
Abstract
OCT4 is a crucial transcription factor that maintains self-renewal and pluripotency of embryonic stem and embryonic carcinoma cells. The human OCT4 gene can generate at least three variants (OCT4A, OCT4B, and OCTB1) via alternative splicing and alternative promoters. It has been previously reported that OCT4A is the main isoform, retaining stemness state in embryonic stem and embryonic carcinoma cells. There are several reports on the expression of OCT4A, OCT4B, and OCT4B1 in some cancers and tumor cells. The expression of OCT4 in cancer tissues and cell lines appeared to be highly controversial since it was believed that OCT4 is exclusively expressed in embryonic stem/embryonic carcinoma cells. Here, we are reporting the detection of a novel alternatively spliced variant of OCT4, OCT4B2, in several pluripotent and tumor cell lines. Moreover, the expression pattern of OCT4B2 in the course of neural differentiation of NT2 and NCCIT, embryonic carcinoma cells, was similar to that of OCT4A. OCT4B2 was highly expressed in undifferentiated cells; however, its expression was sharply downregulated upon induction of differentiation. Overexpression of OCT4B2 did not affect the distribution of cells in different cell-cycle phases of transfected cells, compared to the mock transfected cells. Interestingly, the expression of OCT4B2 transcript was elevated under the heat-shock induction. In conclusion, we are reporting a new variant of OCT4, which is expressed under different physiological conditions. The finding shed more light on complexity of OCT4 expression and functions.
Introduction
OCT4 (a member of POU transcription factor family) is an important transcription factor which regulates the maintenance of pluripotency and self-renewal in embryonic stem (ES) and embryonic carcinoma (EC) cells. 1 The expression of OCT4 is downregulated during stem cell differentiation. 2 Also, OCT4 knocking out resulted in early mortality of mice, due to the lack of inner cell mass (ICM) formation. 3 Mouse OCT4 messenger RNA (mRNA) and protein exist maternally in oocyte and zygote and provides the first step in the establishment of totipotency/pluripotency in early embryo. 4
OCT4 is present in the nuclei of all cells through the morula stage. At day 3.5, OCT4 becomes restricted to the ICM, and later, at day 4.5, to migrating cells of differentiating primitive endoderm. Following implantation, OCT4 expression is limited to primitive ectodermal cells. Expression in primordial cells is detectable at day 8.5. 5
Alternative splicing, alternative translation initiation, and alternative promoter usage result in several OCT4 transcripts and isoforms (OCT4A, OCT4B-265, OCT4B-190, OCT4B-164, and OCT4B1). The OCT4 isoforms differ in their abilities for DNA binding, transactivation, and self-renewal maintenance. OCT4A, but not OCT4B, is the main factor responsible for the stemness properties in ES and EC cells.6,7 OCT4B1 is primarily expressed in human ES and EC cells and is downregulated in differentiated cells. 8
OCT4A transcript is composed of five exons. Exon 1a contains 478 nucleotides and is specific for this isoform. The other OCT4 variants are generated from an alternative internal promoter, different from that of OCT4A. The internal promoter is located within intron 1 of OCT4 gene and utilized for transcription of OCT4B and OCT4B1 variants. OCT4B is composed of common exons 2–5, similar to OCT4A, and a unique exon, Exon 1b, which originates from the distal part of intron 1.7,9 The remaining intron 2 in the OCT4B transcript, as a cryptic exon, can produce OCT4B1. Since intron 2 has an in-frame TGA stop codon, OCT4B1 produces a putative truncated protein which lacks the C-terminal domain. 8
OCT4B transcript has an Internal Ribosome Entry Site (IRES) element in Exon 1b-2 region and can generate three protein isoforms: OCT4B-265 with 265 amino acids, OCT4B-190 with 190 amino acids, and OCT4B-164 with 164 amino acids. All of the OCT4B proteins have the same C-terminal, but are different in their N-terminal domain.6,7,10
However, the function of OCT4 as a marker of pluripotency has been a source of controversy because an increasing number of studies have shown that OCT4 is expressed in various non-pluripotent cells including adult stem cells, cancer cell lines and tumor tissues, and normal differentiated cells.10,11
Using reverse transcription polymerase chain reaction (RT-PCR) approach, seven OCT4-processed pseudogenes have been recently detected in different human cell types. 12 Due to the high similarity to OCT4A variant, the pseudogenes could cause false-positive results in RT-PCR, if the DNase treatment prior to complementary DNA (cDNA) synthesis is inadequate.2,10
In this study, we have discovered a novel variant of OCT4, OCT4B2, during scanning expression pattern of OCT4 variants in a panel of different human pluripotent, cancer, and normal cell types.
Material and methods
Cell lines and cell culture
Various human cell lines were provided by Pasteur Institute and Avicenna Research Institute of Iran, Tehran. The human cell lines of T-cell lymphoma (Jurkat), Burkit’s lymphoma (Raji), ovary adenocarcinoma (OVCAR3), glioblastoma (U87), urinary bladder carcinoma, and pluripotent embryonic carcinoma (NCCIT) were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS), 100 mM sodium pyruvate, penicillin (100 u/mL), and streptomycin (100 µg/mL). Cell types of breast adenocarcinoma (MCF-7), pluripotent embryonic carcinoma (NT2), cervix adenocarcinoma (HeLa), glioblastoma (A172), medulloblastoma (Daoy), embryonic kidney (HEK-293), hepatocellular carcinoma (HepG2), and bone marrow normal fibroblast (HS5) were cultured in high-glucose Dulbecco’s Modified Eagle Medium (DMEM, 4500 mg/L) supplemented with 10% FBS and sodium pyruvate and penicillin/streptomycin as described above. Y-79 cell line was cultured in RPMI-1640 medium supplemented with 20% FBS. The human ES cell of RH6, human induced pluripotent stem (iPS) cells, and human mesenchymal stem cell were provided kindly by Royan Institute.
Heat-shock condition
The human embryonic kidney 293 (HEK-293) cells and medulloblastoma (Daoy) cells were cultured in high DMEM as described in cell culture section and were heat-shock treated at 45°C for 1 hour. The heat-shock condition authenticity was verified by RT-PCR analysis of heat-shock protein of HSP70.
RNA extraction and cDNA synthesis
RNA extraction was performed using TRIzol reagent (Invitrogen) following the instruction of manufacturer. Quantity and quality of isolated RNAs were evaluated by spectrometry and electrophoresis on the agarose gel. Total RNAs were digested with RNase-free DNaseI (Fermentas) to remove any unwanted DNA contamination. The first strand of cDNAs was synthesized using the Reverse Transcription System (Fermentas). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control to assess the quality of synthesized cDNAs.
RT-PCR
RT-PCR was performed with the red master mix (Amplicon) according to the instruction of manufacturer. Primer set FA/R2b, FB1/RB5, and FB1/R2d were used to amplify OCT4A, OCT4B, and OCT4B1, respectively. The sequences of used primers were as follow: FA, 5′-′CTTCTCGCCCCCTCCAGGT-3′; R2b, 5′-AAATAGAACCCCCAGGGTGAGC-3′; FB1, 5′-AGACTATTCCTTGGGGCCACAC-3′; R2d, 5′-CTCAAAGCGGCAGATGGTCG-3′; R2c, 5′-CCCCCTGTCCCCCATTCCTA-3′; RB5, 5′-GGCTGAATACCTTCCCAAATAGA-3′; and FB2, 5′-CTGGGGAGAGTATATACTGAATTTAGCTTC-3′. GAPDH primer sequences were as follows: F, 5′-TGTGGGCATCAATGGATTTGG-3′; R, 5′-ACACCATGTATTCCGGGTCAAT-3′.
RT-PCR approach was performed using 0.5 µL of cDNA and No-RT samples and 4 pmol of mixed forward and reverse primers in total volume of 10 µL. PCR for OCT4B and OCT4B1 transcripts was performed under the following cycling conditions: initiation at 94°C for 4 min, amplification for 35 cycles with denaturation at 94°C for 30 s, and annealing and extension at 65°C for 30 s, with a final extension at 72°C for 7 min. The thermal profile for GAPDH was performed for 28 cycles with annealing of 58°C and extension of 15 s.
The size of amplified PCR products was 245-, 267-, and 116-bp for OCT4B, OCT4B1, and GAPDH, respectively. Consequently, PCR products were confirmed by DNA sequencing.
Specific primers to amplify OCT4B2 variant by RT-PCR were as follow: B2F (5′-CAATTAGGATGCCTCTCTTCATAAC-3′) and B2R (5′-GGATTTTAAAAGGCAGAAGACTTGTAAG-3′). The predicted size for this amplified fragment was 320 bp.
Cell differentiation
Neural differentiation of NT2 and NCCIT cells was carried out using all-trans retinoic acid (RA; Sigma-Aldrich). Then, NT2 cells were harvested at 0, 3, 7, 14, and 21 days after treatment with RA, and NCCIT cells were collected after 21 days and prepared for RNA extraction. Neural differentiation of NT2 and NCCIT cells verified by RT-PCR analysis of neural marker of MAP2.
Western blotting
Untransfected NCCIT cells were lysed with 1 mL of lysis buffer (1% Triton X-100, 5 mM ethylenediaminetetraacetic acid (EDTA), 50 mM Tris-HC1, pH = 7.4, 150 mM NaCl, and 0.1% sodium dodecyl sulfate (SDS)), 1% protease inhibitor cocktail, and phosphatase inhibitor (Sigma). Protein concentrations were measured by the Bradford Protein Assay reagent. Briefly, 20 µg of cell lysates were isolated on 12.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene difluoride (PVDF) membrane (Millipore). Blocking was carried out with 5% skim milk in Tris-buffered saline (TBS), containing 0.05% Tween 20 (TBS-T). The membrane was then incubated for 1 h with primary mouse anti-Oct-3/4 sc-5279 antibody (Santa Cruz Biotechnology) diluted in 3% blocking buffer and washed with phosphate-buffered saline (PBS) for 30 min at room temperature. A secondary horseradish peroxidase (HRP)-conjugated sheep-anti-mouse antibody (Avicenna Research Institute) was added next and incubated at RT for 1 h. After washing, signals were detected using the Amersham ECL Prime Western Blotting Detection kit (GE Healthcare Life Sciences) and quantified by Gel Logic 2200 (Kodak).
Cloning and sequencing
PCR products were extracted from 1.5% agarose gel, ligated into PTZ57R/T vector (TA cloning kit; Fermentas), and incubated at 4°C overnight. Heat-shock transformation was performed with DH5α competent cells (Escherichia coli) and plated onto Luria broth (LB)-agar plates containing 100 µg/mL ampicillin. Positive colonies were selected by colony PCR and expanded, and amplified plasmids were recovered using miniprep kit (GeneAll) according to the manufacturer’s instruction. Finally, sequencing was carried out for all of amplified OCT4 transcript variants.
Production of OCT4B2 construct
The nucleotide sequence of OCT4B2 transcript was obtained by splicing by overhang extension (SOE)-PCR and cloned into PTG19 T-vector (TA cloning vector, vivantis). Using EcoRI and SalI enzymes, OCT4B2 sequence was digested and sub-cloned into pIRES2-EGF expression vector. Positive colonies were selected by colony PCR using forward primer of vector and reverse primer of OCT4B2.
Cell-cycle analysis
HeLa and Daoy cells were transfected with OCT4B2 construct, and cells were harvested 42 h after transfection. DNA of collected cells was stained by propidium iodide (PI). Prepared cells were analyzed by flow cytometry (Becton Dickinson Bioscience).
Results
Identification of a novel OCT4 spliced variant, OCT4B2
Using FB2/R2d primer set (Figure 1(a)), we observed three PCR bands instead of 2 in the 5637 cell line (Figure 1(b)). The lack of any signal in No-RT sample suggested that the unexpected amplified fragment was not generated via DNA contamination, and hence, it was not a non-specific PCR product. Furthermore, the same product re-appeared in nested PCR (data not shown). The DNA sequencing data confirmed the authenticity of the 1175-bp and 950-bp PCR products as OCT4B1 and OCT4B, respectively. The sequence result also confirmed the identity of 545-bp fragment, as a novel spliced variant of OCT4, designated as OCT4B2 (GenBank accession no. KJ700311). Similar to OCT4B1, OCT4B2 retains parts of intron 1, but missing a 630-bp fragment of the intron (Figure 1(c)).

Reverse transcription (RT)-PCR analysis. (a) A schematic representation of OCT4B1 variant, showing the primer locations on the OCT4B1 transcript. (b) RT-PCR analysis using forward primer of FB2 and reverse primer of R2d in 5637 cell line. Two expected 1175-bp and 950-bp fragments were OCT4B1 and OCT4B variants, respectively. DNA sequencing revealed that 545-bp fragment was a novel OCT4 variant, OCT4B2. A no-RT control was used in parallel to confirm a lack of DNA contamination in cDNA samples. (c) A schematic picture to depict the genomic organization of OCT4B2 splice variant. OCT4B2 transcript is very similar to the OCT4B1, in retaining E2d, but lacks a 630-bp fragment of E1b. (d) OCT4B2 sequence conservation analysis by UCSC database. Exons 2, 3, 4, and 5 are conserved between several species, but Exon 1b and Exon 2b are not conserved.
The sequence data demonstrated the presence of a noncanonical AT at 5′ site and a AC at 3′ site of splicing. The introns containing AT-AC have highly conserved 5′ splice site and a presumptive branch site TCCTRAY (TCCTTAAA in exon2a of OCT4B1). AT-AC splicing at position 219-847 could lead to the generation of OCT4B2, which lacks 630-bp of exon2a.
Expression pattern of OCT4B2 in different human cell types
Using the specific primer set B2F/B2R, we employed an RT-PCR approach to examine a potentional expression of OCT4B2 in various pluripotent and non-pluripotent human cell lines. Our data revealed that OCT4B2 is highly expressed in iPS cells and also in MCF-7 (breast adenocarcinoma) and 1321N1 (Astrocytoma) cell lines (RH6, NCCIT, NT2, MSC, iPSCs, cord blood) and cancer cell lines (MCF-7, HepG2, HEK-293, 1321N1, Raji, OVCAR3) and there were no sign of expression in the other examined cell lines (Figure 2).

Expression analysis of OCT4B2 by RT-PCR assay. The expression pattern of OCT4B2 was considered in different human pluripotent and cancer cell lines. Our data revealed that OCT4B2 is expressed in human pluripotent cells (NT2, NCCIT, RH6, MSC, iPSCs, and cord blood) and some cancer cell lines (MCF-7, HepG2, HEK-293, 1321N1, Raji, and OVCAR3). GAPDH was used as internal control.
OCT4B2 variant is downregulated upon the induction of neural differentiation of NT2 and NCCIT cells
To study the expression of OCT4B2 in the course of neural differentiation of NT2 and NCCIT, we treated EC cells (NT2 and NCCIT) with all-trans-RA to induce differentiation, and RT-PCR analysis was performed with specific primers for OCT4B2 (B2F/B2R). In this experiment, OCT4B2 was downregulated during the course of differentiation (Figure 3).

Expression pattern of OCT4B2 variant during differentiation of NT2 and NCCIT cells. (a(1)) Neural differentiation of NT2 was performed by application of retinoic acid and cells were harvested 0, 3, 7, 14, and 21 days after treatment. RT-PCR analysis indicated that the expression pattern of OCT4B2 variant was similar to that of OCT4A during early differentiation of NT2 cells. OCT2B2 was expressed in days 0 and 3 and its expression was downregulated after starting differentiation. (a(2)) RT-PCR analysis of neural marker of MAP2, and the expression of MAP2 in days 14 and 21 confirmed the neural differentiation of NT2 cells. (b(1)) Neural differentiation of NCCIT cells was performed by retinoic acid and cells were harvested 0 and 21 days after differentiation. RT-PCR confirmed the downregulation of OCT4B2 with induction of differentiation. (b(2)) Western blot analysis of OCT4A isoform before and after differentiation of NCCIT cells, and OCT4A protein was downregulated after differentiation of NCCIT cells. GAPDH was used as an internal control for RT-PCRs.
Cell-cycle alteration following overexpression of OCT4B2
OCT4B2 variant was overexpressed in HeLa and Daoy cells, and cell-cycle analysis was carried out with PI. There were no significant difference between cells transfected with mock pIRES2-EGFP vector and cells transfected with OCT4B2 construct (Figure 4).
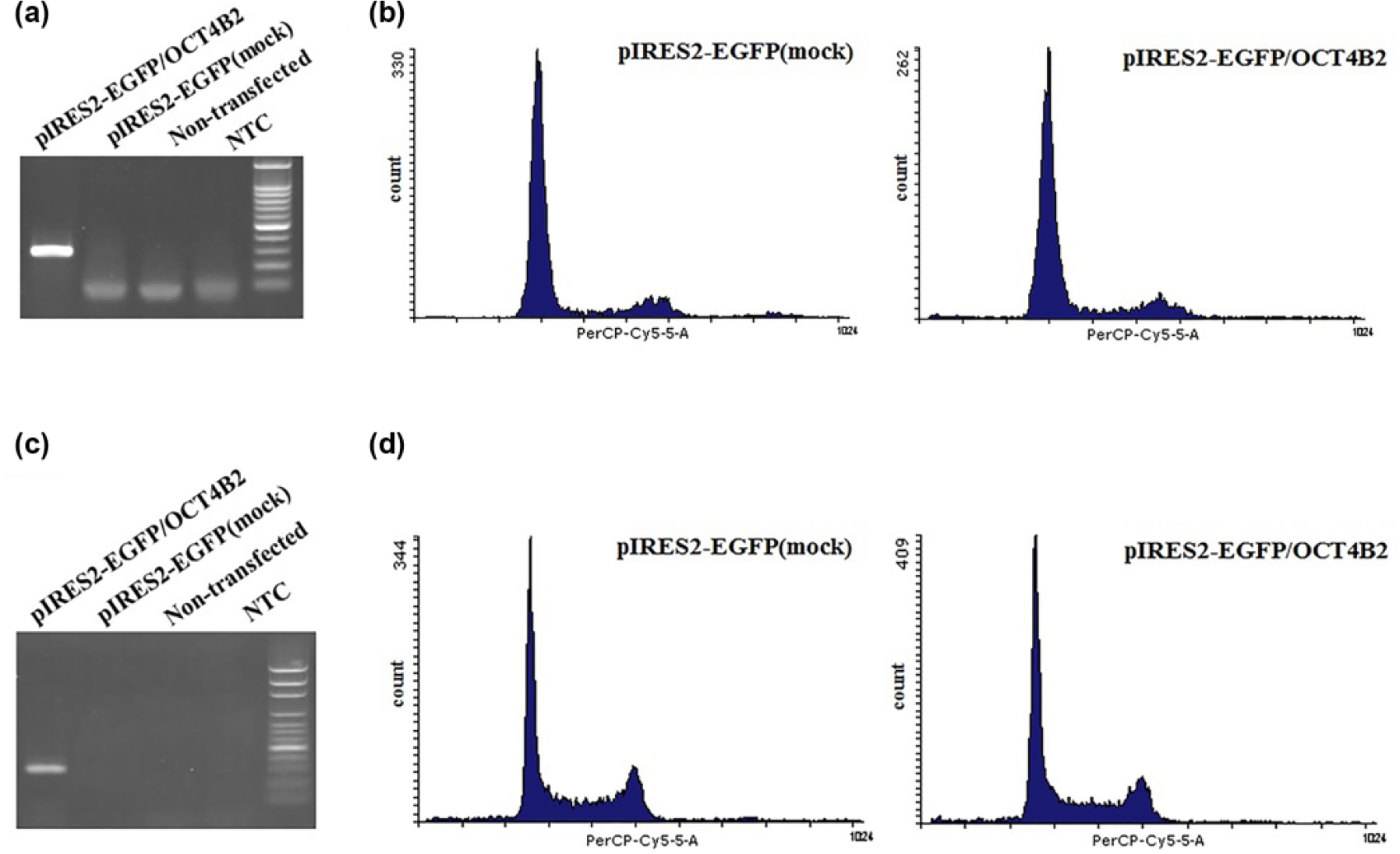
Functional analysis of OCT4B2 variant. OCT4B2 was overexpressed in HeLa cells and Daoy cells and cell-cycle alteration was examined by flow cytometry. (a and c) RT-PCR analysis for HeLa and Daoy cells transfected with OCT4B2 construct, mock vector (pIRES2-EGFP) and non-transfected cells; OCT4B2 mRNA was detected just in cells transfected with OCT4B2 construct. (b and d) Flow cytometry analysis revealed no significant change in the number of cells in G1, S, G2, and sub-G1 phases, 42 h after overexpression of OCT4B2, compared to the control cells transfected with mock vector and non-transfected HeLa cells (p values were >0.05; sub-G1 p value = 0.672, G1 p value = 0.556, S p value = 0.673 and G2 p value = 0.591) and Daoy cells (p values were >0.05; sub-G1 p value = 0.098, G1 p value = 0.175, S p value = 0.624 and G2 p value = 0.853).
The OCT4B2 is upregulated in HEK-293 and OVCAR3 cells in response to heat-stress condition
To determine whether the heat-stress conditions have any differential effects on the level of OCT4B2 production, the level of OCT4B2 transcript was determined after incubating HEK-293 and OVCAR3 cells at 45°C for 1 h. As well as, we evaluated the expression of OCT4B and OCT4B1 in these samples. Our data revealed that the expression of OCT4B and OCT4B1 was elevated which is concordant with the results of our previous studies. 13 Moreover, the expression of the OCT4B2 was changed in this experiment. Real-time PCR data showed that the expression of OCT4B2 transcript was significantly increased under the heat-shock pressure (Figure 5).

Expression analysis of OCT4B2 under heat-stress condition. (a and c) RT-PCR analysis of OCT4B2. RT-PCR was performed using B2F/R2b primers in HEK-293 and OVCAR3 cells before and after heat-stress. An expected 454-bp PCR product was corresponded to be the novel variant of OCT4B2. (b and d) Real-time PCR of OCT4B, OCT4B1, and OCT4B2 in HEK-293 and OVCAR3 cells in comparison with the heat-shocked ones. Based on our data, OCT4B, OCT4B1, and OCT4B2 upregulated after heat-stress.
Discussion
OCT4 gene has been introduced as a key regulator of stemness and maintenance of self-renewal and pluripotency. 14 The expression of this gene has been known to be a hallmark of ES and EC cells. Recent studies, however, indicated that OCT4 expression is not restricted to stem/pluripotent cells and its expression has been detected in normal and tumor tissues and cancer cell lines.15–17 The existence of several splice variants and pseudogenes for this gene and the lack of specific discriminating primers may increase mistakes in interpretation of data acquired by RT-PCR and Real-Time PCR. 18 OCT4A is the most important variant of OCT4 which confers stemness properties in ES and EC cells. This variant is expressed in undifferentiated ES cells, and at the end of the human/mouse embryo evolution, its expression is restricted to the primordial germ cells (PGCs). 19 OCT4B transcript can generate three isoforms by alternative translation initiation: OCT4B-164 with 164 amino acids, OCT4B-190 with 190 amino acids, and OCT4B-265 with 265 amino acids. 10 Several recent studies indicated that OCT4B-190 and OCT4B-265 isoforms are upregulated during cell stresses.7,20 OCT4B1 is a newly discovered variant of OCT4 that is expressed in pluripotent and undifferentiated cells and its expression is downregulated during induction of differentiation. 21 Also, the OCT4B1 expression level is upregulated in some human cancers such as bladder and gastric cancers.8,22–24 Furthermore, it is shown that the expression ratio of OCT4B1/OCT4B transcripts and proteins was significantly elevated under heat-stress condition. 13 All over, these studies showed that OCT4B group variants have important roles in cell response to stress conditions.
In this study, during evaluating the expression pattern of OCT4 variants, namely, OCT4A, OCT4B, and OCT4B1 with RT-PCR, we discovered a novel OCT4 splice variant, designated as OCT4B2, which is expressed in pluripotent cells (NT2, NCCIT, RH6, MSC, iPSCs, cord blood) and some cancer cell lines (MCF-7, HepG2, HEK-293, 1321N1, Raji, and OVCAR3).
Also, the expression pattern of OCT4B2 was considered during neural differentiation of NT2. RT-PCR analysis indicated that OCT4B2 is expressed in undifferentiated NT2 cells and its expression is quickly downregulated after starting differentiation.
Furthermore, we used HEK-293 and OVCAR3 cells to examine the expression of OCT4B2 under heat-shock condition. Our data revealed that the OCT4B, OCT4B1, and specifically OCT4B2 expression was elevated during heat-shock pressure. Since these OCT4 variants might encode from a different promoter in the intron1 of the OCT4 gene, we suggest that cell stresses probably affect this promoter of OCT4 and impress on the expression of B group variants of OCT4 gene; however, more experimental studies are needed to prove the validity of this hypothesis.
We predicted OCT4B2 protein by ExPASy Translate Tool and found that this variant can produce a 164 amino acid protein which was similar to OCT4B-164 isoform. OCT4B2 5′-UTR differs with that of in OCT4B1 which has no effect on its protein sequence but might affect translation deficiency, and this could be a regulatory mechanism for OCT4B2 translation rate in different conditions.
In conclusion, we introduced a novel OCT4 variant, designated as OCT4B2, which downregulated during differentiation and upregulated under heat-shock stress. This variant is expressed in different pluripotent and cancer cell lines and is very similar to the OCT4B1, but has lost a part of its 5′-UTR by splicing. We assume that difference of 5′-UTR of OCT4B2 might affect translation efficiency. Also, we suppose that OCT4B1 might be spliced in its 5′-UTR and produce OCT4B2 in special conditions.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study received financial support from Iranian Stem Cell Council (IRSCC).