
Abstract
Fifty-six dogs from St. John's, Newfoundland, Canada, were evaluated for Angiostrongylus vasorum infection. Small numbers of nematodes were found within pulmonary arteries of 6 dogs. Larvae were identified in fecal samples in 2 of 6 dogs. All 6 dogs had multifocal granulomatous pneumonia and sometimes foci of chronic thrombosis, which varied from very mild to severe. One dog had extensive pulmonary lesions resulting in cor pulmonale. Right heart failure was characterized by right ventricular hypertrophy, hepatic congestion, ascites, and hydrothorax. Microscopically, in most cases, eggs, larvae, and sometimes intravascular adults, were present within lung tissue sections. Small foci of granulomatous inflammation with and without larvae were present in kidney and brain in 4 dogs. An additional dog, diagnosed antemortem with angiostrongylosis via fecal examination, was also examined. Pathological findings consisted of severe pyogranulomatous interstitial pneumonia with myriad eggs, larvae, and numerous intravascular pulmonary adult nematodes with extensive arterial thrombosis. Five hundred and seventy-two adult worms were removed from pulmonary arteries. Foci of granulomatous inflammation, often associated with larvae and/or eggs, were present in tracheobronchial lymph nodes, adrenal gland, brain, and kidneys. Severe seizuring noted antemortem was attributed to several large, discrete areas of acute hemorrhagic infarction within the cerebrum and cerebellum. Natural A. vasorum infection in domestic dogs in eastern Newfoundland causes lung pathology of variable severity, which in some cases, may progress to cor pulmonale and which may be associated with extrapulmonary lesions and clinical signs.
Introduction
Angiostrongylus vasorum, or French heartworm, is a metastrongylid nematode. The adult parasite infects the pulmonary arteries of the definitive host, which is most commonly a wild or domestic canid. Red fox and other species of wild fox serve as natural definitive hosts and are an important reservoir of infection for dogs. 4,26 Natural infection has also rarely been reported in other species including Eurasian badgers (Meles meles), wolves (Canis lupus), and a coyote (Canis latrans). 6,28,33 The life cycle of A. vasorum is indirect. Gastropods (aquatic and terrestrial snails, slugs) most frequently serve as intermediate hosts. 4 Frogs can also serve as paratenic and intermediate hosts. 2 In the definitive host, adult female nematodes in pulmonary arteries shed eggs that are subsequently transported downstream to smaller capillaries in the lung parenchyma. The eggs develop and hatch, releasing first stage larvae (L1) that penetrate capillary and alveolar walls, and move into alveoli and larger airways. Larvae are subsequently coughed up into the pharynx, swallowed, and then excreted in the feces. Gastropods become infected with L1 when foraging on infected feces or on contaminated plant material. In the intermediate host, the larvae mature and develop into second-stage and third-stage larvae (L2 and L3). The final host, usually a fox or dog, becomes infected by eating an intermediate host containing L3. The slug or snail is digested within the stomach or intestine of the canid, releasing L3 that penetrate the gastrointestinal tract wall, migrate to visceral lymph nodes, and eventually develop into immature adults. The juvenile worms migrate via portal circulation to the liver, caudal vena cava, the right atrium, right ventricle, and pulmonary arteries where they reach maturity approximately 33 to 35 days postinfection. The prepatent period for A. vasorum is approximately 38 to 57 days. 4
Angiostrongylus vasorum was first observed in the right heart and pulmonary arteries of dogs in France in the 1800s and was referred to as French heartworm. 26 At present, angiostrongylosis is considered endemic in discrete areas of Europe, including parts of France, southwestern England, Ireland, Denmark, Italy, Spain, Germany, Hungary, Finland, Switzerland, and Turkey, in South America in Brazil and Colombia, and in Uganda on the continent of Africa. 3,22,31,32 Until the late 1990s in North America, A. vasorum infection was relatively rare and was reported largely in domestic dogs that had been imported from or had traveled in Europe. 21 However, in the last decade, a relatively new endemic focus of A. vasorum infection has emerged in eastern Canada, in the provinces of Newfoundland and Labrador. The parasite was first identified in foxes in Newfoundland in a helminth survey in 1973. 29 More recent studies indicate a relatively high infection rate in foxes limited to the southeastern part of the island of Newfoundland, specifically in the Avalon and Burin Peninsulas and along the eastern coast from Hamilton to Bonavista Peninsula. 14 Since 1996, dogs from this area have been diagnosed with angiostrongylosis with increasing frequency via fecal examination using the Baermann technique or at postmortem. 5,9 The purpose of this report is to describe the pathological findings in a group of dogs naturally infected with A. vasorum from this relatively new North American endemic focus.
Materials and methods
A postmortem survey of dogs surrendered to the St. John's, Newfoundland, branches of the Humane Society, and the Society for the Prevention of Cruelty to Animals (SPCA) was conducted over the 3-year period of May 2001 to December 2003. Unfortunately, histories and antemortem clinical examination of these dogs was not obtained. All dogs were surrendered by people living in the city of St. John's, Newfoundland, or the immediately surrounding area. Dogs were humanely euthanized with an intravenous overdose of barbiturate. Dogs were examined over 3 collection periods extending from May to August 2001, November 24 to 29, 2002, and December 1 to 7, 2003.
Postmortem examination was conducted within 24 hours after death. An estimate of age was made. Gender, body weight, heart weight, and measurements of thickness of left and right ventricular free walls taken from below the atrioventricular valves were recorded. Complete postmortem examinations were performed on each dog, and samples of lung, heart (left and right ventricular free wall), liver, tracheobronchial lymph node, kidney, and brain were fixed in 10% buffered formalin. Additional tissue samples were taken of any visible abnormalities identified at necropsy. In each dog, pulmonary arteries were opened and examined from the base of the heart to the narrowest branches possible at the margins of all lung lobes in order to identify intravascular nematodes. When present, worms were collected, identified as described previously, 26 and counted. Rectal fecal matter was collected from each dog and examined using the Baermann technique. 15 Dogs were identified as infected when parasites were found within pulmonary arteries or in feces. Formalin-fixed samples from A. vasorum-positive dogs were processed for sectioning, embedded in paraffin, sectioned, and stained with hematoxylin and eosin (HE).
An additional A. vasorum-infected dog was submitted for postmortem examination during the second collection period (2002) from a veterinary clinic in St. John's, Newfoundland. This dog had been diagnosed with angiostrongylosis antemortem via fecal examination and was euthanized with an intravascular overdose of barbiturate as a result of posttreatment complications. This dog was processed in the same manner as that described in the postmortem survey study.
Results
Over a 3-year period, a total of 56 dogs from the St. John's, Newfoundland Humane Society, and SPCA were examined. From May to August 2001, 30 dogs were examined. Intravascular adult nematodes identified as A. vasorum were found in the pulmonary arteries of 4 of these dogs (dogs 1 to 4). Fecal Baermann tests were positive in 2 of these dogs (dogs 2 and 3). From November 24 to 29, 2002, 18 dogs were sampled. Intravascular nematodes were found in 2 of these dogs (dogs 5 and 6). Fecal Baermann tests did not reveal larvae in either dog. Eight dogs were sampled from December 1 to 7, 2003. Intravascular pulmonary nematodes were not identified in any of these dogs, and Baermann fecal examinations were negative. Gender, age estimates, breed, fecal Baermann test results, body and heart weights, and measurements of left and right ventricular thickness for A. vasorum-infected dogs are given in Table 1. Postmortem findings are summarized in Table 2.
Adult female and male nematodes removed from pulmonary arteries were approximately 1.5–2 cm long, thin, and thread-like. The elongate white ovaries of females often wound around the darker, red intestinal tract giving the worms a “barber pole” appearance. Nematodes were identified as A. vasorum based on the adult male worm bursa morphology and spicule length. 26 First-stage larvae collected from feces using the Baermann technique were approximately 350 μm in length, had a cephalic button, and a kinked tail with a dorsal spine. In histologic sections, cross-sections of adult nematodes were approximately 270–350 μm in diameter and had a thin, smooth cuticle, a thin hypodermis with lateral cords, coelomyarian musculature, and intestines lined by tall cuboidal epithelium. Sections of gonad were also often present (Fig. 1). Larvae were approximately 10-μm thick. Eggs were thin shelled and varied from 80–110 μm in diameter. Immature eggs contained small basophilic and eosinophilic granules. Embryonated eggs contained a coiled larva.
Summary of gender, age estimates, breed, fecal Baermann results, body and heart weights, and left and right ventricular thickness for Angiostrongylus vasorum-infected dogs.*
BW = body weight; FS = female, spayed; MN = male, neutered; A = adult.
Dog diagnosed with cor pulmonale.
Dog presenting with neurological clinical signs.
The lung lesions in infected dogs were classified as mild, moderate, or marked based on the severity of microscopic and gross lesions. Two dogs (dogs 1 and 5) exhibited mild lung changes. Grossly, the lungs of these dogs were unremarkable and very few worms (<3) were found within pulmonary arteries. Microscopically, mild, rare, multifocal, chronic inflammatory lesions, most prevalent at the lung lobe margins, were present in both dogs. These foci consisted of small areas of interstitial fibrosis containing mild infiltrates of lymphocytes, plasma cells, macrophages, sometimes laden with hemosiderin, and rare foreign body-type multinucleated giant cells (MNGCs). In dog 5, a small pulmonary artery was partially occluded by an organized and recanalized thrombus. No parasites were visualized.
The lung lesions in dog 2 were classified as moderate. Grossly, the lungs were unremarkable. Several adult parasites were found within pulmonary arteries. Alveolar septa were multifocally, mildly to moderately expanded, with increased amounts of dense fibrous tissue containing small-to-moderate numbers of lymphocytes, plasma cells, macrophages, occasional MNGCs, and eosinophils. These changes appeared most prominent in apical and subpleural regions, where the pleura were often mildly to moderately thickened with dense fibrous stroma. Numerous primitive and embryonated nematode eggs and nematode larvae were frequently embedded within inflammatory infiltrates. Clusters of hemosiderin-laden macrophages were often scattered within the interstitium. Primitive eggs were occasionally present within arteries.
Summary of postmortem findings in Angiostrongylus vasorum-infected dogs.
Dogs 3, 4, and 6 had lung lesions that were considered severe but had slightly variable histologic features. Small numbers of parasites (on average 5 to 7 worms) were recovered from pulmonary arteries. Macroscopically, the lungs of affected dogs were rubbery to firm with multifocal firmer areas near the edges of lung lobes. These foci were poorly defined and dark red to yellow-brown. In dog 6, 2, discrete, chalky, white, lesions were located at the edges of the right middle lobe. Lungs varied in color from diffusely pale pink to dark red, to mottled red and pink. All failed to collapse.
In dog 3, sections of firmer areas of lung represented areas where the pulmonary parenchyma was effaced by multifocal to coalescing, discrete granulomas characterized by aggregates of epithelioid macrophages, lymphocytes, plasma cells, fibroblasts, and occasional MNGCs (Fig. 2). Granulomas often contained small central deposits of amorphous, pale, eosinophilic, necrotic debris occasionally admixed with small amounts of mineral. Primitive and embryonated nematode eggs and larvae were sometimes embedded within these inflammatory foci (Fig. 2, inset). Surrounding alveolar septa were moderately to markedly thickened with fibrous tissue containing mild-to-moderate lymphohistiocytic infiltrates. One medium-sized artery contained a cross-section of an adult nematode (Fig. 1). Less severely affected sections of lung were moderately congested, but minimal inflammation was evident.
The microscopic lesions in the lungs of dog 4 were similar in distribution to dog 3 but appeared to be more chronic. Numerous, multifocal to coalescing, caseogranulomas consisting of discrete aggregates of amorphous, pale, eosinophilic debris interspersed with granular, mineral, and degenerate remnants of parasite eggs and larvae surrounded by a thin rim of epithelioid macrophages, MNGCs, and fibroblasts with fewer lymphocytes, and plasma cells replaced large areas of parenchyma. These caseogranulomas often distorted and compressed the adjacent intact tissue (Fig. 3). The overlying pleura were moderately thickened with dense fibrous tissue. Pulmonary arteries had markedly hypertrophied tunica media and narrowed lumina. Generalized, often prominent, interstitial fibrosis was present, containing clusters of hemosiderin-laden macrophages and occasional infiltrates of lymphocytes and plasma cells.
Histologic sections of lung from dog 6 were similar in appearance to dog 5, but the interstitial fibrosis present was more severe, diffuse, and accompanied by similar mononuclear inflammatory cell infiltrates. Clusters of macrophages, often laden with hemosiderin, were common. Cross-sections of many large pulmonary arteries were chronically thrombosed and recanalized with small foci of mineralization. Interstitial and vascular smooth muscle hyperplasia and hypertrophy were prominent. The lumina of many affected blood vessels were narrowed (Fig. 4). Alveoli were irregularly distended with pale eosinophilic fluid interspersed with large foamy macrophages and small scattered aggregates of fibrin. The pale, white foci described grossly represented large, discrete caseogranulomas similar to those described in dog 4. Several smaller caseogranulomas were present in the surrounding parenchyma. These foci sometimes contained the remains of degenerate nematode eggs.
Additional prominent gross and microscopic lesions were evident in dog 6. The abdomen was distended with approximately 300–500 ml of red watery fluid. The liver was diffusely enlarged, dark red with rounded edges, and weighed 700 g (approximately 2 to 3 times larger than normal). Histologically, there was prominent, centrolobular, and bridging sinusoidal congestion in the liver. Lymphatics surrounding sublobular veins were often moderately distended with clear-to-pale, pink edema fluid. This finding was consistent with marked, passive, hepatic congestion. Rare acute, fibrin thrombi were present within portal veins. The heart was moderately enlarged and rounded (0.92% of body weight). The pulmonary trunk was prominent and bulging. The right ventricular free wall was thickened, and trabeculae carneae muscles were hypertrophied. There was mild dilation of the lateral ventricles of the brain. Large caseogranulomas effaced approximately 85% of the right adrenal medulla. Fibrin thrombi and small mononuclear inflammatory foci were scattered in the adrenal cortex.
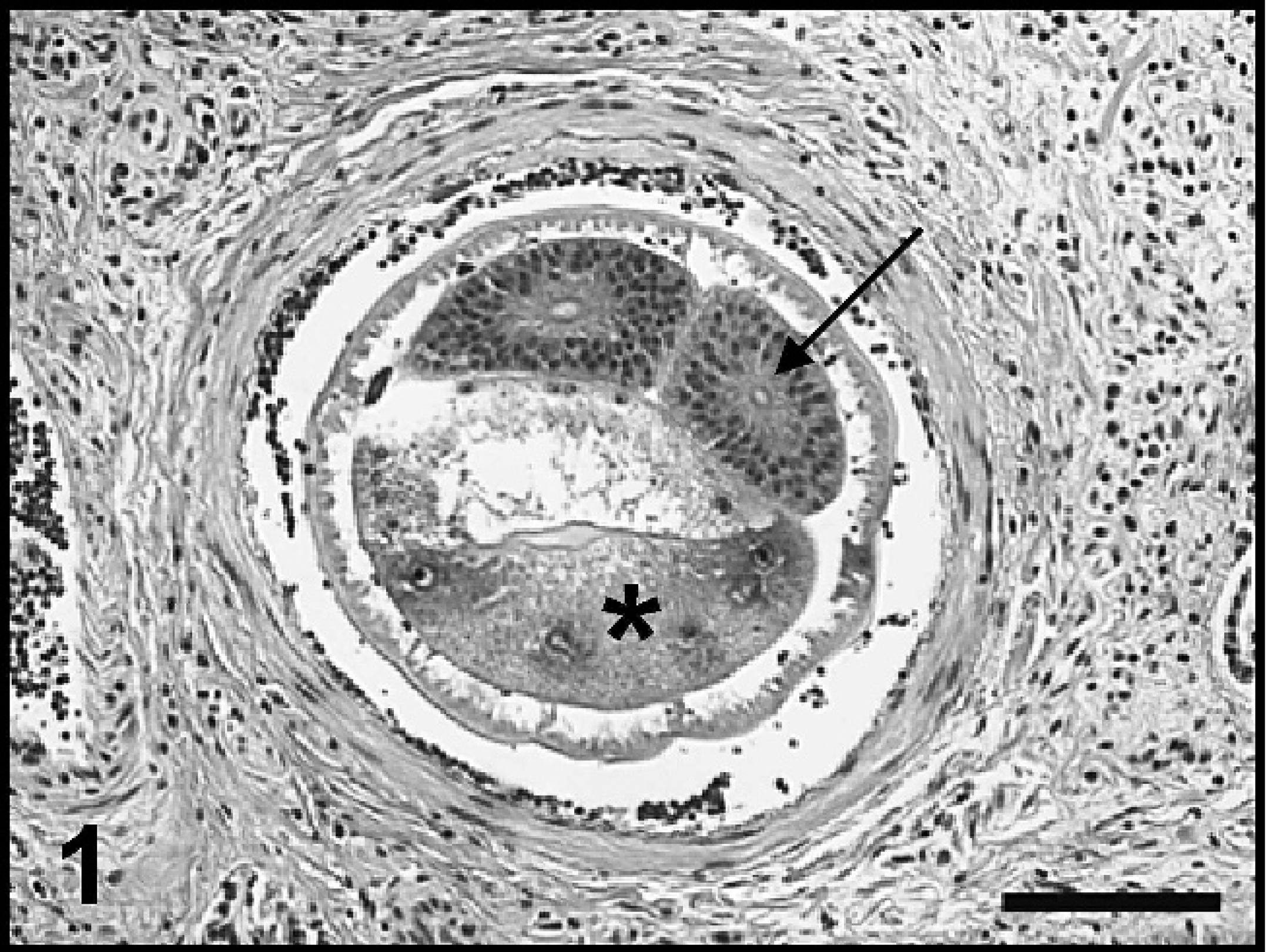
Lung, dog 3. Adult A. vasorum nematode within a pulmonary artery, the intestine (asterisk), and gonads (arrow) are indicated. Hematoxylin and eosin. Bar = 100 μm.

Lung, dog 3. Multifocal-to-coalescing granulomas replace large areas of pulmonary parenchyma. Hematoxylin and eosin (HE). Bar = 500 μm.
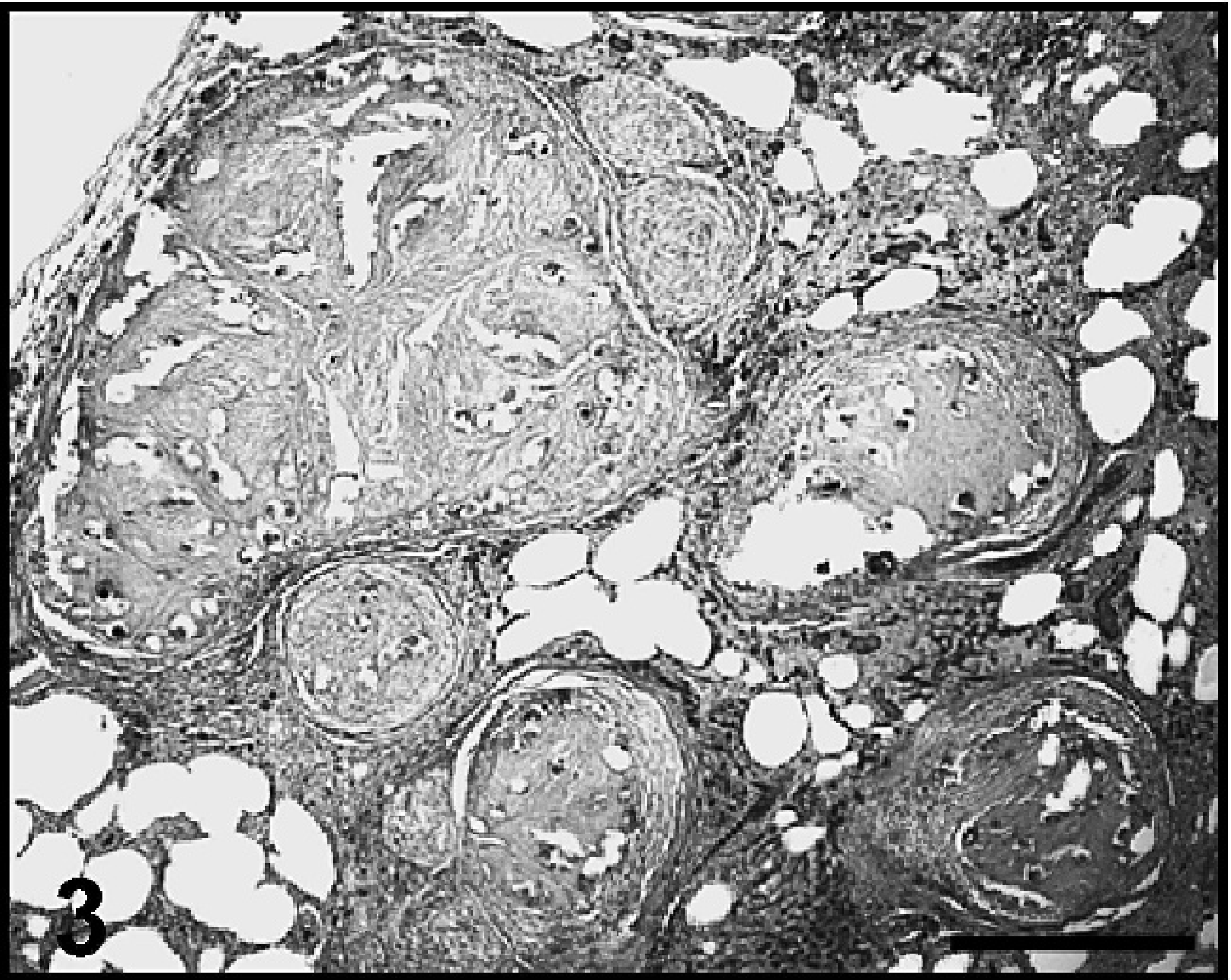
Lung, dog 4. Multifocal-to-coalescing caseogranulomas replace and distort areas of the pulmonary parenchyma and are most prevalent in apical lung lobes. Hematoxylin and eosin. Bar = 500 μm.
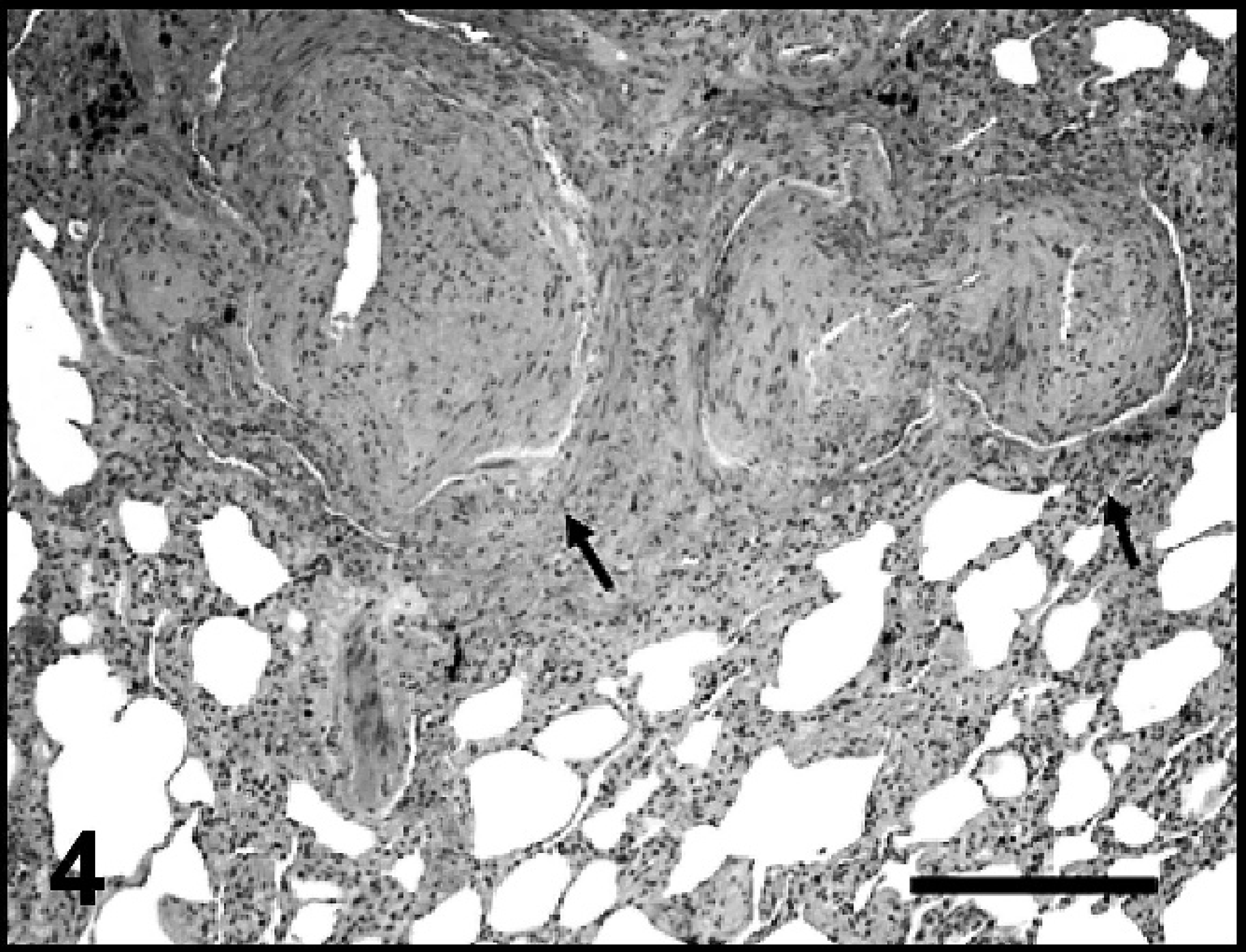
Lung, dog 6. The photomicrograph depicts the prominent interstitial fibrosis noted in this dog. Also shown are 2 large, thickened pulmonary arteries (arrows) with hypertrophied smooth muscle walls and narrowed lumina. Hematoxylin and eosin. Bar = 500 μm.
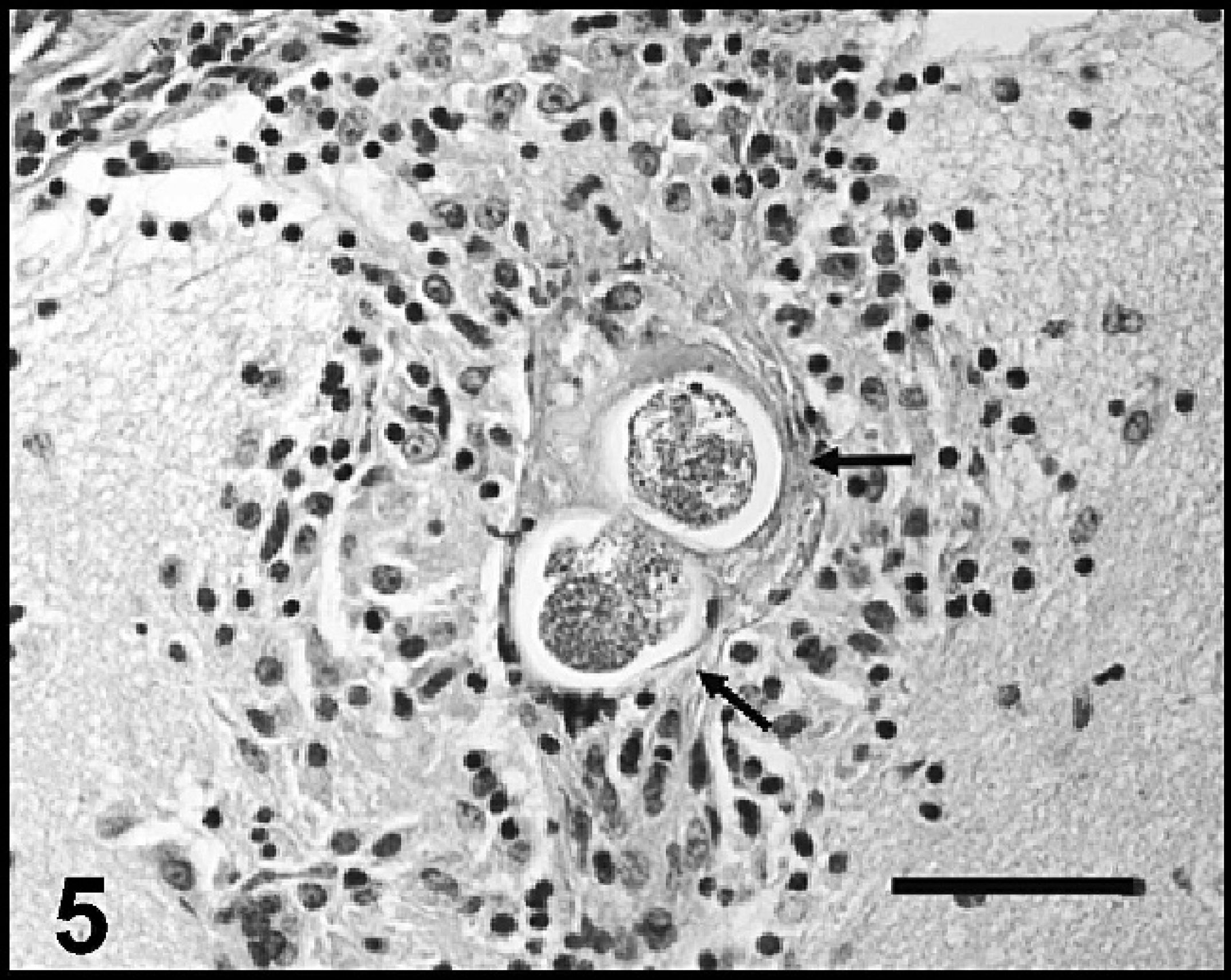
Brain: cerebrum, dog 4. A discrete cluster of epithelioid macrophages, lymphocytes, and plasma cells surrounds 2 nematode eggs within the neuropil subjacent to the leptomeninges in the cerebral cortex. Hematoxylin and eosin. Bar = 50 μm.
Lung, dog 7. The pulmonary interstitium is markedly diffusely expanded as a result of inflammatory infiltrates and marked vascular and interstitial smooth muscle hyperplasia/hypertrophy. Arrows point to several prominent pulmonary arteries with thickened tunica media and narrowed lumina. Hematoxylin and eosin (HE). Bar = 500 μm.
There were rare, mild, multifocal interstitial infiltrates of macrophages, lymphocytes, and plasma cells scattered within the renal cortices of dogs 2, 4, and 6. Nematode eggs were rarely embedded within these inflammatory foci in dog 4.
Mild, small, multifocal, discrete clusters of macrophages, lymphocytes, plasma cells, and rare MNGCs surrounded small blood vessels within the leptomeninges covering the cerebrum or were located within the cerebral neuropil of dogs 3, 4, and 6. In dog 4, one such focus within the cerebrum surrounded 2 degenerate nematode eggs (Fig. 5).
Dog 7 was a castrated male, 3.5-year-old, Dalmatian. The dog first presented to a veterinary clinic in St. John's, Newfoundland, with a complaint of persistent coughing. Fecal Baermann testing 15 days after presentation revealed numerous A. vasorum larvae. On day 16 postpresentation, treatment for infection was initiated with a single dose of milbemycin oxime, which was repeated 7 days later (day 23 postpresentation). Three days later, the dog suddenly developed severe intractable seizures that worsened over the next 24 hours. Serum biochemistry, complete blood counts, and urinalysis results were normal. Seizure activity could not be controlled with medical therapy, and the dog was euthanized the next day.
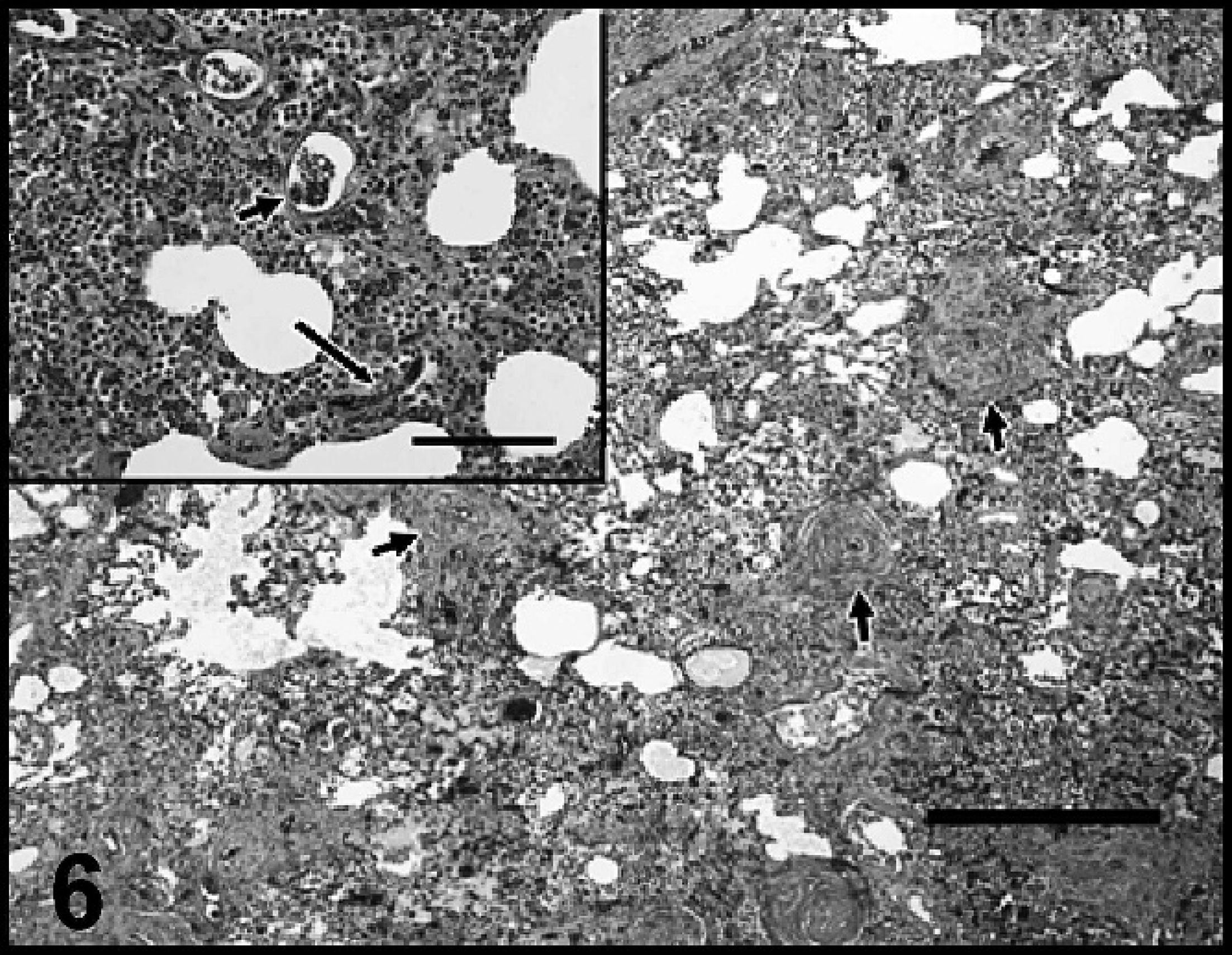
Veterinary clinics within the St. John's area had been notified of this study and that the authors would be interested in performing postmortem evaluations on any euthanized, potentially A. vasorum-infected dogs during collection periods. The owners agreed to include this dog in the present study. At postmortem, 486 intact adult A. vasorum were recovered from pulmonary arteries. In addition, worm fragments indicating the presence of an additional 86 worms were recovered for a total A. vasorum-worm burden estimate of 572. Approximately one third of the intact worms showed signs of activity/movement when placed in warm (37°C) isotonic saline. Numerous L1 were recovered from rectal feces by Baermann examination. The lungs were mottled red and pink and variably rubbery to firm. In all lung sections examined, lesions were severe, widespread, and largely centered on vascular structures. There was marked smooth muscle hyperplasia/hypertrophy of the walls of small, medium, and large pulmonary arteries, often resulting in narrowing of the vessel lumina (Fig. 6). In large arteries, adult nematodes, eggs, and larvae were embedded in partially organized fibrin, which partially to completely occluded several vessel lumina. Many thrombosed pulmonary arteries were recanalized. Less frequently, smaller arteries contained fibrin thrombi. This dog had a severe, diffuse, interstitial pyogranulomatous and suppurative pneumonia with myriad nematode eggs and larvae accompanied by frequent, small, patchy areas of alveolar hemorrhage and clusters of hemosiderin-laden macrophages (Fig. 6, inset). A caseogranuloma similar to those described, which contained numerous degenerate nematode larvae and eggs partially surrounded by dense, fibrovascular connective tissue, was located at the margin of a caudal lung lobe.
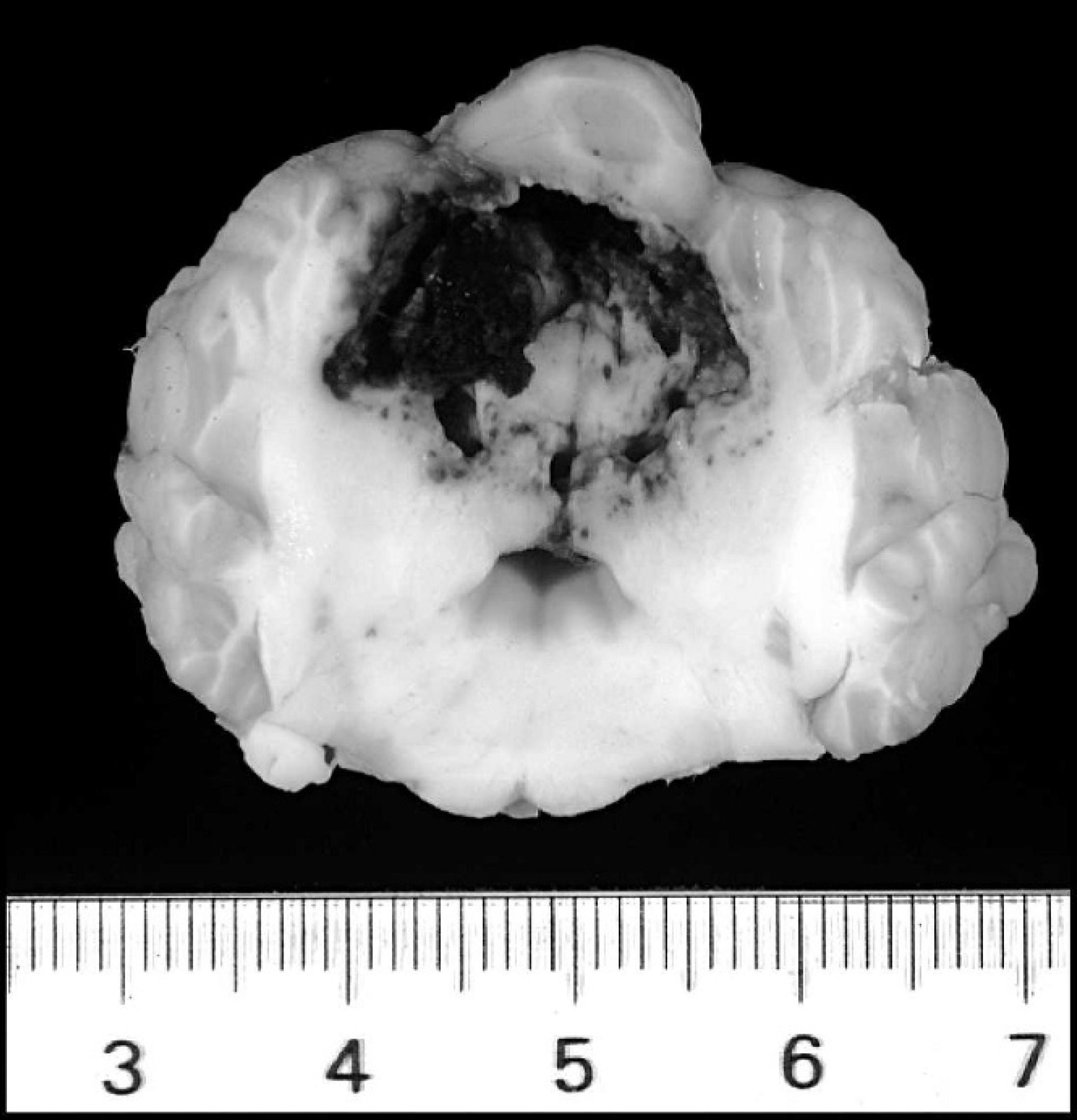
Brain: cerebellum, dog 7. A 2.5-cm-in-greatest-diameter area of hemorrhage obliterates a large portion of the cerebellum (scale is in centimeters).
In sections of tracheobronchial lymph nodes, multifocal, large areas of the cortex were obliterated by many nematode larvae associated with dense, granulomatous, and eosinophilic inflammatory infiltrates. In most sections, medullary sinuses were distended with many hemosiderin-laden macrophages.
Two discrete, dark red foci of hemorrhage and malacia, measuring 1–2 cm in greatest diameter and involving both the gray and white matter, were present within the cerebrum. One such focus was located in the left frontal lobe, while the other was present in the left temporal lobe. An additional 2.5-cm-in-greatest-diameter focus of hemorrhage partially effaced the vermis of the cerebellum (Fig. 7). Microscopically, the brain tissue in these areas was obliterated by hemorrhage. The surrounding neuropil was variably edematous and often contained smaller foci of hemorrhage, scattered swollen axons, and mild infiltrates of neutrophils. Blood vessels in the adjacent tissue were lined by plump reactive endothelium and were often cuffed by small numbers of neutrophils, lymphocytes, and plasma cells. Scattered throughout cerebral and cerebellar sections were small discrete aggregates of macrophages, lymphocytes, plasma cells, and occasional MNGCs often centered on portions of nematode larvae or, less frequently, nematode eggs.
The renal cortices contained multifocal, moderate, lymphoplasmacytic, interstitial infiltrates, and rare small foci of mild fibrosis. Glomerular, and less frequently, interstitial capillaries contained nematode larvae.
Rare nematodes eggs were present within the red pulp and trabecular blood vessels in the spleen. Small clusters of macrophages, lymphocytes, and plasma cells surrounded nematode larvae in the adrenal cortex.
Discussion
Canine angiostrongylosis in Eastern Newfoundland has been increasingly diagnosed by veterinary practitioners since the early 1990s and has become a major concern for dog owners and clinicians. Angiostrongylus vasorum infection in dogs is typically asymptomatic or associated with mild, intermittent clinical signs in the early stages of infection. In later stages of infection or when parasite loads are high, the most common clinical signs reported in affected dogs are coughing, dyspnea, and exercise intolerance, which are generally caused by chronic interstitial pneumonia with variable fibrosis. However, presenting clinical signs can be quite variable and include anorexia, stunted growth, weight loss, weakness, depression, vomiting, collapse, lameness, subcutaneous swelling, bleeding disorders, hypercalcemia, disseminated intravascular coagulation, and neurological signs. 1,4,8,11,12,16,21,24,27,35,36
Unfortunately, in the present study the authors were unable to assess the dogs antemortem, and clinical histories in 6 of 7 infected dogs were not available. Thus, the severity of clinical signs resulting from the pathological changes observed could not be correlated to clinical disease. However, it is speculated that, in at least 2 of these dogs (dogs 1 and 5), where lesions were very mild, small in number and size, and with few parasites found in pulmonary arteries, infection was probably subclinical. In dog 6, pulmonary lesions were widespread, chronic, and accompanied by macroscopic and microscopic evidence of right heart failure. This has been described previously and, along with progressive pulmonary disease and respiratory failure, is a common cause of death in chronically infected, untreated dogs. 4,20
Reference values for heart weights as a percentage of body weight in normal mature dogs at necropsy have been reported as 0.71% (range: 0.43–0.99%). 25 In the present study, heart weights ranged from 0.52–0.97% body weight (thus all were within normal range). Normally, there is an approximately 1:3 ratio in thickness between the right and left ventricular free walls. In dogs 4 and 7 (dogs with severe lung lesions), this ratio was 1:2.5, suggesting mild thickening of the right ventricle, which likely represented mild compensatory hypertrophy secondary to pulmonary hypertension; clinical evidence of right heart failure was not evident at the time of death. In dog 6 (also having severe lung lesions and evidence of right heart failure), the thickness ratio between the right and left ventricle was 1:1.4, consistent with marked thickening of the right ventricle free wall.
The pulmonary lesions described in the present report are similar to those previously described in both naturally and experimentally A. vasorum- infected dogs and foxes. 4,7,23 The most consistent finding in infected dogs was interstitial pneumonia with prominent vascular changes. Pneumonia, in these cases, was generally granulomatous with variable amounts of suppurative and eosinophilic inflammation. Interstitial fibrosis was common and variable in degree. Pulmonary arteries were plugged with fibrin and sometimes parasites or, more commonly, were occluded with dense fibrous tissue or had partially recanalized. In dogs with severe lung lesions (dogs 3, 4, 6, and 7), smooth muscle hypertrophy and hyperplasia of the tunica media of pulmonary arteries was prominent and indicative of pulmonary hypertension secondary to increased resistance to blood flow in altered blood vessels and the interstitium. The presence of parasites was quite variable. When present in the lung, nematode larvae and eggs were found within arteries, smaller capillaries, the interstitium, and alveoli, and were associated with inflammatory infiltrates, while adult worms were restricted to pulmonary arteries. The formation of caseogranulomas, primarily involving the lung lobe margins and subpleural sites, was common and likely represented areas of tissue infarction.
Small granulomatous foci of inflammation sometimes associated with nematode eggs and larvae were found in various organs in dogs with moderate-to-severe lung lesions. These sites included kidney, brain, spleen, tracheobronchial lymph node, and adrenal gland. In addition to sites just mentioned, extrapulmonary A. vasorum parasites have been reported in the skin, liver, pancreas, peripheral blood, cerebrospinal fluid, pericardial sac, urinary bladder, femoral artery, intestinal tract, thyroid gland, pituitary gland, skeletal muscle, heart, and eye. 10,19,21 Parasite eggs and larvae are presumably systemically disseminated via hematogenous routes. Larvae, eggs, and, less frequently, adults may gain access to systemic circulation through the damaged lung, resulting in parasitic emboli to various sites. Aberrant larval migration from the gastrointestinal tract may also result in parasites in extrapulmonary sites where they mature into adult worms. The granulomatous inflammation and tissue damage associated with parasite dissemination can potentially result in multiorgan damage and, when severe, possibly lead to impaired organ function.
Dog 7 developed severe seizures that were unresponsive to treatment necessitating euthanasia and which began shortly after the initiation of treatment for A. vasorum infection. At necropsy, multifocal, large areas of acute hemorrhage obliterated regions of the cerebrum and cerebellum. It is impossible to say if treatment, which included the killing of large numbers of intravascular worms in this dog, was a factor in the development of this lesion. The most common anthelmintics used to treat canine angiostrongylosis are fenbendazole, mebendazole, ivermectin, levamisole, and milbemycin oxime. 4,9 Studies have not definitively shown whether any of these medications completely clear infection. Complications following treatment are uncommon and usually consist of dyspnea and vomiting. Hypovolemic shock, presumably caused by an anaphylactic reaction to the massive release of worm antigen following the rapid death of adult worms, has been reported after levamisole treatment. 30 To further complicate things, intracranial (and intraspinal hemorrhage) has been reported in untreated infected dogs. 12,35
Bleeding tendencies caused by acquired von Willebrand factor deficiency and immune-mediated throm-bocytopenia have been reported in dogs in association with A. vasorum infection. 13,36 In the latter case, increased monocyte-macrophage activation and platelet consumption caused by the formation of antiplatelet antibodies secondary to A. vasorum infection was proposed as the cause of thrombocytopenia. More commonly, the cause of abnormal bleeding in A. vasorum-infected dogs is postulated to be an intravascular consumptive coagulopathy caused by the chronic formation of intravascular fibrin thrombi in the lung, resulting in the consumption of clotting factors and platelets. The presence of intravascular parasites has been thought to activate the intrinsic coagulation pathway by direct physical damage to endothelium, by the deposition of immune complexes, immunoglobulins, and/or complement within the tunica intima and via complement-mediated damage to vascular endothelium resulting in exposure of subendothelial collagen. The extrinsic coagulation pathway may also be activated via the release of tissue factor (or thromboplastin) from damaged tissues secondary to chronic inflammation and areas of tissue infarction. 7 It is also possible that the parasite secretes substances that promote intravascular coagulation. Commonly reported abnormalities in affected dogs include thrombocytopenia, prolonged activated partial thromboplastin time, and prothrombin time. Occasionally anemia secondary to blood loss has been reported. Elevations in fibrin degradation products in association with prolonged clotting times and thrombocytopenia is not uncommon and is indicative of disseminated intravascular coagulation. 24 In most cases, these acquired coagulopathies resolve once treatment for angiostrongylosis is initiated. In addition to intracranial hemorrhage, neurological signs in infected dogs could potentially be caused by embolism of fibrin thrombi or parasites and/or tissue infarction within the central nervous system or hypoxia secondary to chronic heart failure and respiratory disease.
It is interesting to note that in the present study, the number of worms recovered from the lungs did not always correspond to the severity of pathological findings. In dogs with relatively mild infections and rare lesions, it was necessary to dissect quite far along pulmonary arteries toward the margins of lung lobes to find parasites. In dog 6, despite the fact that pulmonary lesions were severe and widespread, relatively few worms were found. In the latter case, it is possible that the severe, diffuse, chronic pulmonary inflammation and fibrosis resulted in an inhospitable local environment incapable of supporting parasites, leading to die-off of the organisms. Alternatively, it is possible that attempts were made to treat this dog for angiostrongylosis in the past or that this dog was able to mount a limited immune response, which controlled parasite numbers but was unable to eradicate infection. In both scenarios, it is possible that intrapulmonary worm numbers were kept low but infection was not cleared, resulting in chronic, progressive lung damage.
Dog 7 had the largest worm burden and severe, diffuse, chronic, pyogranulomatous interstitial pneumonia. Thrombosis, vascular, and interstitial smooth muscle hypertrophy/hyperplasia with areas of tissue infarction and hemorrhage were prominent findings. Pulmonary fibrosis was relatively mild. This suggested a more acute and active stage of infection and a much heavier parasite load caused by ingestion of large numbers of infective larvae in a relatively short time period. This was the only dog in which the authors were able to obtain a clinical history, and interestingly the owners report that this dog was known to eat slugs frequently.
Sudden death in dogs caused by angiostrongylosis, thought to be associated with large worm burdens in relatively young dogs, has been reported. 4,24 Little is known concerning the magnitude of A. vasorum infection required to cause sudden death, making it difficult to place the worm burden observed in this dog (>500 worms) in proper context. A recent reported cited the recovery of 210 worms from a dog that died acutely caused by angiostrongylosis. 19 It is likely that the number of nematodes recovered from dog 7 represents an extremely high parasite burden.
Antemortem diagnosis of A. vasorum infection is generally confirmed by identifying L1 via fecal examination using the Baermann technique. Larvae may be found in bronchoalveolar lavage (BAL) samples, although less frequently. Larvae are shed intermittently, so a negative fecal test or BAL sample does not rule out A. vasorum infection. In this group of 7 infected dogs, 3 had positive Baermann fecal tests. Often multiple fecal tests are required to make a diagnosis. In many cases, by the time dogs are exhibiting clinical signs of infection, chronic and often irreversible lung damage has occurred that can impair organ function and potentially decrease the life span of affected individuals. 4 The development of a reliable, commercially available screening test to detect early or subclinical infections, such as is available for Dirofilaria immitis, would significantly improve the prognosis for dogs with angiostrongylosis. Circulating antigen and antibody has been detected in dogs naturally infected with A. vasorum by the use of a serum enzyme-linked immunosorbent assay and Western blot analysis, respectively. 34
In Europe, A. vasorum infection has been recognized as a common parasite in discrete, well-defined areas, with sporadic cases occurring elsewhere. In recent years, A. vasorum has become established in several new geographical areas, including new areas in Scandinavia and this North American focus in Newfoundland, Canada. 5,17 Many of the same intermediate and reservoir hosts and wet climatic conditions present in Newfoundland are located in the nearby Maritime Provinces of Canada and elsewhere. A 2004 study 17 postulated that local changes in climate that favor the proliferation of snails and slugs and an abundance of reservoir hosts would favor the expansion of this parasite beyond its current geographical range. Increased travel of pet dogs worldwide could facilitate the introduction of A. vasorum from endemic areas to new sites. The potential for other wildlife species, such as coyotes, to act as reservoir hosts, also cannot be ignored. All these factors favor the eventual spread of A. vasorum to the nearby Canadian Maritime Provinces and potentially elsewhere in North America.
Acknowledgements
The authors heartily thank the St. John's Newfoundland branches of the SPCA and Humane Societies for allowing the examinations of these dogs. The value of these animals in ongoing research may help to alleviate suffering in the living. The authors thank Rebecca Jeffery, Dr. Tracey Matthews, Cathy Roberts, and Nicole Murphy for their essential input and assistance in this endeavor, the staff of the Animal Health Division of the Department of Natural Resources for their hospitality, and Dr. David Sims for his assistance with digital imaging. This study was funded through the Animal Welfare Unit of the Atlantic Veterinary College, which enjoys the generous support of the Sir James Dunn Foundation.