
Abstract
By the end of 2004, the Canadian swine population had experienced a severe increase in the incidence of Porcine circovirus-associated disease (PCVAD), a problem that was associated with the emergence of a new Porcine circovirus-2 genotype (PCV-2b), previously unrecovered in North America. Thus, it became important to develop a diagnostic tool that could differentiate between the old and new circulating genotypes (PCV-2a and PCV-2b, respectively). Consequently, a multiplex real-time quantitative polymerase chain reaction (mrtqPCR) assay that could sensitively and specifically identify and differentiate PCV-2 genotypes was developed. A retrospective epidemiologic survey that used the mrtqPCR assay was performed to determine if cofactors could affect the risk of PCVAD. From 121 PCV-2-positive cases gathered for this study, 4.13%, 92.56%, and 3.31% were positive for PCV-2a, PCV-2b, and both genotypes, respectively. In a data analysis using univariate logistic regressions, the PCVAD-compatible (PCVAD/c) score was significantly associated with the presence of Porcine reproductive and respiratory syndrome virus (PRRSV), PRRSV viral load, PCV-2 viral load, and PCV-2 immunohistochemistry (IHC) results. Polytomous logistic regression analysis revealed that PCVAD/c score was affected by PCV-2 viral load (P = 0.0161) and IHC (P = 0.0128), but not by the PRRSV variables (P > 0.9), which suggests that mrtqPCR in tissue is a reliable alternative to IHC. Logistic regression analyses revealed that PCV-2 increased the odds ratio of isolating 2 major swine pathogens of the respiratory tract, Actinobacillus pleuropneumoniae and Streptococcus suis serotypes 1/2, 1, 2, 3, 4, and 7, which are serotypes commonly associated with clinical diseases.
Introduction
Postweaning multisystemic wasting syndrome (PMWS) is a disease of swine initially identified in Canada in 1991. 13 Now, it is known as a worldwide disease, and outbreaks have been reported in swine herds of North and South America, Europe, and Asia. 7 The disease affects 5- to 12-week-old piglets and is characterized in part by weight loss, dyspnea, jaundice, and enlarged lymph nodes, as well as by degeneration and necrosis of hepatocytes, multifocal lymphohistiocytic pneumonia, lymphocytic depletion, and multinu-cleated giant cell formation. 26 The etiologic agent responsible for PMWS has been identified as a circovirus particle named Porcine circovirus-2 (PCV-2; family Circoviridae, genus Circovirus). 1,19,25 The PCV-2 is a small nonenveloped virus that possesses a single-stranded ambisense circular DNA genome of about 1.76 kb in length. 20,33,41,54 Viral DNA possesses at least 3 functional open reading frames (ORFs): ORF1 encodes the Rep proteins involved in virus replication, 9,10,38 ORF2 encodes the nucleocapsid (Cap) protein, 42 and ORF3 encodes a protein that induces apoptosis and is involved in viral pathogenesis in vivo. 35,36 Today, the clinical expression of PCV-2 infection in swine is acknowledged to be more complex than initially established because it can play a pivotal role in several syndromes, such as porcine dermatitis and nephropathy syndrome, porcine respiratory disease complex, reproductive failure, granulomatous enteritis, necrotizing lymphadenitis, exudative epider-mitis, and congenital tremor. 8,27 Consequently, the term “Porcine circovirus-associated disease” (PCVAD) is now accepted to describe the syndromes in which PCV-2 plays a role.
At the end of 2004, the swine industry in the province of Quebec in Canada started to experience a significant increase in death rate related to PCVAD. 14 It was hypothesized that this emerging problem was caused by the presence of a new type of circulating PCV-2 strain. This was confirmed by sequencing the entire genome of several PCV-2 strains originating from PMWS-affected herds. 23 Interestingly, this newly recovered PCV-2 genotype (named PCV-2b in comparison with the circulating genotype named PCV-2a) had already been reported in Asia and Europe 15 and had also been recently reported in the United States. 11,30
The current international consensus is that at least 2 major PCV-2 genotypes are presently circulating worldwide. 11,17,23,24,43 It is noteworthy that different nomenclatures have been used to identify the PCV-2 genotypes: for instance, the PCV-2b and PCV-2a genotypes, 23 respectively, correspond to the restriction fragment length polymorphism (RFLP) classification 321 and 422 6 and to the genotypes 1 and 2. 24 Based on RFLP and gene-sequence comparison results, PCV-2b was suggested to be associated with the mortality rate increase observed in several Canadian swine herds. 6 Consequently, even if no experiment confirms yet if the PCV-2b strains found in Canada are more virulent than previously circulating PCV-2a strains, it became important to develop a low-cost (compared with entire viral genome sequencing) diagnostic test that could rapidly and efficiently differentiate both genotypes. Previous results indicated that the protein that best distinguishes PCV-2a from PCV-2b was the Cap protein, with an amino acids (aa) sequence identity varying from 88–94% between both genotypes. 23 Therefore, a new multiplex real-time quantitative polymerase chain reaction (mrtqPCR) assay was developed to detect with high sensitivity PCV-2 strains present in tissue samples of pigs. Furthermore, this new polymerase chain reaction (PCR) assay was able to identify PCV-2 genotypes by targeting the ORF2 gene. Afterward, a retrospective epidemiologic survey was conducted using PCV-2 mrtqPCR-positive cases that were submitted to the molecular diagnostic laboratory of the Faculté de médecine vétérinaire (FMV) of the University of Montreal to determine the prevalence of PCV-2 genotypes and to establish if other viral or bacterial pathogens were associated with the clinical disease. Furthermore, because PCV-2 antigen detection in microscopic lesions by immunohistochemistry (IHC) is one of the three criteria used for the diagnosis of PCVAD cases, 51 the survey was conducted to determine the relationship between the PCV-2 IHC results and the PCV-2 viral load with PCVAD.
Materials and methods
Organisms and culture methods
All bacteria and viruses used as controls were isolated and characterized at the FMV. The PCV-2 viruses, IAF 2897 (genotype PCV-2a) and FMV-06-1717 (genotype PCV-2b), used in the mrtqPCR assay were previously characterized reference strains. 23 Those viruses were propagated into PK15A cells, a subclone of PCV non-infected PK15 cells, 49 and the viruses were purified after a 4-hr period of ultracentrifugation on a 30% sucrose cushion using the SW28 Beckman Coulter rotor a at 83,000 X g. The virus pellets were resuspended in 2 ml of a phosphate buffered saline (PBS) containing 2% fetal bovine serum, and aliquots of the virus stocks were then conserved at −70°C for future use. The infectious dose of the virus stock was calculated from a 96-well microplate of PK15A-infected cells after fluorescent antibody test 49 by the Kärber method. 47 Virus titers were expressed in tissue culture infectious dose 50 per ml (TCID50/ml).
Porcine circovirus-2 recombinant plasmids
Extraction of PCV-2 IAF2897 and FMV-06-1717 isolates of DNA was performed using the Qiagen QIAamp DNA Mini Kit b according to the manufacturer's instructions. The PCR products encompassing the entire viral genome of both strains were produced using the following primers: Fw-SacII-PCV2 (GAACCGCGGGCTGGCTGAACTTTTGAAAGT) and Rv-SacII-PCV2 (GCACCGCGGAAATTTCTGACAAACGTTACA). The PCR products were subsequently cloned in the plasmidic vector pCR2.1 using the TA (Taq-amplified) Cloning Kit c according to the manufacturer's instructions. Finally, the recombinant plasmids (pCR2.1/PCV-2a and pCR2.1/PCV-2b) were sequenced to verify the accuracy of the nucleotide sequences and were used as DNA templates to validate the new mrtqPCR diagnostic assay.
DNA/RNA extraction
Extraction of DNA and RNA from tissue samples (lungs and/or lymph nodes) and virus isolates was performed using the Qiagen QIAamp DNA Mini Kit
b
and the Qiagen QIAamp Viral RNA Mini Kit,
Multiplex real-time quantitative polymerase chain reaction assay
Primers and probes were selected within 2 regions of the PCV-2 genome based on nucleotide alignments generated by the BioEdit Sequence Alignment Editor (version 7.0.5.2) e of available sequences in public domain databases (more than 76 sequences of PCV-2a and PCV-2b strains were obtained from GenBank and compared) and sequences obtained from viral specimens in the authors' molecular diagnostic laboratory (more than 49 sequences of PCV-2a and PCV-2b strains). 23 For general detection of PCV-2, a conserved region of PCV-2 genomes was targeted by the mrtqPCR using the following primers and probe: Circo-Gen-F (GGCCACCTGGGTGTGGTAAA), Circo-Gen-R (CCCACCACTTGTTTCTAGGTGGTT), and Circo-Gen-Probe (6-FAM-TTTGCAGACCCGGAAACCACATACTGGA-BHQ-1). For genotyping, a hypervariable region localized in the ORF2 of the PCV-2 was targeted by the mrtqPCR using the following primers and probes: Circo-DF (GGGCCAGAATTCAACCTTAA), Circo-DR (CGCACCTTCGGATATACTATCA), Circo-2a-Probe (Cy5-GGGGACCAACAAAATCTCTATACCCTTT-BHQ-2), and Circo-2b-Probe (Cy3-GGGCTCAAACCCCCGCTCTGTGCCCTTT-BHQ-2).
In the multiplex reaction, 5 μl of sample containing the DNA template was added to a 20-μl reaction mixture (4.8 μl of LightCycler FastStart DNA MasterPLUS HybProbes, f 400 nM of each primer and genotyping probes, and 120 nM of Circo-Gen-Probe). Thermocycling and detection were performed in a SmartCycler 6400 System. g The PCR reaction started with an initial denaturation and polymerase-activating step of 95°C for 10 min, followed by 45 amplification cycles of a 3-step PCR (95°C for 30 sec; 60°C for 60 sec; and 72°C for 60 sec).
To assess the sensitivity and specificity of the mrtqPCR assay and to determine its ability to differentiate the 2 PCV-2 genotypes, 10-fold serial dilutions of PCV-2a and PCV-2b reference viruses and PCV-2 recombinant plasmids (pCR2.1/PCV-2a and pCR2.1/PCV-2b) described previously were assayed. The viruses and recombinant plasmids were then quantified by mrtqPCR. Furthermore, validation of the mrtqPCR PCV-2 genotype classification from clinical submitted samples was confirmed after sequencing 49 PCV-2-positive cases (10 PCV-2a and 39 PCV-2b cases). A conventional multiplex PCR amplification (mPCR) for detecting type 1 and 2 circoviruses was also performed as previously described. 46 The specificities of the primer-probe sets were determined by testing several specimens from different animals, bacterial DNA, and nucleic acids of swine viruses. The samples consisted of PCV-2-negative porcine tissues, semen, and sera, as well as bovine tissues and various pathogens isolated by the FMV diagnostic laboratory, such as Brachyspira hyodysenteriae, Campylobacter spp., Clostridium perfringens, Enterococcus spp., Escherichia coli, Haemophilus parasuis, Lawsonia intracellularis, Mycoplasma hyopneumoniae, Pasteurella multocida, PCV-1, Porcine reproductive and respiratory syndrome virus (PRRSV), Salmonella spp., Staphylococcus hyicus, Streptococcus suis, Swine influenza viruses (SIVs) H1N1 and H3N2, and Transmissible gastroenteritis virus.
Retrospective epidemiologic survey
Sick or dead pigs that were submitted to and processed by the Institut National de Santé Animale (INSA), a subsidiary of the Ministère de l'Agriculture, des Pěcheries et de l'Alimentation du Québec (MAPAQ), were subsequently submitted to the diagnostic services of FMV. All live animals or carcasses submitted to INSA originated from the province of Quebec. After necropsy of all submitted animals and evaluation of the gross and microscopic lesions, several diagnostic tests were performed on tissue samples (such as lung and lymph nodes), including standard bacteriologic identification, PCR identification of viruses and bacteria, and PCV-2 antigen assessment by IHC. All test results and observed clinical signs reported by the veterinarians were gathered in a databank at the INSA. The following pathogens were tested: Actinobacillus pleuropneumoniae, Bordetella bronch-iseptica, Brachyspira hyodysenteriae, E. coli, H. parasuis, M. hyopneumoniae, M. hyorhinis, P. multocida, PCV-2, PRRSV, Salmonella spp., Staphylococcus aureus, Streptococcus suis with serotype identification, SIV, and a few others in very rare occasions (e.g., rotaviruses, Klebsiella spp., Lactobacillus spp., and Pasteurella spp.). A total of 121 cases submitted between October 2006 and January 2007 were selected based on the results of the PCV-2 mrtqPCR. In these cases, a positive mrtqPCR result was recorded for at least one of the 2–3 submitted live animals or carcasses, all of which were examined and tested individually. All pigs submitted for a case originated from the same farm and had the same age, which ranged between 3 and 15 weeks. In the selected population of PCVAD-compatible (PCVAD/c) cases, 40.5% had the highest PCVAD/c rank and showed moderate to severe signs of wasting or weight loss, dyspnea, coughing, pallor, and jaundice, with characteristic histopathologic lesions such as multifocal lymphohistiocytic pneumonia, lymphocytic depletion, and multinucleated giant cell formation. An additional 40.5% of cases had the lowest PCVAD/c rank and showed neither clinical signs nor histopathologic lesions (Table 1). Among the 121 PCVAD/c cases, 42 had their PCVAD status confirmed by IHC testing and 79 were IHC untested. In addition, the PRRSV and PCV-2 PCR quantification results were included in the databank generated for statistical analysis. Quantification of PCV-2 was determined with the mrtqPCR by comparing the sample results with a standard curve based on the amount of serially diluted PCV-2 recombinant plasmids (pCR2.1/PCV-2a and pCR2.1/PCV-2b). Moreover, the probe that could identify all PCV-2 genotypes (Circo-Gen-Probe) was also used to quantify PCV-2 in submitted samples. The PCV-2 quantification results were reported as the number of DNA copies/g of tissue. A PRRSV real-time PCR diagnostic kit h specific for the North American genotype of PRRSV was used for PRRSV identification as recommended by the manufacturer. The quantification of PRRSV was determined with the kit by comparing the sample results with a standard curve based on the amount of serially diluted IAF-Klop PRRSV reference strain 22 titrate after infection of MARC-145 cells and expressed as TCID50/ml. The PRRSV quantification results were reported as the number of TCID50/g of tissue. The amount of PCV-2 and PRRSV in each case was established by calculating the mean value of virus quantification of positive animals. The IHC assay was performed as previously described 53 using an anti-PCV-2 rabbit polyclonal serum (kindly provided to the MAPAQ by Dr. Dave Cavanaugh, Iowa State University). A case was considered IHC positive if tissue (lung or lymph nodes) of at least one of its submitted animals gave a positive result.
Design of the Porcine circovirus-associated disease compatible (PCVAD/c) score and distribution of clinical cases in the score system.

Statistical analyses were performed with the SAS system, version 9.1. i The relevance of the PCVAD/c score system was evaluated by establishing its statistical relationship with the PCV-2 IHC test using the Mantel-Haenszel chisquare test. Afterward, the association between the PCVAD/c score system and the presence of PRRSV was established with a likelihood ratio chi-square test; separate univariate logistic regressions were performed to test its relationship with the logarithms of the PCV-2 and PRRSV viral loads (PCV-2/vl and PRRSV/vl, respectively), which were expressed in DNA copies/g of tissue and TCID50/g of tissue, respectively. In addition, Kruskal-Wallis nonpara-metric tests were performed to detect differences in PCV-2/vl as a function of IHC result (untested, negative, and positive) in each of the 4 PCVAD/c score levels. This series of tests were performed at the α = 0.0125 level (i.e., Bonferroni adjustment) to set the overall type-1 error risk at 0.05. Then, a multivariate polytomous logistic regression 2,31 was performed to examine the concurrent effects on the PCVAD/c score of the following variables: 1) the PCV-2/vl, 2) the presence of PRRSV, 3) the PRRSV/vl, 4) IHC testing, 5) IHC result, and 6) all possible interactions of these predictor variables. Inclusion of 2 IHC-related variables allowed comparison of the differences in PCVAD/c score of the 2 subgroups, and assessment of the potential redundancy of the mrtqPCR with respect to IHC, which is a key diagnostic criterion. 51 The reference outcome value of the PCVAD/c score was set to 0 to focus on variation from clinical normality, and the cumulative logit function was used as a link between the predictors and response variables. The statistical model was built by stepwise-forward selection of the predictors, where marginal P-value thresholds of 0.15 and 0.20 were used, respectively, for introducing a predictor in the model or deleting it from the model. 31 The likelihood ratio test was performed at each step to evaluate the significance of the newly added predictor. The score test for the proportional odds assumption i was used to verify that the proportional odds model 39 was valid for analyzing the ordinal response variable. In addition, the authors dichotomized among the levels of the PCVAD/c score system (i.e., between 0 and ≥1, between ≤1 and ≥2, and so on) and performed separate binary logistic regressions to further investigate the validity of the proportional odds assumption. 3 Once the best logistic regression model was obtained, odds ratio (OR) and 95% confidence intervals (95% CI) were calculated for each significant predictor or interaction of predictors. 31 Afterward, a series of binary logistic regressions was performed to determine the odds of recovering the previously listed viral and bacterial pathogens as a function of the PCVAD/c score, the PCV-2/vl, the presence of PRRSV, and the PRRSV/vl. Relevant predictors and interactions were introduced in the model using the stepwise-forward strategy, with P = 0.15 and P = 0.20 thresholds for including and deleting predictors.
Results
Identification of the ORF2 hypervariable region by sequence analysis
Based on sequence analysis of 125 PCV-2 strains (46 PCV-2a strains and 79 PCV-2b strains), the ORF2 gene was identified to be the most variable gene between both PCV-2 genotypes, as previously described by the authors and others. 23,24 Consequently, the hypervariable region of ORF2 was targeted to differentiate both PCV-2 genotypes. The hypervariable region of ORF2 was localized between nucleotides (nt) position 222 and 279 from the gene start codon (data not shown). In the hypervariable region, 20 nt over 57 are different between both genotypes, which represents overall 35% nt variation in this region (data not shown). Because the 3'end hyper-variable region is more conserved than the 5'end nt sequence within PCV-2a strains, the 3'end region was selected to design DNA probes that could differentiate genotypes and quantify PCV-2 viruses (data not shown).
Sensitivity of Porcine circovirus-2 primers and probes
To compare the sensitivity of the mrtqPCR with that of a conventional mPCR (an assay designed to differentiate PCV-1 from PCV-2 viruses but not between PCV-2 genotypes), 46 serial 10-fold dilutions of PCV-2b FMV-06-1717 and PCV-2a IAF2897 infectious isolates were tested with both assays (Fig. 1). The mrtqPCR was determined to be at least 1,000-fold more sensitive than the mPCR (Fig. 1). The mrtqPCR standard curves were generated for both genotypes with 10-fold dilutions of quantified recombinant DNA (Fig. 2c). The coefficients of determination (R 2 ) were over 0.99 for all the regressions of the standard curves. By using the cycle threshold (Ct) values and the regression equations of those standard curves, the DNA copy number detected was calculated for each positive mrtqPCR reaction with a detection limit estimated to be 25 DNA viral genome copies/mrtqPCR reaction for all primers and probes sets (data not shown) because a nonreproducible detection of PCV-2 recombinant DNA could be observed below this value. The Ct value obtained at 25 DNA viral genome copies/mrtqPCR reaction was 37. All results with a Ct value greater than 37 were considered to be negative for PCV-2. Moreover, when the mrtqPCR was compared with a previously described PCV-2 realtime PCR assay that could not differentiate PCV-2 genotypes, 23 the sensitivity of both real-time PCR assays was similar for all mrtqPCR primers and probes sets (data not shown).
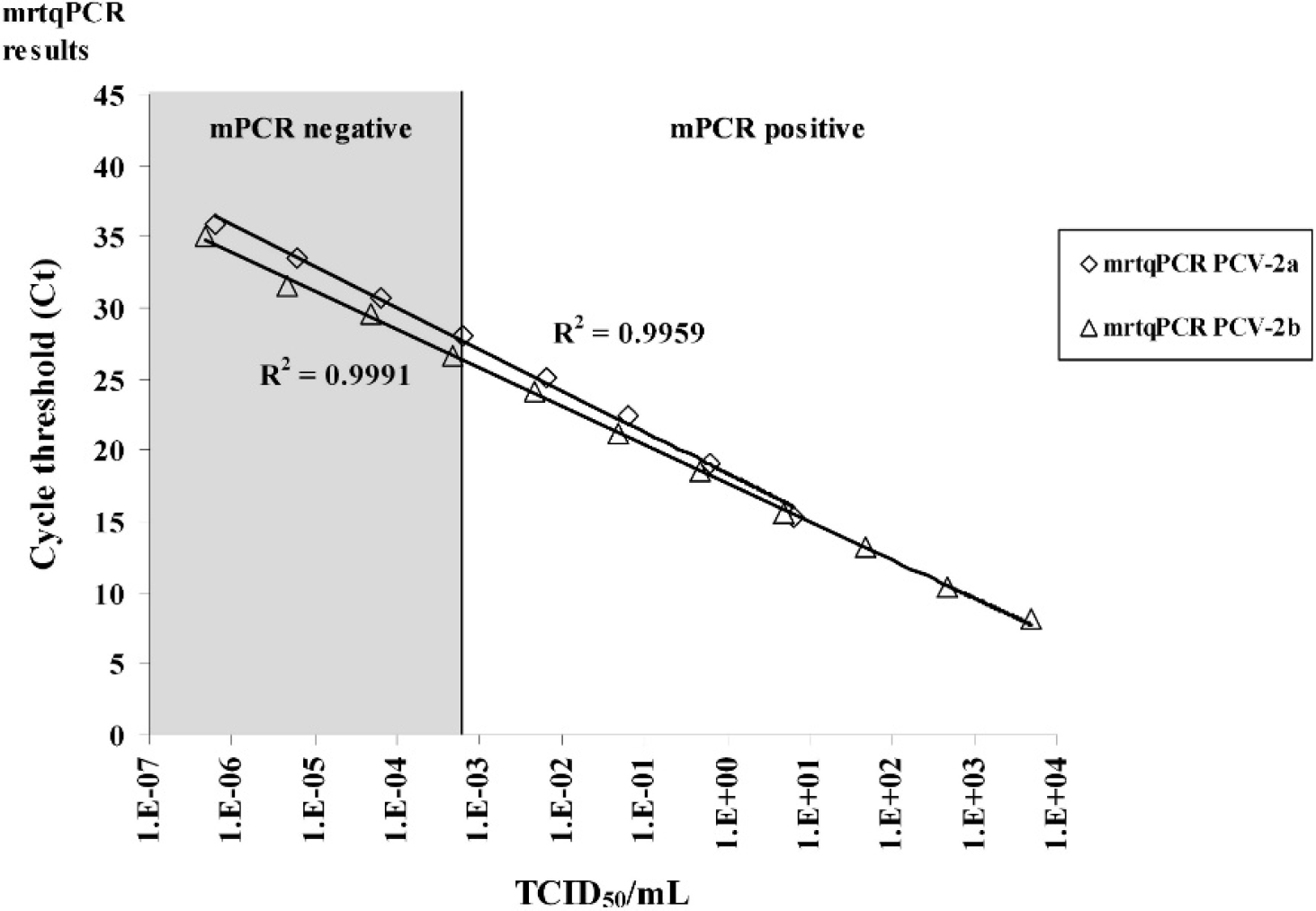
Comparison of the sensitivity of the multiplex real-time quantitative polymerase chain reaction (mrtqPCR) assay with that of a conventional multiplex PCR (mPCR) assay. Ten-fold serial dilutions of Porcine circovirus-2 genotype a (PCV-2a; strain IAF 2897) and PCV-2b (strain FMV-06-1717) were used as templates in the mrtqPCR and mPCR assays. The R 2 curve values are indicated next to their corresponding curve in the figure. The mPCR was carried out as previously reported. 44 The mrtqPCR and mPCR assays were done in triplicate, and similar results were obtained. Therefore, to improve the clarity of the figure, standard deviation results have been omitted. Ct = threshold cycle value obtained with the mrtqPCR assay; mrtqPCR PCV-2a (⋄) = PCR results obtained with the Circo-2a-Probe; mrtqPCR PCV-2b (Δ) = PCR results obtained with the Circo-2b-Probe.
Two types of targets, recombinant DNA and purified viral DNA, were used to evaluate the PCV-2 quantification efficiency of the mrtqPCR assay. From an initial concentration of pCR2.1/PCV-2b recombinant plasmid corresponding to 1.0 × 1010 PCV-2 viral genome copies/ml, 10-fold serial dilutions were made and tested using mrtqPCR. At the same time, the FMV-06-1717 PCV-2 infectious virus (4.7 × 103 TCID50/ml) was serially diluted and DNA was subsequently extracted from each dilution and tested with the mrtqPCR assay. Interestingly, the mrtqPCR standard curves obtained with the 2 different templates (recombinant DNA and purified viral DNA) were parallel with almost the same slope (Fig. 3). This indicates a very good correlation between the mrtqPCR quantification using both templates. For each time point, a 106-fold difference was consistently observed in the mrtqPCR using the 2 different templates. Overall, similar results were obtained when the mrtqPCR was performed with the pCR2.1/PCV-2a recombinant purified DNA compared with DNA purified from IAF 2897 infectious PCV-2a isolate (data not shown). The mrtqPCR appears to be more sensitive than the culture of infectious virus, as the limit of detection of the technique is under a single TCID50 (Fig. 3).
Specificity and differentiation capacity of the mrtqPCR
From a panel of RNA and DNA extracted from bacterial and viral cultures, serum, and tissues, only the samples containing PCV-2 were positive by mrtqPCR (data not shown). As illustrated in Figures 2a and 2b, the mrtqPCR assay detected each genotype specifically. Also, when recombinant DNA of both genotypes was mixed together, the mrtqPCR assay was able to identify both genotypes simultaneously (Fig. 2c).
Retrospective epidemiologic survey
Examination of the clinical samples (lungs and lymph nodes) from the authors' databank of 121 cases originating from different farms revealed that 92.56%, 4.13%, and 3.31% cases were positive for PCV-2b genotype alone, PCV-2a genotype alone, and both genotypes, respectively (Table 2). This latter result indicates the existence of PCV-2 mixed infections in Quebec swine herds, but the number of PCV-2a submitted cases recovered was too low to be able to determine whether the PCVAD/c score is affected significantly by the genotype of PCV-2 that was involved, as illustrated in Table 2 by the prevalence of PCV-2 genotypes in regards to PCVAD/c status.
The PCVAD/c score was linearly associated with the result of the PCV-2 IHC assay (Mantel-Haenszel chi-square; P < 0.0001), as an average of 93% (14/15) of cases with a score ≤1 came out negative, whereas an average of 81 % (22/27) of cases with a score ≥2 were positive (Fig. 4a). This confirmed the validity of the PCVAD/c score system. In each PCVAD/c score grade, no significant difference in PCV-2/vl was recorded for the IHC untested and IHC tested cases (Kruskal-Wallis tests; P > 0.25). In the case of PCVAD/c score = 0, the median PCV-2/vl value was 5.7735 for the IHC untested cases and 6.8348 for the IHC negative cases. In the case of PCVAD/c score = 3, the median PCV-2/vl value was 10.6821 for the IHC untested cases and 10.3729 for the IHC positive cases. These results strongly suggest that the IHC untested cases with a PCVAD/c score value of 0 are true negative PCVAD cases, and those with a score value of 3 are true positives PCVAD cases.
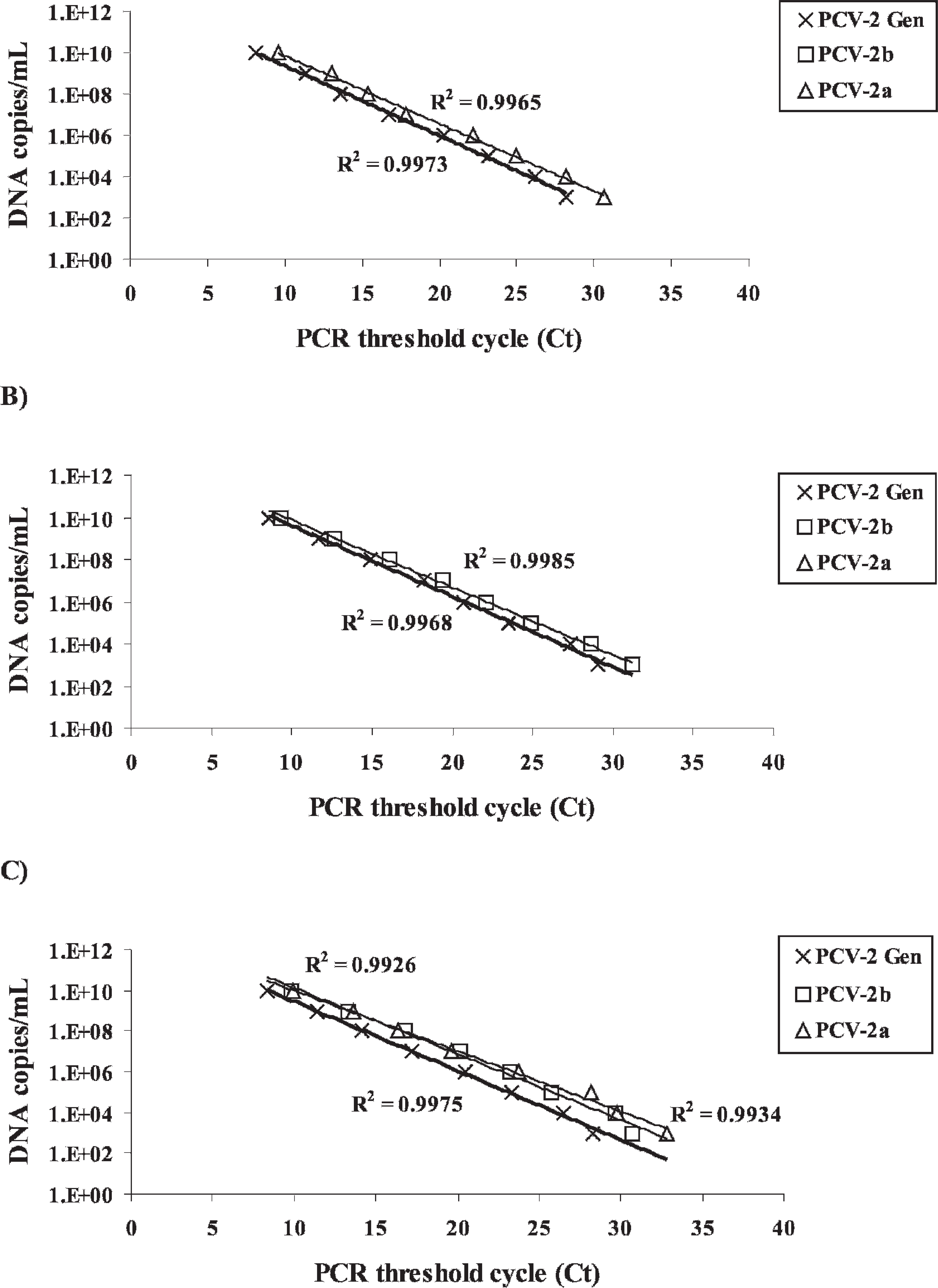
Differentiation of Porcine circovirus-2 (PCV-2) genotypes by the multiplex real-time quantitative polymerase chain reaction (mrtqPCR) assay. Ten-fold serial dilutions of PCV-2 recombinant plasmids (pCR2.1/PCV-2a and pCR2.1/PCV-2b) were used as templates in the mrtqPCR assay. The R
2 curve values are indicated next to their corresponding curve in the figure.
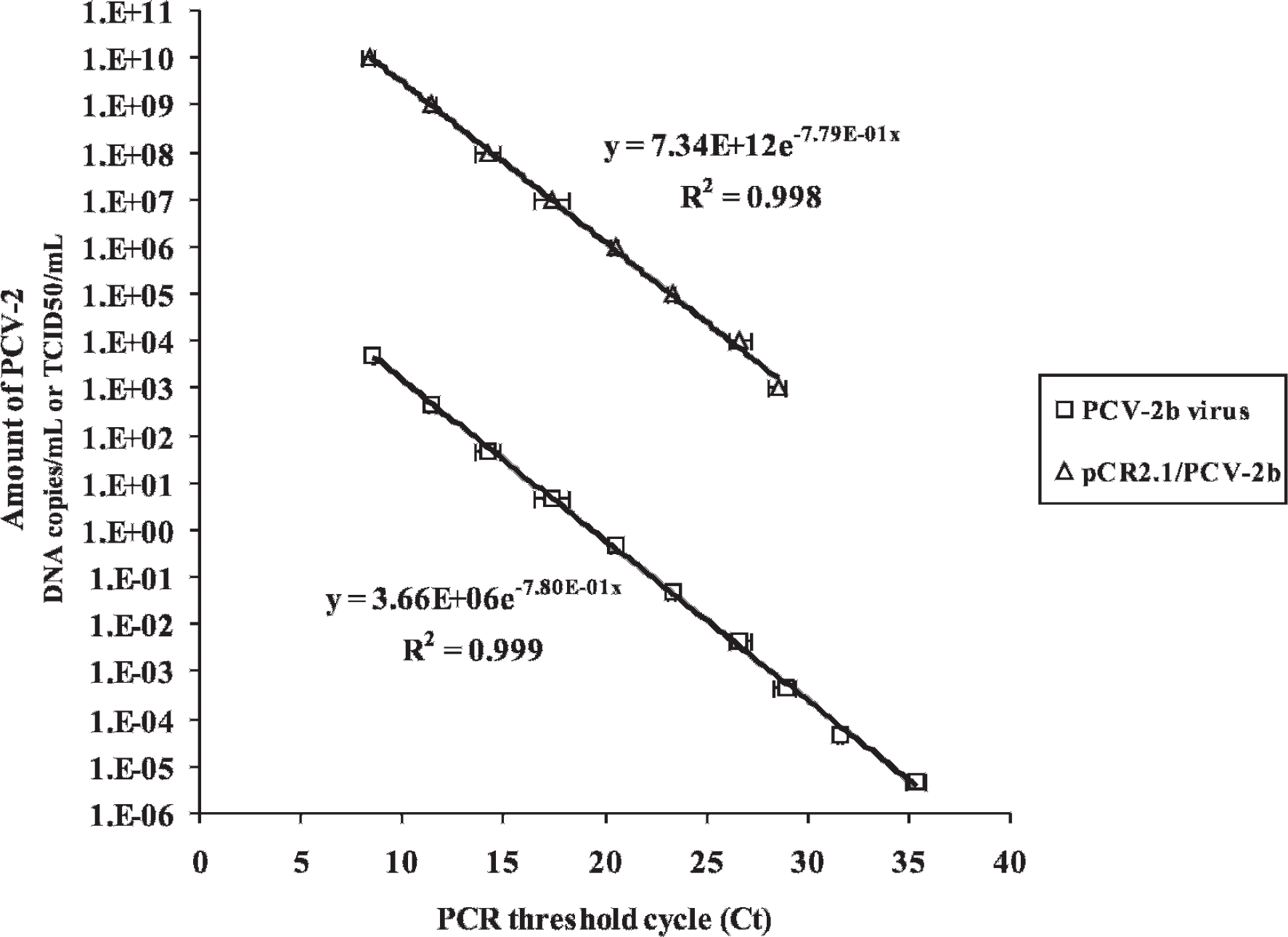
Quantification of Porcine circovirus-2 (PCV-2): infectious particles versus DNA viral genome copies. Ten-fold serial dilutions of known amounts of PCV-2b reference virus (FMV-06-1717) and PCV-2b recombinant plasmid (pCR2.1/PCV-2b) were used as DNA templates in the multiplex real-time quantitative polymerase chain reaction (mrtqPCR) assay. The amount of PCV-2b virus was expressed in TCID50/μl and the amount of pCR2.1/PCV-2b was expressed in number of DNA copies/ml. The R 2 curve values and the equations of the curve are indicated next to their corresponding curve in the figure. The data obtained in regards to PCV-2a virus and recombinant PCV-2a plasmid were not shown as the curves overlapped with those already present in the figure. Ct = threshold cycle value obtained with the mrtqPCR assay.
The univariate logistic regression analysis indicated that the value of the PCVAD/c score was significantly affected by the PCV-2/vl (P < 0.0001). To support this result, the mrtqPCR Ct mean values obtained were 30.31, 27.41, 19.74, and 15.54 for PCVAD/c grade score 0, 1, 2, and 3, respectively. Furthermore, this result is clearly illustrated in Figure 4a where a higher amount of PCV-2 correlated with a higher PCVAD/c grade score. In addition, the PCVAD/c score was significantly affected by the presence of PRRSV (likelihood ratio chi-square; P = 0.0258), as well as by the PRRSV/vl (univariate logistic regression; P = 0.0249), which granted further evaluation with the multivariate polytomous logistic regression.
The Porcine circovirus-2 (PCV-2) genotype classification results based on the new multiplex real-time quantitative polymerase chain reaction assay and their prevalence in PCV-associated disease compatible (PCVAD/c) and noncompatible cases.*
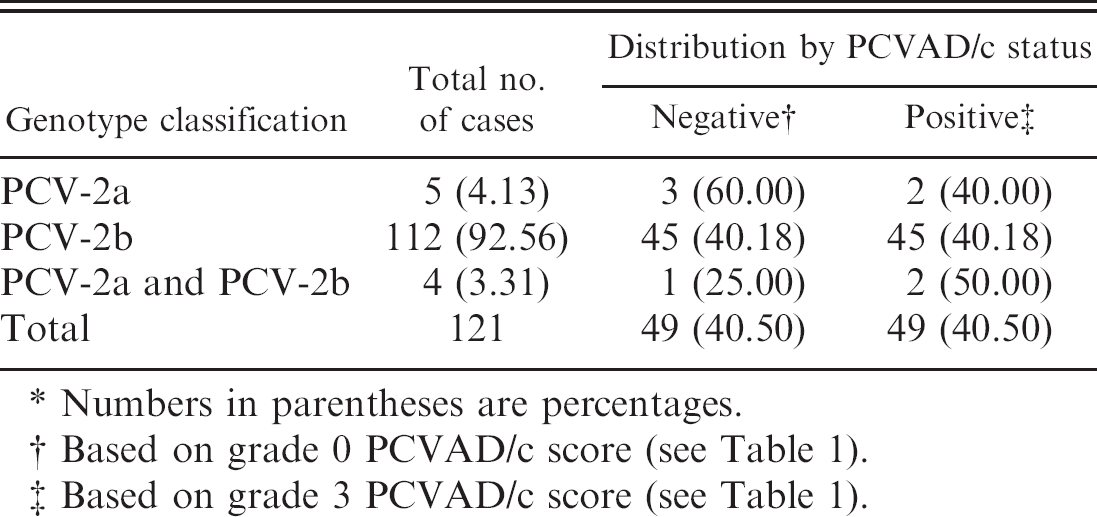
Numbers in parentheses are percentages.
Based on grade 0 PCVAD/c score (see Table 1).
Based on grade 3 PCVAD/c score (see Table 1).
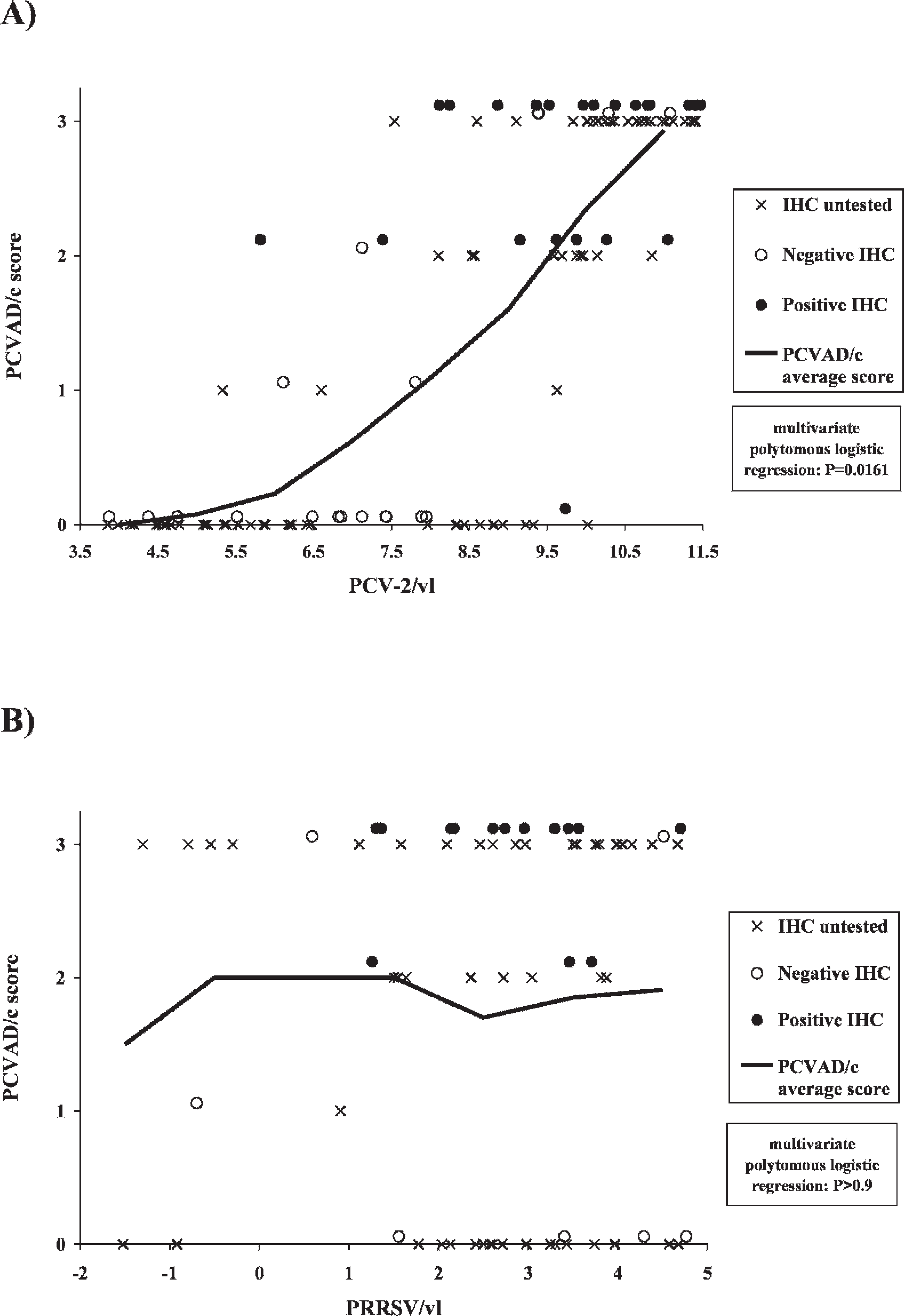
Relationship between the logarithms of the Porcine circovirus-2 (PCV-2) and Porcine reproductive and respiratory syndrome virus (PRRSV) viral loads, and the PCV-associated disease compatible (PCVAD/c) grade score classification.
Odds ratios of having a Porcine circovirus (PCV)-associated disease compatible score greater than zero, as a function of PCV-2 viral load, and result of immunohistochemistry (IHC) testing.*

PCV-2/vl = logarithm of PCV-2 viral load. Odds ratios were determined using the following reference case: negative IHC result and PCV-2/vl = 0.
This polytomous logistic regression for ordinal data, which simultaneously takes into account the previously listed variables and their interactions, revealed that the level of PCVAD/c score was significantly affected by the PCV-2/vl (P = 0.0161) and by the result of the IHC assay (P = 0.0128). In addition, a significant interaction between the results of the IHC assay and the PCV-2/vl (P = 0.0304) was recorded. But the PCVAD/c score was significantly affected neither by IHC testing (untested vs. tested; P = 0.3502), nor by the presence of PRRSV and PRRSV/vl (P > 0.9; Fig. 4b). The coefficient estimates of the final model (data not shown) revealed that IHC and PCV-2/vl considerably decreased the odds of a null score. The odds ratios of having a PCVAD/c score greater than 0 after PCV-2 PCR quantification and IHC testing are presented in Table 3. A positive IHC result increased the odds ratio of having a PCVAD/c score greater than 0 by 3.52 × 104 (Table 3). Furthermore, the PCV-2 quantification (PCV-2/vl) increased the odds ratio of having a PCVAD/c score greater than 0 by 4.08 with a PCV-2/vl log-scale unit increase (Table 3). Interestingly, both variables (IHC-positive result and PCV-2/vl) increased the odds ratio of having a PCVAD/c score greater than 0 by 5.45 × 104 per PCV-2/vl log-scale unit increase (Table 3).
Table 4 presents the effects of the PCVAD/c score, the PCV-2/vl, the presence of PRRSV, and the PRRSV/vl on the odds of recovering a number of swine pathogens from samples of a given case from the authors' databank. The prevalence of those pathogens in regards to the PCVAD/c grade score 0 and 3 (which are considered to be non-PCVAD and PCVAD cases, respectively, based on Kruskal-Wallis and multivariate polytomous logistic regression analyses) are also given in Table 4. None of these variables (PCVAD/c score, the PCV-2/vl, the presence of PRRSV, and the PRRSV/vl) had a significant effect on the odds of isolating A. pyogenes, B. hyodysenteriae, S. hyicus, or SIV (P > 0.20 in all cases). From the predictors tested, the PCV-2 viral load (i.e., PCV-2/vl) was found to be a risk factor for 2 pathogens, and the PRRSV viral load (i.e., PRRSV/vl) was found to be a risk factor for 5 pathogens. The PCV-2/vl significantly increased the risk of recovering A. pleuropneumoniae (OR = 1.887) and S. suis serotypes 1/2, 1, 2, 3, 4 and 7, which are serotypes commonly associated with clinical diseases 29 (S. suis pathogenic serotypes; OR = 1.3690). In contrast, PCV-2/vl had no significant effect on the odds of isolating S. suis when all of its serotypes were included (i.e., generic S. suis). The odds of isolating M. hyopneumoniae were significantly higher in pigs with a PCVAD/c score of 2, compared with the reference population with a PCVAD/c score of 0(OR= 7.800). Nevertheless, PCV-2 had a null or a significantly negative effect on the risk of recovering other pathogens, especially B. bronchiseptica, E. coli, and Salmonella spp. On the contrary, PRRSV significantly increased the risk of recovering B. bronchiseptica, E. coli, H. parasuis, P. multocida, and generic S. suis (PRRSV/vl; OR = 1.305, 1.479, 1.443, 1.474, and 1.203, respectively). In 2 cases, the PCV-2 viral load had opposite sign coefficients compared with PRRSV viral load, suggesting that their combined effect was not additive.
Effect and statistical significance of the Porcine circovirus (PCV)-associated disease compatible (PCVAD/c) score, PCV-2 viral load, and presence and viral load of Porcine reproductive and respiratory syndrome virus (PRRSV) on the odds of isolating common swine pathogens.*
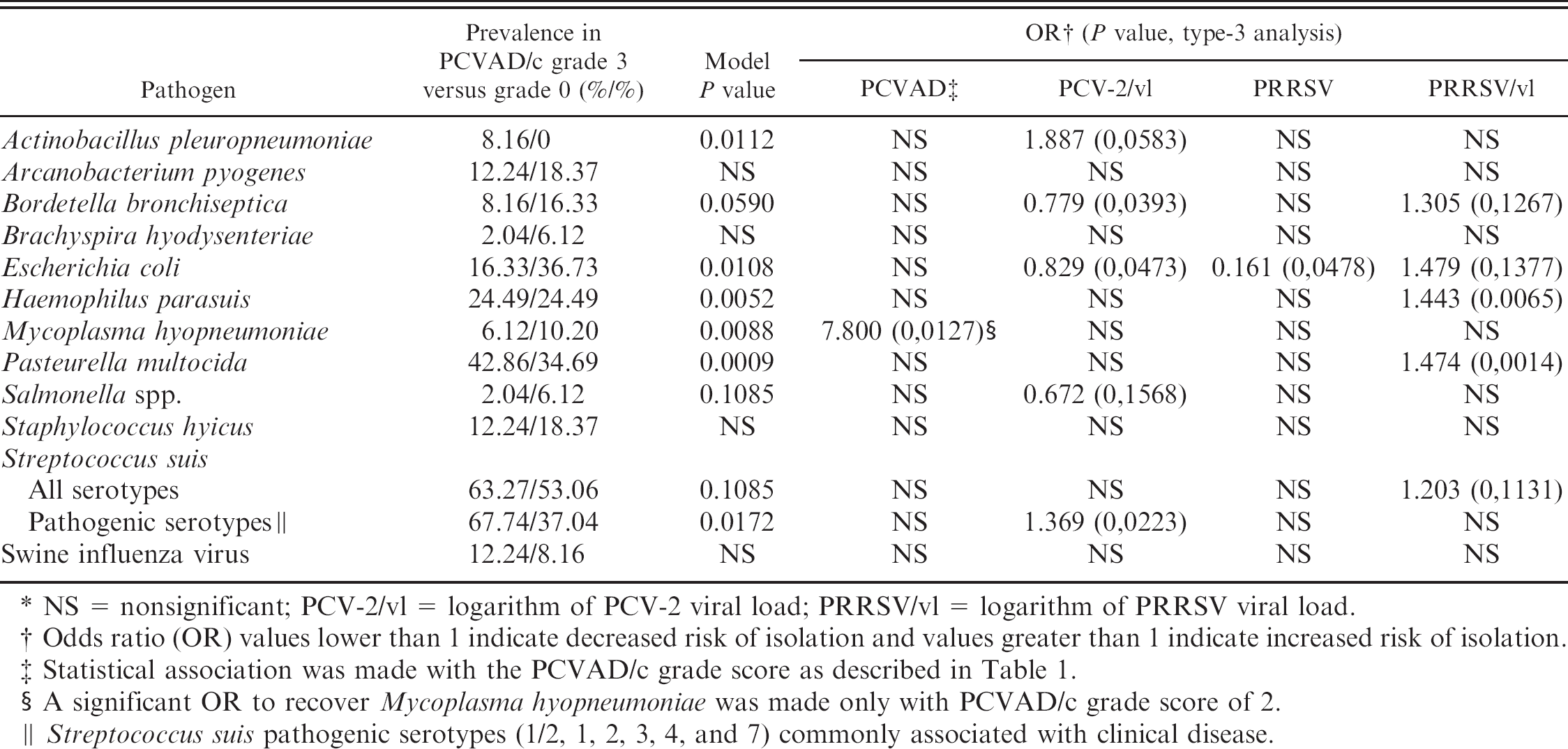
NS = nonsignificant; PCV-2/vl = logarithm of PCV-2 viral load; PRRSV/vl = logarithm of PRRSV viral load.
Odds ratio (OR) values lower than 1 indicate decreased risk of isolation and values greater than 1 indicate increased risk of isolation.
Statistical association was made with the PCVAD/c grade score as described in Table 1.
A significant OR to recover Mycoplasma hyopneumoniae was made only with PCVAD/c grade score of 2.
Streptococcus suis pathogenic serotypes (1/2, 1, 2, 3, 4, and 7) commonly associated with clinical disease.
Discussion
One of the main objectives of the swine industry and health scientists is to establish if there is virulence variation between PCV-2a and PCV-2b viruses. In fact, a recent retrospective study 5 showed a statistically significant increase in the frequency of PCVAD characteristic histologic lesions observed in lymph node, spleen, lung, small intestine, colon, and kidney in pigs infected with PCV-2b strains compared with pigs infected with PCV-2a strains. Viral burden, as estimated with IHC staining in lymph node, was also significantly increased in pigs infected with the PCV-2b strains compared with the PCV-2a strains. 5 Because these results were derived from retrospective analysis of cases submitted to diagnostic laboratories, the conclusion may be biased. Nonetheless, it strongly suggests that PCV-2b virus could be more virulent than PCV-2a virus. Moreover, others have reported that genetic variations may have a great impact on the virulence of PCV-2. 21 Consequently, it has become critical for swine producers and veterinarians to establish which PCV-2 genotypes are present in their herds. In that context, an mrtqPCR assay was developed to sensitively identify and differentiate PCV-2 viruses in clinical samples (Figs. 1, 2; Table 2). With the exception of the conventional PCR assay developed previously, 30 all other PCR assays reported until now did not differentiate between PCV-2 genotypes. 4,12,34,55 Moreover, the conventional PCR diagnostic test reported previously 30 required 2 PCR reactions to identify both genotypes whereas only 1 reaction is required in the authors' mrtqPCR. Furthermore, real-time PCR assays are usually more sensitive than conventional PCR assays, as illustrated in Figure 1.
Interestingly, a good correlation was established with the mrtqPCR assay between 2 quantification methods, the DNA copies/ml versus the TCID50/ml (Fig. 3). The relationship between the 2 PCV-2 quantification methods (DNA copies vs. infectious virus) was determined to be DNA copies/ml value = (1.76 × 106) × TCID50/ml value (Fig. 3). It is unknown whether this equation could be generalized to all PCV-2 isolates, as new recombination or mutations of the viral genome within the mrtqPCR targeted regions could interfere with both quantification and genotyping results. Furthermore, PCR inhibitors in clinical specimens could adversely affect the PCV-2 quantification. Nonetheless, it will help veterinarians in their interpretation of the diagnostic results obtained with the mrtqPCR assay.
The standard PCVAD definition consists of three criteria: 1) presence of characteristic clinical signs, 2) presence of characteristic histopathologic lesions in lymphoid tissues, and 3) detection of moderate to high amount of PCV-2 within the lesions in lymphoid and other tissues by different techniques such as IHC. To statistically test the respective roles of PCV-2 viral load and the result of IHC testing in regards to the PCVAD status of the clinical cases, it was purposely decided not to apply the previous definition. 51 Specifically, the third criterion was omitted in the PCVAD case definition as detailed in Table 1. Of note, both the Kruskal-Wallis and multivariate polytomous logistic regression analyses revealed that at identical PCV-2/vl values, the IHC untested cases had similar PCVAD/c score grades compared with IHC negative and positive cases. Consequently, these results indicated that the PCVAD/c grade 0 and 3 cases are not significantly different from their IHC-tested counterparts (standard negative and positive PCVAD cases), and strongly suggests that both groups are identical.
A very strong logistic (sigmoid) relationship was recorded between the PCVAD/c grade score and the result of IHC assay, as well as the logarithm of the amount of PCV-2 virus recovered from the tissue samples using the mrtqPCR assay and the interaction between these 2 independent variables (Fig. 4a). Interestingly, the stepwise-forward logistic regression analysis suggests that the PCV-2 viral load may be a reliable alternative to the IHC assay as the third criterion of the PCVAD definition since at PCVAD/c grade 3 score, the lowest amount of PCV-2 detected was 107.5 DNA copies/g of tissue, at which the OR of having PCVAD/c was established to be 1/e(−1.4057 × 7.5) = 3.79 × 104 (95% CI = 1/e(−1.4057×7.5±1.96×7.5×0.2180) = [1.53 × 103−9.34 × 105]), which favorably compares to the OR and 95% CI of a positive IHC result (Table 3, scenario 2). Moreover, with the median PCV-2/vl value of the grade-3 score, the resulting OR and 95% CI are 3.11×106 and [3.31 × 104−2.93 × 108], respectively. These results are not surprising because PCR has been shown to be more sensitive than IHC. Indeed, other investigators have reported that from 20 PCV-2 experimentally infected pigs, 0% were PCR negative but 5% were IHC negative when testing lymph nodes, and 10% were PCR negative but 44% were IHC negative when testing the lung tissue. 32 Furthermore, from 88 pigs tested in a commercial herd with severe PMWS, only 3% were PCR negative but 23% were IHC negative. 40 The latter result is similar to our findings, as 4 of the 19 (21%) cases with PCVAD/c score = 3 were IHC negative despite showing high PCV-2 viral load at the mrtqPCR testing.
This relationship between PCV-2 viral load and clinical expression of PCVAD confirms results previously reported. 4,44 Consequently, the higher the PCV-2 viral load in tissue samples, the higher the probability to have PCVAD (Fig. 4a). This remains a probability as a high amount of PCV-2 viral load has been observed in some cases unrelated to PCVAD and vice versa (Fig. 4a). Surprisingly, the stepwise-forward logistic regression analysis revealed that a positive IHC result significantly increased the odds of clinical disease, even if PCV-2 viral load is already present in the model (Table 3). These results suggest that PCV-2 viral load and IHC testing may represent 2 separate components of the overall disease process, but their statistically independent biological meaning could not be determined within the retrospective study.
In the present study, the prevalence of PCV-2b virus was very high, as previously reported. 23 In fact, 95.9% of the submitted cases were infected with PCV-2b virus (Table 2), even though 40.5% of those submitted cases had no clinical sign and microscopic lesion related to PCVAD. This suggests that not all animals infected with PCV-2b virus may develop PCVAD. However, the difference observed between the presence of PCVAD and the number of PCV-2b cases may be due to the stage and time course of infection when the tissue samples were collected. As reported previously, 30 PCV-2a and -2b mixed infections were also observed but at a lower prevalence in Quebec province (3.31%) compared with that in a Kansas study (62.5%). The difference between prevalence values could be due to specimen origin because samples were obtained from pigs experiencing severe PCVAD in the Kansas study, 30 whereas in the present study almost half of samples were obtained from animals that had no PCVAD clinical signs (Table 1). Nonetheless, even if the disease evolution after PCV-2 mixed infections is unknown, it may have an effect as recombination between PCV-2 genomes has been reported. 28,37,43
The course of PCV-2 infection that will lead to the occurrence of the disease has been associated with the presence of other pathogens, such as M. hyopneumoniae, Porcine parvovirus, PRRSV, and SIV. 16,18,45,48 Thus, it was not surprising to find at the preliminary stage of data analysis, that a significant association existed between the presence of PRRSV and the appearance of PCVAD, as previously reported. Similar to the correlation between PCV-2 viral load and PCVAD/c (Fig. 4a), there was a significant association between PRRSV viral load and the appearance of PCVAD. In contrast, the multivariate polytomous logistic regression analysis suggested that the presence and viral load of PRRSV had no significant effect on the PCVAD/c score, as shown in Figure 4b, when both PCV-2 viral load and IHC result parameters are included in the regression model. This suggests that PRRSV may be less involved as a cofactor responsible for the induction of PCVAD than expected. 18,48 Indeed, animals with PCVAD may be PRRSV negative or have PRRSV low titers (Fig. 4b). In contrast, animals that have no PCVAD may possess a high viral load of PRRSV (Fig. 4b). Because of bias inherent to a retrospective epidemiologic survey, caution must be taken in interpreting these results.
In contrast to what has been reported previously, 16 epidemiologic assessment of the OR of PCV-2 coinfection with other swine pathogens in the present study did not involve SIV as an important cofactor (Table 4). This could be explained by the fact that the number of SIV-positive cases was very low in this studied population. Interestingly, statistical analyses revealed a high OR (OR = 7.800) of recovering M. hyopneumoniae in PCVAD/c grade 2 score animals, which represents animals with PCVAD-specific microscopic lesions without having PCVAD clinical signs (Tables 1, 4). In a previous report, 16 a 3.77 OR of recovering M. hyopneumoniae was found in PCV-2-positive animals, irrespective of their PCVAD status. Surprisingly, no association was found in the present study between M. hyopneumoniae and PCVAD/c grade 3 score, which corresponds to pigs that show PCVAD clinical signs with the presence of PCVAD-specific histopathologic lesions. Consequently, even if there is a good chance of identifying M. hyopneumoniae in PCV-2-positive animals, the present study did not involve M. hyopneumoniae in animals that have clinical signs related to PCVAD (Table 4). The most interesting finding was the significantly increased ORs of isolating 2 major porcine respiratory pathogenic bacteria, A. pleuropneumoniae and S. suis pathogenic serotypes, as PCV-2 viral load increases (Table 4). As opposed to previous studies, 16 the present study did distinguish between serotypes of S. suis. Herein, association with PCV-2/ vl was significant only for the pathogenic serotypes of S. suis (Table 4). Yet, despite a good statistical relationship between A. pleuropneumoniae, S. suis pathogenic serotypes, and PCV-2 viral load, it is impossible to determine if those bacterial pathogens increased the risk of PCVAD development or if they colonized the host respiratory tract after PCV-2 infection.
In conclusion, the new mrtqPCR diagnostic assay is suitable for the sensitive identification and differentiation of PCV-2 and is a more convenient approach than RFLP, partial or entire viral genome sequencing, and conventional PCR techniques. 6,23,30 Furthermore, it could efficiently quantify PCV-2 in submitted samples and provide a risk assessment in regards to the odds of developing PCVAD (Fig. 4a). Until now, it was believed PRRSV played a major role in PCVAD development, and experimental infections with both pathogens (PRRSV and PCV-2) may confirm that. 50 Nevertheless, if there is apparent statistical relationship with PCVAD and PRRSV in the present report, the impact of PRRSV in regards to PCVAD seems negligible, as previously reported, 52 when statistical analysis takes into account other relevant variables like the PCV-2 viral load. Since the odds of isolating A. pleuropneumoniae and pathogenic S. suis is significantly higher when the PCV-2 viral load increases, new experiments are now being conducted to evaluate the impact of PCV-2 and the mechanisms involved on the bacterial colonization of the swine respiratory tract.
Acknowledgements
The authors are grateful to Cynthia M. Guilbert for critically reviewing the manuscript. This work was supported by the National Sciences and Engineering Research Council of Canada (NSERC) discovery grants, the Fédération des producteurs de porcs du Québec (FPPQ), the Centre d'insémination porcine du Québec Inc. (CIPQ), and the Conseil pour le développement de l'agriculture du Québec (CDAQ). The authors are also extremely grateful to the Ministère de l'Agriculture, des Pěcheries et de l'Alimentation du Québec (MAPAQ) and its subsidiary, the Institut National de Santé Animale (INSA), for permission to access complete diagnostic result files to compile the databank that was used in the retrospective epidemiological survey.
Footnotes
a.
Beckham Coulter Cananda Inc., Mississauga, Ontario, Canada.
b.
Qiagen Inc., Valencia, CA
c.
Invitrogen Canada Inc., Burlington, Ontario, Canada.
d.
BioSpec Products Inc., Bartlesville, OK.
e.
Ibis Therapeutics, Carlsbad, CA.
f.
Roche Diagnostics, Laval, Quebec, Canada.
g.
Cepheid Inc., Sunnyvale, CA.
h.
Tetracore Inc., Rockville, MD.
i.
SAS Institute Inc., Cary, NC.