
Abstract
Base excision repair, which is initiated by the DNA N-glycosylase proteins, is the frontline for repairing potentially mutagenic DNA base damage. Several base excision repair genes are deregulated in cancer and affect cellular outcomes to chemotherapy and carcinogenesis. Endonuclease VIII-like 3 (NEIL3) is a DNA glycosylase protein that is involved in oxidative and interstrand crosslink DNA damage repair. Our previous work has showed that NEIL3 is required to maintain replication fork integrity. It is unknown whether NEIL3 overexpression could contribute to cancer phenotypes, and its prognostic value and use as potential drug target remain unexplored. Our analysis of cancer genomics data sets reveals that NEIL3 frequently undergoes overexpression in several cancers. Furthermore, patients who exhibited NEIL3 overexpression with pancreatic adenocarcinoma, lung adenocarcinoma, lower grade glioma, kidney renal clear cell carcinoma, and kidney papillary cell carcinoma had worse overall survival. Importantly, NEIL3 overexpressed tumors accumulate mutation and chromosomal variations. Furthermore, NEIL3 overexpressed tumors exhibit simultaneous overexpression of homologous recombination genes (BRCA1/2) and mismatch repair genes (MSH2/MSH6). However, NEIL3 overexpression is negatively correlated with tumor overexpressing nucleotide excision repair genes (XPA, XPC, ERCC1/2). Our results suggest that NEIL3 might be a potential prognosis marker for high-risk patients, and/or an attractive therapeutic target for selected cancers.
Background
DNA base damage induced by oxidative and alkylating agents is removed by base excision repair (BER). 1 BER is initiated by a DNA glycosylase that excises damaged bases and is subsequently processed by apurinic/apyrimidinic endonuclease I (APE1). 2 This leads to a DNA gap, which is then filled in by DNA polymerase β (Pol β) and DNA ligase III (LIG3). 3 The DNA glycosylase not only recognizes and removes the damaged base, but also protects cells from mutagenic and/or cytotoxic effects of DNA base lesions while still maintaining genomic integrity.4,5 Several studies have provided multiple lines of evidence that identify the role of DNA glycosylases in protecting humans from different disease including neuro-degeneration, autoimmunity, and cancer.6–8 Aberrant expression of DNA glycosylase creates an imbalance in the BER pathway, which leads to saturation of downstream repair enzymes and/or accumulation of genotoxic BER intermediates.2,9,10
Recent advances in high-throughput sequencing technology have revealed how specific gene abnormalities contribute to somatic mutation load and chromosomal variation, as reported in The Cancer Genome Atlas (TCGA). It is known that abnormal regulation or mutation in DNA repair genes can increase the risk of cancer development and impact treatment response. 11 Numerous DNA repair gene expressions influence DNA replication efficiency and have been associated with different stages of cancer including metastasis.12,13 In particular, BER expression is deregulated in several types of cancer, which leads to genomic instability, ultimately impacting treatment outcomes.14–17 To date, four mammalian DNA glycosylases that recognize and excise oxidized bases have been thoroughly characterized: OGG1, NTH1, NEIL1, and NEIL2.8,18,19 Several DNA glycosylases are involved in repair of post-replication DNA lesions including repair of oxidation DNA damage and are localized at nascent DNA structures associated with DNA replication.20,21 Mammalian endonuclease VIII-like 3 (NEIL3) DNA glycosylase removes the further oxidized DNA damage products of 8-oxo-7, 8-dihydroguanine, spiroiminodihydantoin, and guanidinohydantoin from ssDNA substrates.22–25 The catalytic domain of NEIL3 contains a DNA binding pocket that can accommodate a broad spectrum of damage. 26 NEIL3 excises the oxidized DNA base from telomere G-quadruplexes, DNA structures that block DNA processing enzymes.27,28 Furthermore, NEIL3 cleaves 5′ hydroxyl cytosine (5′-HOC) and thymine glycol (Tg) DNA lesions from double stranded DNA. 29 NEIL3 is highly expressed in various human cancer cells and is associated in a few cases with metastatic cancer, suggesting that it may be required for the maintenance of cancer cell growth or malignancy progression.30,31
NEIL3 is expressed in proliferating cells during S/G2 phase and has been reported to be a cell cycle regulated gene, suggesting that it is needed to maintain replication fork stability during replication stress.32–34 NEIL3 is the DNA glycosylase that is associated with replisomes, suggesting it is mainly acting close to the replication fork. 20 In support of this observation, cells deficient in NEIL3 accumulate spontaneous DNA damage associated with replication forks, suggesting that NEIL3 has a non-catalytic role in maintaining genomic integrity during DNA replication. 32
The up-regulation of NEIL3 expression in diverse cancer types is consistent with the results from previously published data, 30 but the effect of such up-regulation on clinical outcomes has not yet been determined. In this study, using TCGA data, we show that cancer patients with tumors have higher NEIL3 expression and a significantly lower overall survival (OS) rate compared to patients whose tumors express lower levels of NEIL3. Furthermore, NEIL3 overexpressing tumors harbor greater genomic instability that is clearly evidenced by significantly higher levels of total mutation counts and chromosomal alteration. Finally, our study showed a positive correlation between NEIL3 overexpression and homologous recombination (HR) and mismatch repair (MMR) gene expression, but not nucleotide excision repair (NER). While the biological relevance of this correlation is unclear, it raises the interesting possibility that this particular profile would help explain the worse prognosis seen in NEIL3 overexpressing cancer patients. High levels of NEIL3 expression could contribute to cancer phenotype via increase genomic instability and/or interfering in alternative DNA repair. These findings have clinical implications for cancer patients with NER gene (XPA/XPC/ERCC1) deficiency for whom NEIL3 should be explored as a potential drug target. Our results suggest that NEIL3 could serve as a potential prognostic marker for selection of patients for chemotherapy or could be used as a therapeutic target for cancer.
Methods
Data acquisition
TCGA provisional data sets were pulled from cBioProtal (http://cbioportal.org) between the months of November 2017 and January 2018. The pulled data sets consisted of: acute myeloid leukemia (AML; n = 200), bladder urothelial carcinoma (BUC; n = 413), brain lower grade glioma (LGG; n = 530), breast invasive carcinoma (BC; n = 1105), cervical squamous cell carcinoma and endocervical adenocarcinoma (n = 309), colorectal adenocarcinoma (CADC; n = 633), esophageal carcinoma (EC; n = 186), glioblastoma multiform (GBM; n = 604), head and neck squamous cell carcinoma (HNSCC; n = 530), kidney renal clear cell carcinoma (KRCCC; n = 538), kidney renal papillary cell carcinoma (KPCC; n = 293), lung adenocarcinoma (LUAC; n = 522), lung squamous cell carcinoma (LSCC; n = 504), ovarian serous cystadenocarcinoma (OSCAC; n = 606), pancreatic adenocarcinoma (PADC; n = 186), prostate adenocarcinoma (PC; n = 499), skin cutaneous melanoma (SCM; n = 478), stomach adenocarcinoma (SADC; n = 478), testicular germ cell cancer (TGCC; n = 156), thyroid carcinoma (TC; n = 516), and uterine corpus endometrial carcinoma (UCEC; n = 548). Copy number altered genome (CNA) data were pulled from each specific cancer’s Study Summary page on cBioPortal.
Exclusion and inclusion criteria
To limit the total number of cancer types to be examined, any cancer type with an overall rate of survival of 90% after 100 months post-onset was excluded from further analysis. This resulted in the removal of PC, TGCC, and TC from further analysis. Since NEIL3 is the primary source of interest for this analysis, only individuals with valid RNA Seq V2 RSEM data for NEIL3 were included, reducing the sample sizes of the remaining cancer types. To group individuals as having either low or high expression of NEIL3, the z scores of each individual were considered. Individuals with a z score of ≤−0.5 were placed into the low expressing group while individuals with a z score of ≥0.5 were placed into the high expressing group. Individuals falling between −0.5 and 0.5 were excluded from analyses. In addition, individuals in the low or high expressing group reporting a null value for OS months were excluded from the group analyses, and individuals in the low or high expressing group reporting a null value for OS months were excluded from the group analyses.
Mutation count and CNA
The mutation counts for each individual were pulled from the TCGA data set for each selected individual. Values were grouped according to low and high NEIL3 expression for a given cancer type and an unpaired t test was used to assess the statistical differences between the two groups. The CNA data for each individual were pulled from the cBioPortal website and merged with the TCGA data set of a given cancer line. For correlation pairing, we used correlation analysis with Pearson correlation coefficient to determine the association between NEIL3 and HR repair gene expression of BRCA1, BRCA2, MMR (MSH2/MSH6), NER (XPA/XPC/ERCC1/2).
Statistical analysis
Group comparisons for continuous data were conducted using t tests or χ2 tests, and quantitative variables were analyzed with the Wilcoxon signed-rank test or the Spearman rank correlation test. Kaplan–Meier analysis was used to assess OS. Uni-variable associations between NEIL3 overexpression and clinicopathologic variables were tested using nonparametric tests. Linear regression analyses to evaluate the correlation between DNA repair pathways were performed and goodness fit measure using R2. The results of the cell function experiments were presented as the mean values ± SEM. Statistical significance was set at P < 0.05.
Results
Patient data analysis and clinical characteristics of NEIL3 overexpressing tumors
Data were accessed from cBioPortal (www.cBioPortal.org). TCGA provisional RNA-seq data and corresponding clinic information for 18 cancer types were retrieved for validation. All cancer data extracted from TCGA provisional data sets were analyzed based upon NEIL3 expression levels for 18 different cancers with 8662 cancer patients. Complete data of eight different cancers were selected for 4282 cancer patients and further refined to 3474 cases with complete information related to NEIL3 expression and the patients’ clinical information. Overall, we have found that 23.4% of primary tumors exhibit overexpressed NEIL3 (Table 1). The percentage of tumors with NEIL3 overexpression ranges between 13% and 89%. Remarkably high overexpression was noted in GBM tumors, followed with BC (27%), PADC (26%), LUAC (15%), KPCC (15%), KRCCC (15%), and LGG tumors (14%). We investigated whether the difference in NEIL3 overexpression among these tumors was associated with any pathological factors in the PADC, LUAC, KPCC, and KRCCC patients. The results showed that high NEIL3 expression levels were significantly associated with a positive lymph node status in PADC (****P < 0.0001), and higher T3 and T4 stages of tumor in PADC (81%, ****P < 0.0001), KPCC (61%; **P < 0.01), and KRCCC (60%; **P < 0.01) as shown in Table 2. In contrast, as shown in Table 2, NEIL3 overexpression is significantly observed at T1/T2 tumor stage in LUAC (78%; P < 0.0001) and BC (86%; P < 0.0001).
NEIL3 is overexpressed in different types of cancers.

All patient RNA-seq data extracted from TCGA provisional data sets. Percent of overexpression of NEIL3 calculated by dividing the number of patients with NEIL3 expression z score > 0.05 by total number of patients.
NEIL3 overexpression is associated with different stages of tumor progression.

Statistical significance calculated using pair wise comparison of the numbers of positive samples in tumor stage 1/2 versus 3/4.
NEIL3 overexpression leads to poor OS in selected types of cancer
To investigate whether the expression profile of the NEIL3 in tumors could be related to patient survival, Kaplan–Meir survival curves were generated based upon the designation of individuals as either low expressing NEIL3 (z score <0.5) or high expressing NEIL3 (z-score >0.5) to assess the impact of survival for each cancer type. We have found that NEIL3 overexpression significantly and negatively impacts the OS of patients with PADC (**P < 0.01, Figure 1(a)), LUAC (*P < 0.05, Figure 1(b)), LGG (****P < 0.001, Figure 1(c)), KRCCC (Figure 1(d), ****P < 0.001), and KPCC (Figure 1(e), ****P < 0.001). There was no statistically significant difference in invasive breast cancer among high versus low NEIL3 overexpressing groups (data not shown). However, we have found that NEIL3 overexpression significantly decreases the OS of triple negative breast cancer (TNBC) patients versus non-triple negative breast cancer (Figure 1(f), ****P < 0.0001). Our data show that as NEIL3 mRNA levels increased, the outcomes for the PADC, LUAC, KRCCC, and KPCC cancer patients became worse, and the relative risk of death increased (Figure 1). No significant difference was observed in OS of other types of cancer patients (AML, BUC, cutaneous squamous-cell carcinoma (CSCC), CADC, EC, subungual squamous cell carcinoma (SSCC), OSCA, SCM, and UCEC). However, overexpression of NEIL3 enhanced good prognosis in colorectal and stomach cancer patients, suggesting that NEIL3 may have a specific function related to tumor types (Supplement Figure 1).

Overexpression of NEIL3 expression unfavorably affected the overall survival of patients in different types of cancer: (a) PADC, (b) LUAC, (c) LGG, (d) KRCCC, (e) KPCC, and (f) BC.
NEIL3 overexpression significantly increases genomic alteration in cancer patients
To assess deregulation of NEIL3 in human PADC, LUAC, LGG, BC, KRCCC, and KPCC associated with genomic alteration. First, we analyzed whether NEIL3 overexpression was associated with mutation load in tumors in patients with that showed poor OS. The number of mutation counts significantly increase in NEIL3 overexpressing PADC, LUAC, BC, and LGG tumors rather than lower expressing tumors (Figure 2(a), P < 0.0001). In addition, we have found that other cancers that exhibit overexpressed NEIL3 (LSCC, HNSC, and SCM) have significantly increased tumor mutation loads with no impact on patient OS (Supplement Figure 2). Furthermore, NEIL3 overexpression may enhance chromosomal rearrangement, and we found that patients overexpressing NEIL3 accumulate a fraction of the copy number variation (CNV) in PADC, LUAC, BC, and LGG patients versus low-expressing groups (Figure 2(b), P < 0.001). In contrast, tumors with NEIL3 overexpression do not increase or impact the fraction of chromosomal variation or mutation counts in KRCCC and KPCC (Figure 2(a) and (b)). Interestingly, all PADC tumors harbor 100% Kirsten rat sarcoma viral oncogene homolog (Kras) mutation and 64% TP53 mutation, suggesting that there is a strong association of cancer driving genes (data not shown).
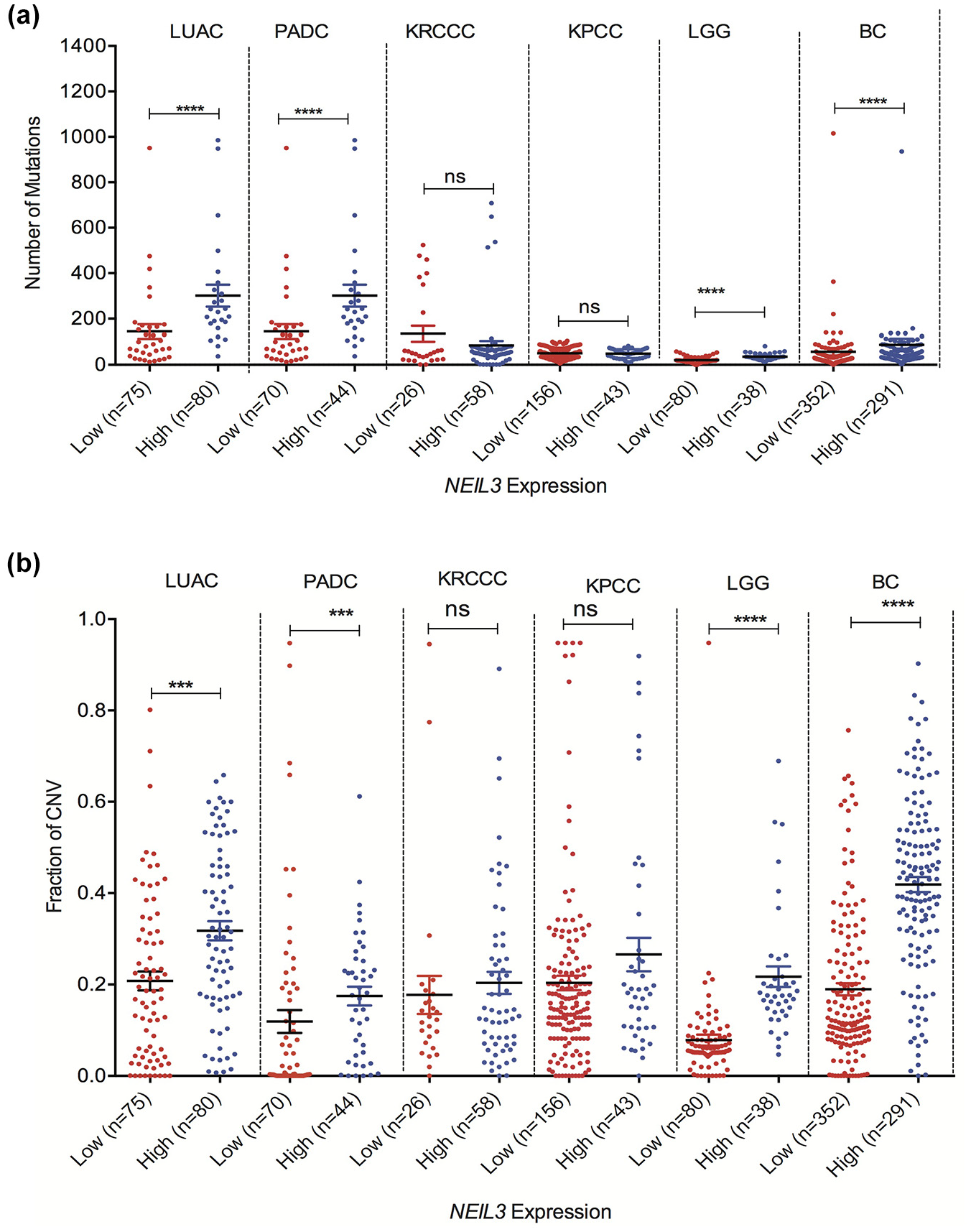
Fraction of genome altered and mutational load increases in patents with high NEIL3 overexpressing tumors. (a) Samples from TCGA database were curated and we compared the magnitude of copy number variation between high NEIL3 expressing tumors versus low NEIL3 expressing tumors. (b) Mutational load between NEIL3 overexpressing tumors versus low expressing tumors.
High NEIL3 expressed tumors are negatively correlated with expression of nucleotide excision repair genes and are positively correlated with expression of MMR and HR DNA repair genes
Recently, an alternative interstrand crosslink (ICL) repair pathway that depends on the NEIL3 DNA glycosylase has been discovered 35 suggesting that NEIL3 is required for replication fork convergence at an ICL. Interestingly, our data show that NEIL3 overexpressing tumors are negatively correlated with the expression of NER genes (XPA, XPC, and ERCC1/2) suggesting a backup mechanism of platinum-based ICL DNA lesions (Figure 3(c), (f), (i), and (l)). Furthermore, our in-silico data analysis shows a positive correlation between the NEIL3 overexpressing tumors with over expression of homologous recombination proteins (BRCA1/2; Figure 3(a), (d), (g), and (j)) and MMR gene expression (MSH2/MSH6; Figure 3(b), (e), (h), and (i)) suggesting that the potential DNA damage tolerance to genotoxic chemotherapy agents. Furthermore, NEIL3 overexpressed tumors harbor mutation of the most common tumor driver genes including Kras, TP53, epidermal growth factor receptor (EGFR) genes that likely contribute to promote tumor initiation via spontaneous genomic instability.
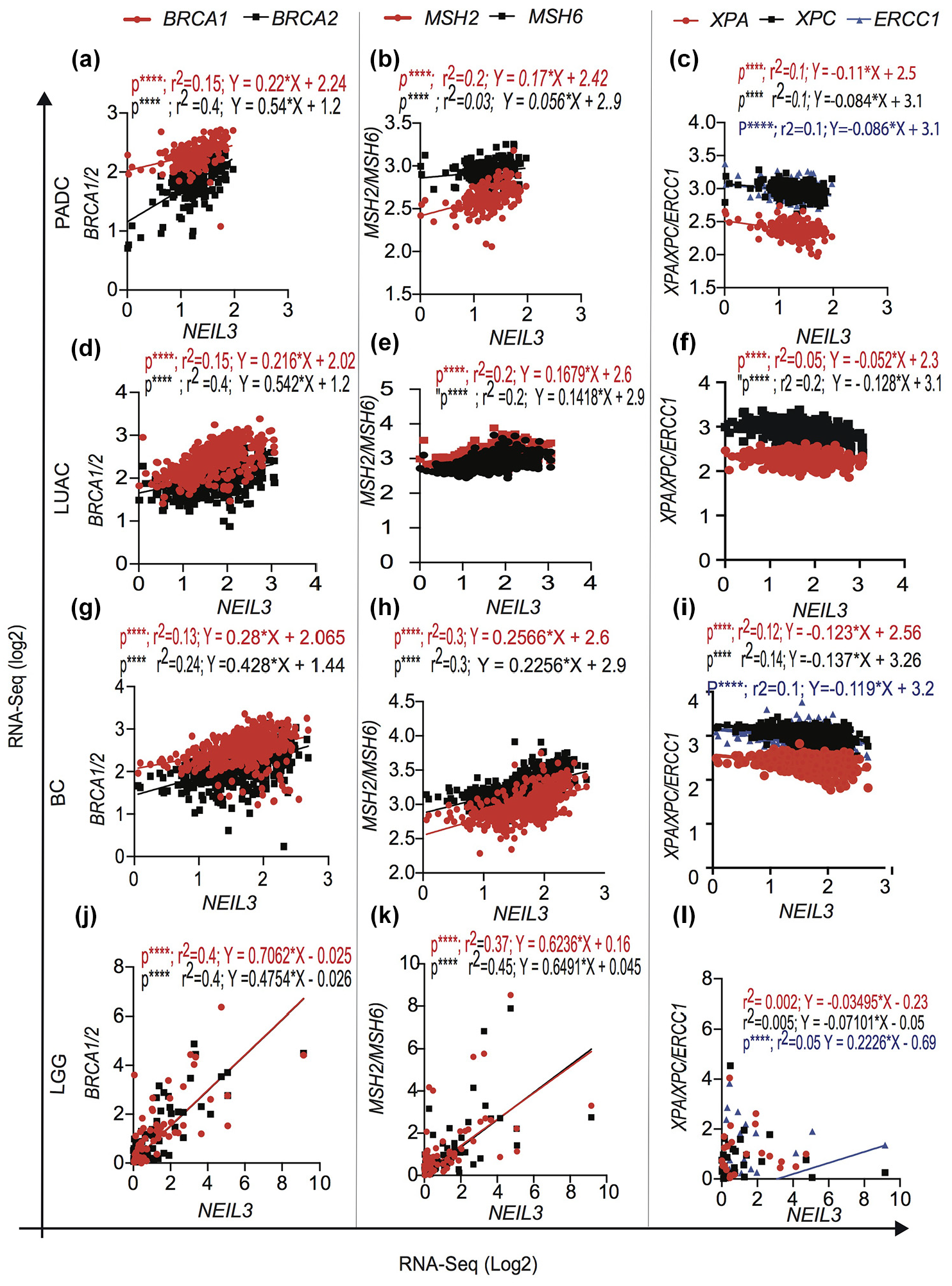
Association of NEIL3 overexpression with homologous recombination repair, mismatch repair and nucleotide excision repair genes transcript. Different types of cancer overexpressing NEIL3 positively correlated with BRCA1/2, MSH2/MSH6 and negatively correlated in XPA/XPC/ERCC1 in: (a)–(c) PADC; (d)–(f) LUAC; (g)–(i) BC; (j)–(l) LGG.
Discussion
In this work, we report that NEIL3 is overexpressed in a majority of human cancers including GBM, LGG, PADC, LUAC, KRCCC, and KPCC (Table 1). The frequent overexpression of NEIL3 in a wide range of tumors suggests that this event is unlikely to be related to a particular carcinogen and may play a more general role favoring high rates of proliferation and growth of tumor cells in selected types of cancer (GBM, LGG, PADC, LUAC, KRCCC, and KPCC). Our most important discovery is that NEIL3 overexpression is significantly associated with poor clinical outcomes in patients suffering from six different types of cancer (GBM, LGG, PADC, LUAC, KRCCC, and KPCC). Furthermore, the correlation between high NEIL3 expression and the triple-negative phenotype (estrogen receptor (ER) negative, progesterone receptor (PR) negative, HER2 negative phenotype) supports an important feature of a specific subgroup of breast cancer with poorer prognosis (Figure 1(f)). Our data are consistent with the observation of the presence of other DNA repair and replication genes (e.g. DNA polymerase θ (POLQ)) in tumors that induce poor OS of patients.36,37 In contrast, in stomach cancer and colorectal cancer patients, high NEIL3 expression was associated with better cancer-specific survival. Therefore, NEIL3 overexpression may play a very different role in cancers arising from different tissues, as other studies have shown. 38 Interestingly, overexpression of NEIL3 in tumors significantly increases the spread of PADC cancer cells to the lymph nodes sites (P < 0.001; Table 2), clearly demonstrating that up-regulation of NEIL3 impacts the outcome of different types of tumor progression.
Our genomic analysis indicates that NEIL3 overexpression is independent of gene amplification including oncogene amplification on chr4Q34. However, previous studies have identified several putative cell cycle-regulated promoter elements and demonstrated the involvement of the Ras-dependent extracellular signal-regulated kinase (ERK)–mitogen activated protein (MAP) kinases pathway in the induction of NEIL3 expression. 33 Results from this study and previous published work suggest that unbalanced expression of NEIL3 mimics an oncogenic effect by perturbing the replication forks and enhancing genetic instability in selected types of cancers. We also report the positive association between NEIL3 overexpression levels and genomic alteration in PADC, LUAC, LGG, and BC tumors. The overall tumor mutation load and chromosomal number variation significantly increased in four groups of cancers (P < 0.001, Figure 2(a) and (b)). NEIL3 overexpression increased mutation load in PADC, LUAC, LGG, and BC suggesting that NEIL3 may contribute to cancer development via accumulation of mutations similar to other DNA repair genes. 39 However, no significant difference in mutation load and chromosomal number variation was observed in KRCCC and KPCC tumors (Figure 2(a) and (b)), suggesting that NEIL3 overexpression may be involved in different DNA metabolism or cell cycle pathways to cause distinct tumor physiological processes. This observation may imply three possible scenarios: first, tumor cells with excess NEIL3 may speed up replication progression and enhance cancer cell proliferation. An increase in the level of cellular NEIL3 could be a protective response that occurs in cancer in an attempt to manage an increased DNA damage burden and mutation load in response to increased proliferation of cells. These processes may help to develop an unstable genome and trigger tumor initiation. Second, NEIL3 overexpression may impact the recruitment of other DNA repair factors such as homologous recombination (BRCA1/2) and interfere with the repair process at the replication fork that could contribute to tumorigenesis. Third, excessive NEIL3 could unbalance the BER pathway by generating more incisions than the remaining proteins can repair, resulting in accumulation of toxic BER repair intermediates that likely block replication forks.
Several studies have shown that deregulation of BER proteins induces genomic instability and cellular transformation and impacts treatment response for chemo and radiation therapy.40,41NEIL3 plays a critical role in DNA repair; it has been proven that up-regulated NEIL3 may contribute to intrinsic or acquired drug resistance, whereas depletion of NEIL3 led to spontaneous DNA damage and chromosomal instability.32,42 The positive correlation between NEIL3 overexpression and an increase in HR repair proteins such as BRCA1/2 might lead to an increase in DNA repair and promote resistance to different chemotherapy and radiation therapy. In addition, NEIL3 overexpressing tumors are strongly correlated with high proliferation rates and overexpression of genes that participate in cell cycle regulation, DNA replication and DNA repair genes (data not shown). In contrast, excess NEIL3 may interfere in replication associated DNA repair factors such HR, MMR, non-homologous end joining, NER repair, similar to others DNA glycosylase. 40 Furthermore, NEIL3 overexpression is negatively correlated with NER genes including XPA, XPC, and ERCC1 in PADC, LUAC, LGG, KPCC, KRCCC tumors. Previous works have demonstrated that BER is involved in cisplatin ICL repair and modulates cisplatin cytotoxicity.43–46 In addition, BER proteins are involved in repair of ICL induced DNA damage via a lesion bypass repair mechanism that leads to mutation.47,48 Conversely, cancer cells that have lost nucleotide excision repair genes such as XPA, XPC, and ERCC1 may depend on NEIL3 dependent ICL repair as an alternative repair pathway and can be rendered vulnerable by targeting the functional pathway using the principle of synthetic lethality, whereas non-cancer cells (with two functional repair pathways) would demonstrate resistance.
In the future, it will be necessary to uncover the impact of excess NEIL3 on genomic instability and cellular transformation. The overexpression of NEIL3 may also contribute to poor response to different types of chemotherapy and ionization radiation therapy in different types of cancer including PADC, LUAC, GBM, BC, and LUAC similar to other DNA repair genes.38,49NEIL3 overexpression in tumors may lead to resistance to chemotherapy, so inhibitors of repair proteins may have the potential to sensitize selected types of cancer. Given the prevalence of NEIL3 overexpression in tumors, such a strategy may have a significant impact on developing targeted cancer treatment. Overexpression of NEIL3 may affect the response of cells to DNA damaging anti-cancer treatment similar to other DNA repair enzymes. 50 Thus, NEIL3 overexpression potentially provides a growth advantage, allowing cell survival of oncogenic stress in cancer cells. It is possible that NEIL3 overexpression may be involved in recovery of stalled replication forks.
Our study likely provides a basis for the identification of NEIL3 overexpression associated genes that contribute to clinical heterogeneity and progression of cancer, an understanding that is critical to accurate classification of patients and appropriate therapy. 51 In addition, the total number of somatic coding mutations in a tumor is emerging as a promising biomarker for immunotherapy response in cancer patients. In particular, the mutational burden in NEIL3 overexpressed tumors (LUAC, non-small cell lung cancer (NSCLC), BC, PADC, and SCM) likely provides a base for future researchers to test whether those cancer types respond to a class of cancer immunotherapy drugs. Our results could identify overexpression of NEIL3 in selected types of cancer as a target or prognostic factor to better forecast potential outcomes and to determine which patients merit more aggressive therapeutic measures.
Supplemental Material
Supplement_Figure_1_revised – Supplemental material for Overexpression of NEIL3 associated with altered genome and poor survival in selected types of human cancer
Supplemental material, Supplement_Figure_1_revised for Overexpression of NEIL3 associated with altered genome and poor survival in selected types of human cancer by Oanh TN Tran, Serkalem Tadesse, Christopher Chu and Dawit Kidane in Tumor Biology
Supplemental Material
Supplement_Figure_2_revised – Supplemental material for Overexpression of NEIL3 associated with altered genome and poor survival in selected types of human cancer
Supplemental material, Supplement_Figure_2_revised for Overexpression of NEIL3 associated with altered genome and poor survival in selected types of human cancer by Oanh TN Tran, Serkalem Tadesse, Christopher Chu and Dawit Kidane in Tumor Biology
Footnotes
Acknowledgements
The authors thank Stephanie D. Scott for editing the manuscript.
Author contributions
O.T.N.T. performed data extraction and participated in data analysis; S.T. performed data analysis, data interpretation, and manuscript writing; C.C. performed data extraction and participated in data analysis; D.K. performed data analysis, interpretation, and manuscript writing. All authors read and approved the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: D.K. was supported by start-up funds from The University of Texas at Austin.
Availability of data and materials
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.