at ambient temperature to the laboratory before MAP antigen added 24 hr after blood collection; PWM = pokeweed mitogen.
Discussion
The current study analyzes a suite of variables impinging on the reliable and reproducible assay of IFNγ from whole blood cultures in sheep. The finding that lithium heparin was the only anticoagulant enabling optimal IFNγ production with almost complete abrogation of the IFNγ response occurring with the use of sodium fluoride/potassium oxalate and potassium EDTA was consistent with results observed when the Bovigam sandwich assay was evaluated in cattle. 3,9 Similar results have been obtained in whole blood culture techniques for evaluating cytokine production 11 and lymphocyte reactivity 12 in humans.
In the present study, optimum IFNγ production was obtained after incubation of the ovine blood with the antigen or mitogen for at least 48 hr compared with overnight (16 hr) in cattle. 3 Interestingly, the finding that longer incubation times are required for optimum IFNγ production in sheep is consistent with that obtained in some studies with human blood. 11 In one study using a commercially available IFNγ assay n in humans, prolonged incubation for 6 days enhanced the sensitivity of the assay to detect latent Mycobacterium tuberculosis infection. 13
A dramatic decline in IFNγ production was observed associated with relatively minor reductions in the total white blood cell count. Cell density has been demonstrated to play a role in the reactivity of lymphocytes to stimulants in human studies 12 with marked decreases in lymphocyte responses demonstrated when cells are dispersed over larger surface areas, such as occurs if larger welled plates or flat bottom vessels are used for culture. This is presumably due to intercellular interactions that are hindered if the cells are not in close contact, which might occur with reductions in the total white cell count. Although the cell population predominantly responsible for production of IFNγ in sheep has not yet been clarified, the population in cattle has been identified as cluster of differentiation (CD)4 T lymphocytes. 14 In sheep, preliminary studies at the authors' laboratory suggest that this population comprises approximately 30% of the peripheral white blood cell count (D. Begg, unpublished data, 2009), and therefore, serial dilutions may result in a relatively low number of cells capable of producing IFNγ. In locations or circumstances where endemic parasitism could engender whole blood loss via the gastrointestinal tract, reductions in leukocyte number could potentially affect the sensitivity of the test. Similarly, sensitivity may also be affected by variations in leukocyte number because of physiologic and stress responses, such as the endogenous glucocorticoid-mediated lymphopenia that occurs in response to periods of chronic stress.
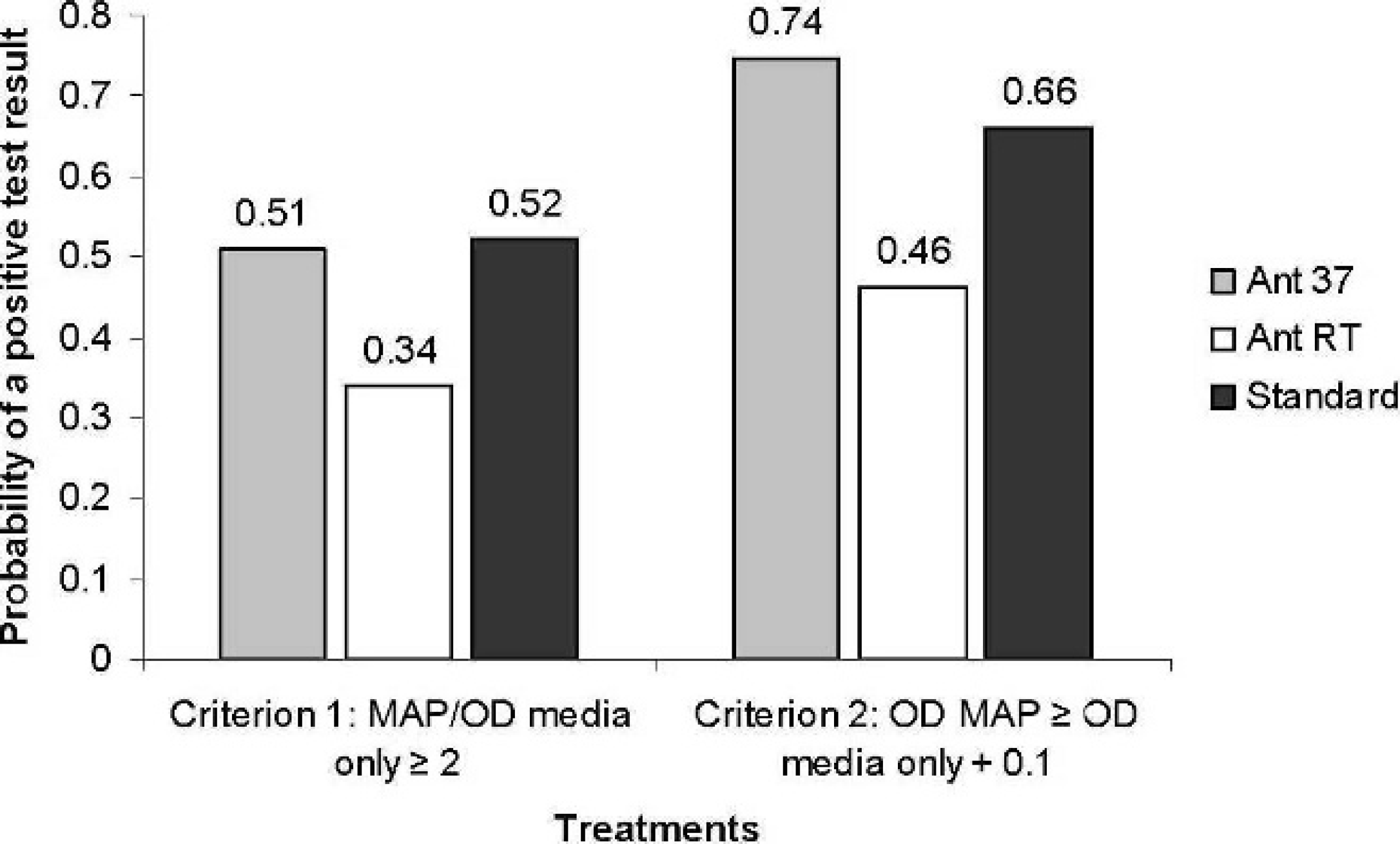
Logistic regression analysis to determine the effect of timing of addition of antigen and temperature during transport on the classification of the level of gamma interferon as positive or negative based on 2 classification criteria. Ant RT = Mycobacterium avium subspecies paratuberculosis (MAP) antigen added at the time of blood collection with subsequent transportation to the laboratory at ambient temperature; Ant 37 = MAP antigen added at blood collection with subsequent incubation at 37°C; standard = transportation at ambient temperature to the laboratory before MAP antigen added 24 hr after blood collection; OD = optical density.
Field studies with blood samples from children in remote areas of South Africa have demonstrated that immediate incubation with antigen at 37°C and throughout transportation to the laboratory resulted in optimal IFNγ production as measured by flow cytometry (Stewart DJ: 2005, MLA Harvest Year Conference Programme. In: Validation of the gamma-interferon test for diagnosis of ovine Johne's disease: Proceedings of the MLA OJD Harvest Year Conference, ed. Stewart DJ, Stiles PL, Horvatic B, et al., p. 4, December 8–9, 2005, Adelaide, Australia. Meat and Livestock Australia, North Sydney, New South Wales, Australia). However, the interaction of adding antigen at the time of blood collection with the temperature of transportation has not previously been described in production animals. A loss in IFNγ production associated with a delay in sample processing has been well documented. 3,4 ,16 Studies in cattle have demonstrated that cells transported to the laboratory at 37°C, without the addition of antigen during the transportation process, synthesized less IFNγ than those transported at room temperature. 9 More specifically, transport temperatures of 15.6–21.1°C were optimal by comparison with 4uC and values at or greater than 37.8°C, which resulted in reduced IFNγ synthesis.16 The results of the large field experiment in the present study suggested that immediate incubation of blood samples with antigen at 37°C could result in larger IFNγ responses in some samples. It was also demonstrated that if the MAP antigen is to be added to the blood at collection, it is imperative that the blood be transported at 37°C because significantly lower IFNγ values were obtained if the samples were transported at ambient temperature. This was not the case for those samples in which PWM was the additive, suggesting that the mechanism of mitogen activation was less temperature sensitive than antigen-induced reactions. A possible explanation for this finding could be that there is a component of the MAP antigen that negatively affects the cell's ability to remain viable or to respond to the antigen and that effect is abrogated at 37°C. The MAP antigen used in the present study is a crude bacterial preparation, and further studies should be directed toward determining whether this effect is repeatable with different sources of purified protein derivative (PPD) or methods of PPD production.
There have been other novel ways of potentiating IFNγ responses in production animals where the delay from sample collection on farm to assay in the laboratory is prolonged. One such study (Jungersen G, Grell SN, Clemensen A, et al.: 2005, Interleukin-12 potentiation of the interferon-gamma test rescues day-old blood samples for diagnosis of paratubercu-losis PPD specific cellular mediated immune responses. In: Proceedings of the 8th International Colloquium on Paratuberculosis, pp. 501–506, August 14–17, Copenhagen, Denmark. International Association for Paratuberculosis, Kennett Square, PA) reported that the addition of interleukin (IL)-12 to day-old samples collected from cattle and subsequent culture with MAP antigen produced IFNγ levels that were comparable to those from fresh, stimulated blood samples. Futures studies could also be directed at examining the interaction between storage temperature, timing of antigen addition, and the addition of potentiating cytokines, such as IL-12 or IL-10.
Acknowledgements
Research was funded by Meat and Livestock Australia (MLA). The authors would like to thank Ms. Natalie Schiller, Mrs. Anna Waldron, and Mr. Craig Kristo (Farm Animal Health Group, The University of Sydney); Mr. Jeff Eppleston (Rural Lands and Parks Board, Bathurst); and Mr. Hilton Brown (“Grovesdale,” Trunkey via Bathurst, New South Wales) for their excellent technical and animal assistance. Bovigam test kits were supplied by Pfizer Animal Health Australia with technical assistance provided by Ms. Eurife Abadin. Dr Stephen Jones (Cellestis International Pty Ltd.) kindly provided the portable incubator for the field trial.
Footnotes
a.
Bovigam®, CSL Ltd., Parkville, Victoria, Australia.
b.
Gudair®, Pfizer Animal Health, West Ryde, New South Wales, Australia.
c.
Provided by NSW Department of Primary Industries, Menangle, New South Wales, Australia.
d.
Vacutainer® tubes, BD, Franklin Lakes, NJ.
e.
RPMI medium 1640, Gibco-Invitrogen, Grand Island, NY.
f.
Sigma Chemicals Pty Ltd., Castle Hill, New South Wales, Australia.
g.
BD, Franklin Lakes, NJ.
h.
Multiscan Biochromatic 348, Helsinki, Finland.
i.
Diff-Quik, Lab Aids, New South Wales, Australia.
j.
Cellestis International Pty Ltd., Carnegie, Victoria, Australia.
k.
VSNI, Hemel Hempstead, Herts, United Kingdom.
l.
Minitab Inc., State College, PA.
m.
SAS Institute Inc., Cary, NC.
n.
QuantiFERON-TB®, Cellestis International Pty Ltd., Carnegie, Victoria, Australia.