
Abstract
The effect of the dipeptidyl peptidase-IV inhibitor gemigliptin alone or in combination with the heat shock protein 90 inhibitor NVP-AUY922 (AUY922) on survival of thyroid carcinoma cells was elucidated. The SW1736 and TPC-1 human thyroid carcinoma cells were used. Cell viability, the percentage of viable cells, cytotoxic activity, the percentage of apoptotic cells, and mitochondrial membrane potential were measured. To evaluate the combined effect of gemigliptin with AUY922, the interactions were estimated by calculating combination index. Gemigliptin led to cell death in conjunction with overexpression of the phosphorylated protein levels of Akt, extracellular signal–regulated kinase 1/2, and adenosine monophosphate–activated protein kinase. In gemigliptin-treated cells, wortmannin augmented cell death, whereas AZD6244 and compound C did not affect cell survival. Wortmannin decreased phosphorylated adenosine monophosphate–activated protein kinase protein levels, and AZD6244 increased phosphorylated Akt protein levels. Meanwhile, cotreatment of both gemigliptin and AUY922, compared with treatment of AUY922 alone, potentiated cell death. All the combination index values were lower than 1.0, suggesting synergistic cytotoxicity of gemigliptin with AUY922. In treatment of both gemigliptin and AUY922, compared with AUY922 alone, the protein levels of total and phosphorylated Akt, phosphorylated extracellular signal–regulated kinase 1/2, and phosphorylated adenosine monophosphate–activated protein kinase increased without alteration in those of total extracellular signal–regulated kinase 1/2 and total adenosine monophosphate–activated protein kinase. The percentage of apoptotic cells increased. The protein levels of Bax and cleaved poly (ADP-ribose) polymerase increased, whereas Bcl2 protein levels were unchanged, resulting in increment of Bax/Bcl2 ratio. Transfection of Bax small interfering RNA did not cause any variation in cell viability, the percentage of viable cells and cytotoxic activity. Our results demonstrate that gemigliptin exerts a cytotoxic activity with concomitant alterations in expression of Akt, extracellular signal–regulated kinase 1/2, and adenosine monophosphate–activated protein kinase in thyroid carcinoma cells. Furthermore, gemigliptin synergizes with AUY922 in induction of cytotoxicity via regulation of Akt, extracellular signal–regulated kinase 1/2, and adenosine monophosphate–activated protein kinase as well as involvement of Bcl2 family proteins in thyroid carcinoma cells.
Introduction
Follicular cell–derived thyroid cancer is divided as well-differentiated thyroid carcinoma (WDTC) and undifferentiated thyroid carcinoma (UDTC) according to differentiation of carcinoma cells. 1 WDTC including papillary thyroid carcinoma (PTC) and follicular thyroid carcinoma (FTC) is the most common subtype with an excellent prognosis, but it is refractory to radioactive iodine (RAI) therapy in two thirds of patients with metastatic lesions. 1 UDTC including anaplastic thyroid carcinoma (ATC) is a highly aggressive malignancy characterized as extrathyroidal invasion and distant metastasis with a very poor prognosis.2,3 In clinical situation, the treatment of patients with RAI therapy-refractory WDTC and UDTC is sometimes problematic because of unresponsiveness to conventional therapies, and thus novel therapeutic strategies to improve the outcome of the patients are under consideration.2,3
Dipeptidyl peptidase-IV (DPP-IV), known as CD26, belongs to serine protease expressed on a variety of cells and cleaves N-terminal depeptides from peptide substrates.4,5 In human body, DPP-IV rapidly inactivates endogenous incretin such as glucagon-like peptide-1.4,5 DPP-IV inhibitor stimulates secretion of endogenous incretin, which suppresses blood glucose levels.4,5 Currently, DPP-IV inhibitor is widely used as monotherapy or combination therapy for treatment of patients with type 2 diabetes mellitus.4,5
DPP-IV modulates diverse cellular processes including survival, proliferation, and differentiation and thereby enhances or diminishes tumorigenesis relying on types or phases of tumors. 6 In this regard, DPP-IV is overexpressed or underexpressed in human solid tumor tissues, which is associated with survival, suggesting significance of DPP-IV as a potential diagnostic marker and therapeutic target in solid tumors. 6 With respect to activity of DPP-IV in thyroid carcinoma cells, it was shown that DPP-IV was seldom active in thyroid follicular cells, whereas it was upregulated in thyroid carcinoma cells.7–9 Furthermore, it was reported that expression of DPP-IV in aspirated or surgical specimens was abundant in WDTC and absent in follicular adenoma, implying consequence of DPP-IV as a useful tool to aid in distinguishing malignant from benign thyroid tumors. 10 In regard to the influence of DPP-IV inhibitor on cancer, the DPP-IV inhibitors sitagliptin and saxagliptin activate migration and invasion in various cancer cell lines and accelerate development of metastatic nodules in experimental animal models. 11 By contrast, it was shown that DPP-IV inhibitor was not related to increased risk of metastasis in type 2 diabetic patients with breast, prostate, and gastrointestinal cancers. 12 However, the impact of DPP-IV inhibitor on thyroid carcinoma cells has not been elucidated.
Gemigliptin is a novel and potent DPP-IV inhibitor and improves glucose tolerance by elevating insulin secretion and reducing glucagon secretion during oral glucose tolerance test. 13 Moreover, gemigliptin attenuates glycosylation of hemoglobin and ameliorates β-cell damage in high-fat diet/streptozotocin-induced diabetic mice. 14 Besides glucose-lowering properties, gemigliptin inhibits expression of vascular adhesion molecules and inflammatory cytokines in human vascular endothelial cells through Akt and adenosine monophosphate–activated protein kinase (AMPK). 15 In addition, gemigliptin represses tunicamycin-induced endoplasmic reticulum stress, apoptosis, and inflammation in cardiomyocytes. 16 Furthermore, gemigliptin mitigates apoptosis in the heart of db/db mice, a type 2 diabetic animal model, with phosphorylation of extracellular signal–regulated kinase (ERK) 1/2. 17 Despite the pleiotropic effects of gemigliptin, the influence of gemigliptin on malignant cells including thyroid carcinoma cells has not been investigated.
Heat shock protein 90 (hsp90), a molecular chaperone, plays a pivotal role in promoting survival in response to environmental stress in cells. 18 While hsp90 exists in an activated state in complex with cochaperones in cancer cells, it exists in an uncomplexed latent state in normal cells. 19 At present, several hsp90 inhibitors alone or in combination with other chemotherapeutic agents are undergoing clinical trials in patients with a variety of cancers. 20 NVP-AUY922 (AUY922), a synthetic hsp90 inhibitor, belongs to the family of isoxazole resorcinol based on 4,5-diarylisoxazole scaffold and has a high affinity for NH2-terminal nucleotide site of hsp90. 21 AUY922 exerts a potent anti-tumor activity and synergizes with other chemotherapeutic agents in induction of death of cancer cells.22–24 We recently reported that AUY922 had a synergistic activity with the histone deacetylase inhibitor PXD101 in induction of death of ATC cells. 25 However, the impact of gemigliptin in combination with AUY922 on thyroid carcinoma cells has not been identified.
The aim of this study was to evaluate the effect of gemigliptin alone or in combination with AUY922 on survival of thyroid carcinoma cells. Our results demonstrate that gemigliptin causes cell death and has a synergistic cytotoxicity with AUY922 in thyroid carcinoma cells.
Materials and methods
Materials
RPMI-1640 medium, fetal bovine serum (FBS), and streptomycin/penicillin were obtained from Life Technologies (Carlsbad, CA, USA). Gemigliptin was provided by LG Life Sciences (Seoul, Korea). AUY922, the phosphoinositide 3-kinase (PI3K)/Akt inhibitor wortmannin, the mitogen-activated protein kinase kinase (MEK)/ERK inhibitor AZD6244, and the AMPK inhibitor compound C were purchased from BioVision (Linda, CA, USA) and dissolved in dimethyl sulfoxide (DMSO), which was provided to the control within permissible concentrations. The final concentration of the vehicle DMSO in the control did not exceed 0.1% in all treatments. The primary antibodies raised against total and phosphorylated ERK1/2 (phospho-ERK1/2; Thr402/Tyr404), total and phospho-AMPK (Thr172), hsp90, Bcl2, Bax, and cleaved poly (ADP-ribose) polymerase (PARP) were obtained from Cell Signaling Technology (Danvers, MA, USA). The primary antibodies raised against total and phospho-Akt (Ser473) and β-actin were purchased from Sigma (St. Louis, MO, USA). All other reagents were obtained from Sigma unless otherwise stated.
Cell culture
For experiments, SW1736 and TPC-1 human thyroid carcinoma cells were used. SW1736 cells were purchased from Cell Lines Service (CLS GmbH, Eppelheim, Germany) and grown in RPMI-1640 supplemented with 2 mM
CCK-8 assay
Cell viability was measured by the CCK-8 Assay Kit (Dojindo Molecular Laboratories, Kumamoto, Japan). Cells (5 × 103/100 μL) in each well on 96-well plates were incubated overnight and treated with gemigliptin and/or AUY922 for an additional 4 h at 37°C. Absorbance was measured using GloMax™ Discover System GM3000 (Promega, Madison, WI, USA). All experiments were performed in triplicate.
Multiplexed cytotoxicity assay
Cells (5 × 103/100 μL) were seeded in 96-well plates, and reagents of the MultiTox-Glo Multiplex Cytotoxicity Assay Kit (Promega) were added to the cells after treatments as indicated by the manufacturer’s protocol. Fluorescent and luminescent values were measured using GloMax™ Discover System GM3000 (Promega). Viability was calculated as a ratio of live/dead cells and expressed as percentage of untreated cells. All experiments were performed in triplicate.
Cytotoxicity assay
Cytotoxic activity was measured by the LDH Cytotoxicity Assay Kit (BioVision). Cells (5 × 103/100 μL) in each well on 96-well plates were incubated and centrifuged at 250g for 10 min. Supernatant of 100 μL was transferred in clear 96-well plates. After addition of reaction mixture (2.5 μL of catalyst solution in 112.5 μL of dye solution), cells were incubated for 30 min at room temperature. Absorbance was measured using GloMax™ Discover System GM3000 (Promega). All experiments were performed in triplicate.
Mitochondrial membrane potential assay
Mitochondrial membrane potential was measured by the JC-1 MMP Assay Kit (Abcam, Cambridge, MA, USA). Cells (1 × 104/100 μL) in each well on 96-well plates were incubated with 20 µM of JC-1 in 1× dilution buffer for 10 min at 37°C. Fluorescent values were measured using GloMax™ Discover System GM3000 (Promega). All experiments were performed in triplicate.
Fluorescence-activated cell sorting analysis
The dead cells were analyzed by the Annexin V-FITC Apoptosis Detection Kit (BD Biosciences Pharminogen, San Diego, CA, USA). Cells (1 × 105/mL) in each well on six-well plates were incubated, harvested, washed, and fixed according to the manufacturer’s protocol. Fluorescein isothiocyanate (FITC) Annexin V and/or propidium iodide (PI) in 1× binding buffer was added for 15 min at room temperature and analyzed using a FACSort flow cytometry (Becton Dickinson, San Jose, CA, USA) and CellQuest software program (Becton Dickinson). The percentage of apoptotic cells was calculated according to the following equation: dead cells (%) = (Annexin V–stained cells + Annexin V– and PI-stained cells)/all cells × 100. All experiments were performed in triplicate.
Drug combination analysis
Combination index (CI) and isobologram were calculated by CalcuSyn program version 2.11 (Biosoft, Cambridge, UK), and the effect of drug interactions was quantitatively determined. CI values less than 1.0, equal to 1.0, and greater than 1.0 indicate synergism, additivity, and antagonism, respectively. The isobologram is formed by plotting the doses of each drug required for 50% inhibition (ED50) on the x- and y-axis and connecting them to draw a line segment, which is ED50 isobologram. Combination data points that fall on, below, and above the line segment indicate additivity, synergism, and antagonism, respectively. All reactions were performed in triplicate.
Western blotting
The total protein was extracted by radioimmunoprecipitation assay (RIPA) buffer (Sigma) containing 1× protease inhibitor cocktail and 1× phophatase inhibitor cocktail set V (Calbiochem, La Jolla, CA, USA). Western blotting was performed using specific primary antibodies and horseradish peroxidase–conjugated anti-rabbit and anti-mouse secondary antibodies. Bands were detected using ECL Plus Western Blotting Detection System (Thermo Fisher Scientific, Rockford, IL, USA). All reactions were performed in triplicate.
Transfection of small interfering RNA
A Bax small interfering RNA (siRNA) was purchased from Cell Signaling Technology. Cells were transfected with siRNA using Lipofectamine 3000 Reagent (Life Technologies) according to the manufacturer’s protocol. Transfection efficiency was tested by western blotting.
Statistical analysis
All data are expressed as mean ± standard error (SE). Data were analyzed by unpaired Student’s t test or analysis of variance (ANOVA) as appropriate. A p value less than 0.05 was considered to be statistically significant. All analyses were performed using SPSS program version 24.0 (SPSS, Chicago, IL, USA).
Results
Gemigliptin induces cell death with concomitant alterations in Akt, ERK1/2, and AMPK in thyroid carcinoma cells
To identify the impact of gemigliptin on survival of SW1736 and TPC-1 human thyroid carcinoma cells, cells were treated with gemigliptin at 0.25, 0.5, 0.75, 1, 1.5, and 2 mM for 24 and 48 h, and cell viability was measured using CCK-8 assay (Figure 1(a)). As a result of treatment, cell viability was reduced in a dose- and time-dependent manner. Cells were treated with at the same doses for 48 h, and then the percentage of viable cells was measured using multiplexed cytotoxicity assay (Figure 1(b)), and cytotoxic activity was measured using cytotoxicity assay (Figure 1(c)). After treatment, the percentage of viable cells was reduced, and cytotoxic activity was elevated.
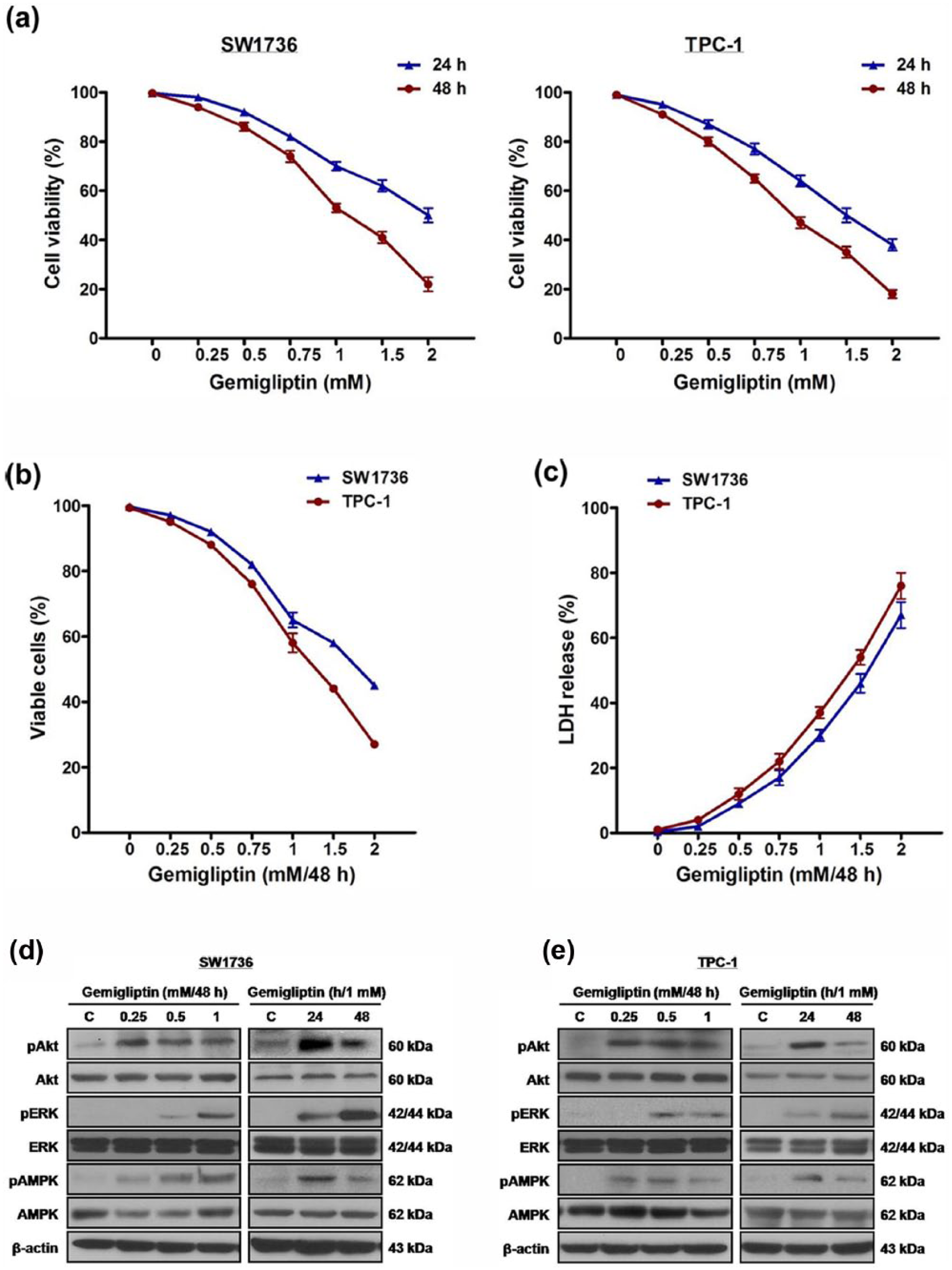
The effect of gemigliptin on cell survival and expression of Akt, ERK1/2, and AMPK in thyroid carcinoma cells. (a) SW1736 and TPC-1 cells were treated with gemigliptin at 0.25, 0.5, 0.75, 1, 1.5, and 2 mM for 24 and 48 h, and cell viability was measured using CCK-8 assay. (b and c) SW1736 and TPC-1 cells were treated with gemigliptin at 0.25, 0.5, 0.75, 1, 1.5, and 2 mM for 48 h, and (b) the percentage of viable cells was measured using multiplexed cytotoxicity assay and (c) cytotoxic activity was measured using cytotoxicity assay. (d) SW1736 and (e) TPC-1 cells were treated with gemigliptin at 0.25, 0.5, and 1 mM for 48 h and at 1 mM for 24 and 48 h, and the total and phospho-protein levels of Akt, ERK1/2, and AMPK were measured. All experiments were performed in triplicate. The blots are representative of independent experiments. Data are expressed as mean ± SE.
Gemigliptin represses expression of adhesion molecules and cytokines via activation of Akt and AMPK in human vascular endothelial cells. 15 In this study, to investigate the influence of gemigliptin on Akt and AMPK as well as ERK1/2 in thyroid carcinoma cells, cells were treated with gemigliptin at 0.25, 0.5, and 1 mM for 48 h and at 1 mM for 24 and 48 h, after which the total and phospho-protein levels of Akt, ERK1/2, and AMPK were measured (Figure 1(d) and (e)). Under treatment, the phospho-protein levels were elevated, while the total protein levels were not changed.
Inactivation of Akt, but not ERK1/2 and AMPK, potentiates gemigliptin-induced death of thyroid carcinoma cells
Gemigliptin stimulates Akt and AMPK activities in human vascular endothelial cells, whereas ERK1/2 suppresses AMPK activity in human leukemia cells.15,26 In this study, to explore the relationship among Akt, ERK1/2, and AMPK in thyroid carcinoma cells under exposure of gemigliptin, cells were administered with the Akt inhibitor wortmannin and the ERK1/2 inhibitor AZD6244 before treatment of gemigliptin at 1 mM for 48 h, and the total and phospho-protein levels of Akt, ERK1/2, and AMPK were measured (Figure 2(a)). In gemigliptin-treated cells, wortmannin decreased phospho-AMPK protein levels in SW1736 and TPC-1 cells. In contrast, wortmannin decreased phospho-ERK1/2 protein levels in SW1736 cells and increased the protein levels in TPC-1 cells. Meanwhile, AZD6244 increased phospho-Akt protein levels in SW1736 and TPC-1 cells. By contrast, AZD6244 decreased phospho-AMPK protein levels in SW1736 cells and increased the protein levels in TPC-1 cells.

The role of Akt, ERK1/2, and AMPK in survival of thyroid carcinoma cells under exposure of gemigliptin. (a) SW1736 and TPC-1 cells were administered with wortmannin and AZD6244 before treatment of gemigliptin at 1 mM for 48 h, and the total and phospho-protein levels of Akt, ERK1/2, and AMPK were measured. (b–d) SW1736 and TPC-1 cells were administered with wortmannin and AZD6244 prior to treatment of gemigliptin at 1 mM for 48 h, and (b) cell viability, (c) the percentage of viable cells, and (d) cytotoxic activity were measured. All experiments were performed in triplicate. The blots are representative of independent experiments. Data are expressed as mean ± SE (*p < 0.05 vs control and †p < 0.05 vs cells treated with gemigliptin alone).
To examine the relevance of the signal proteins to survival of thyroid carcinoma cells exposed to gemigliptin, cells were administered with wortmannin and AZD6244 prior to treatment of gemigliptin at 1 mM for 48 h, and then cell viability (Figure 2(b)), the percentage of viable cells (Figure 2(c)), and cytotoxic activity (Figure 2(d)) were measured. In cells treated with gemigliptin, wortmannin decreased cell viability and the percentage of viable cells and increased cytotoxic activity. In contrast to wortmannin, AZD6244 did not alter cell viability, the percentage of viable cells, and cytotoxic activity. When cells were administered with the AMPK inhibitor compound C before treatment of gemigliptin at 1 mM for 48 h, cell viability, the percentage of viable cells, and cytotoxic activity were not affected by administration of compound C (Supplementary Figure 1).
Gemigliptin synergizes with AUY922 in induction of death of thyroid carcinoma cells
We recently reported that AUY922 had a synergistic cytotoxicity with the histone deacetylase inhibitor PXD101 in ATC cells, and thus the effect of gemigliptin in combination with AUY922 on survival of thyroid carcinoma cells was evaluated. 25
First, to clarify the combined effect of two agents, cells were simultaneously treated with both gemigliptin and AUY922, and the interactions were estimated by calculating CI using Chou–Talalay equation, where CI < 1.0 indicates synergism, CI = 1.0 indicates additivity, and CI > 1.0 indicates antagonism (Figure 3(a) and (b), Table 1). Cell viability was measured using CCK-8 assay, and death rate was calculated as 100-cell viability (%). As a result of cotreatment, all the CI values were lower than 1.0 in combination of gemigliptin with AUY922. In the isobologram analysis, the combination data points were all located below the isobologram line at ED50, suggesting the synergism between gemigliptin and AUY922-induced cytotoxicity in thyroid carcinoma cells.
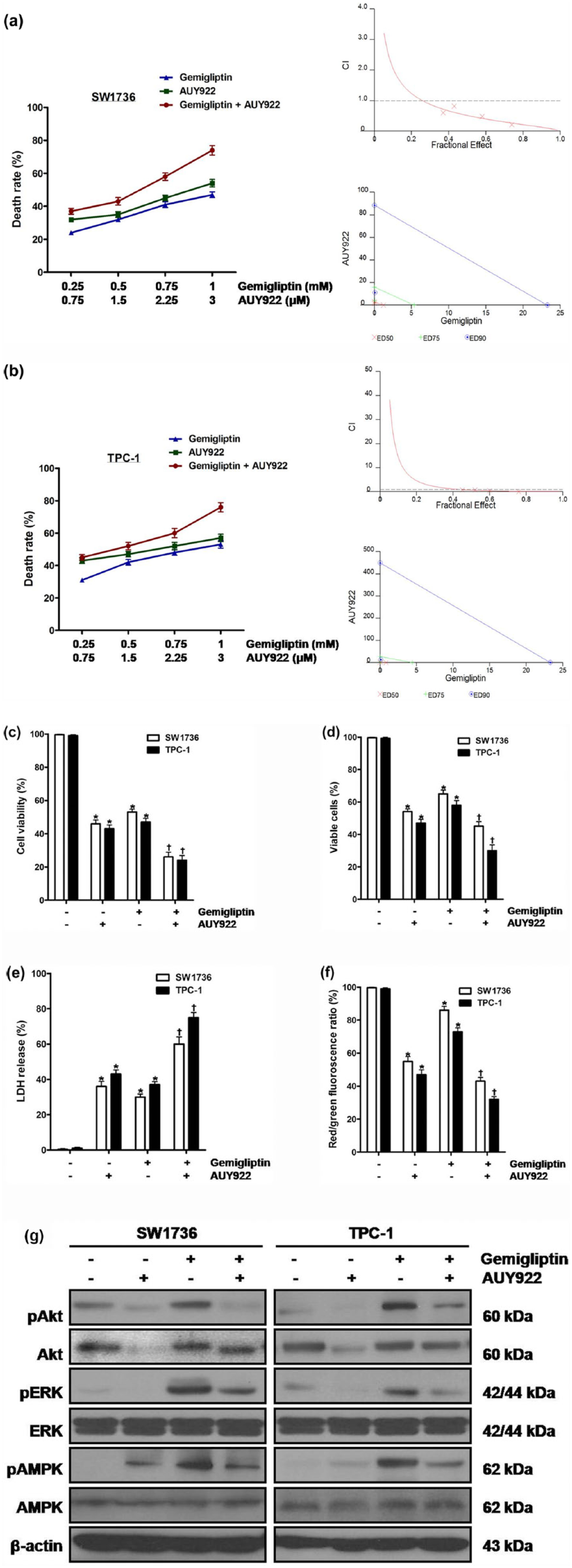
The influence of gemigliptin in combination with AUY922 on survival of thyroid carcinoma cells. (a) SW1736 and (b) TPC-1 cells were simultaneously treated with both gemigliptin at 0.25, 0.5, 0.75, and 1 mM and AUY922 at 0.75, 1.5, 2.25, and 3 μM for 48 h. Cell viability was measured using CCK-8 assay, and death rate was calculated as 100-cell viability (%). Combination index (CI) and isobologram were calculated. The horizontal dash lines at CI = 1.0 are drawn. (c–g) SW1736 and TPC-1 cells were simultaneously treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, and (c) cell viability, (d) the percentage of viable cells, (e) cytotoxic activity, (f) mitochondrial membrane potential, and (g) the total and phospho-protein levels of Akt, ERK1/2, and AMPK were measured. All experiments were performed in triplicate. The blots are representative of independent experiments. Data are expressed as mean ± SE (*p < 0.05 vs each matched control and †p < 0.05 vs cells treated with AUY922 alone).
CI values at combined doses determined by the median effect analysis method in thyroid carcinoma cells.

CI: combination index; GEM: gemigliptin.
CI values less than 1.0, equal to 1.0, and greater than 1.0 indicate synergism, additivity, and antagonism, respectively.
Next, to confirm synergistic activity of gemigliptin with AUY922 in induction of cell death, cells were simultaneously treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, and then cell viability (Figure 3(c)), the percentage of viable cells (Figure 3(d)), cytotoxic activity (Figure 3(e)), and mitochondrial membrane potential (Figure 3(f)) were measured. After cotreatment of gemigliptin and AUY922, compared with treatment of AUY922 alone, cell viability, the percentage of viable cells, and mitochondrial membrane potential were diminished and cytotoxic activity was enhanced.
In regard to the role of Akt, ERK1/2, and AMPK in synergistic cytotoxicity of gemigliptin with AUY922, cells were treated with AUY922 at 0.5, 1, and 3 μM for 48 h, after which hsp90 protein levels, and the total and phospho-protein levels of Akt, ERK1/2, and AMPK were measured (Supplementary Figure 2). In AUY922-treated cells, the protein levels of hsp90 and phospho-AMPK were enhanced and those of total and phospho-Akt, and phospho-ERK1/2 were diminished without change in those of total ERK1/2 and total AMPK. When the cells were simultaneously treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, compared with AUY922 alone, the protein levels of total and phospho-Akt, phospho-ERK1/2, and phospho-AMPK were enhanced without alteration in those of total ERK1/2 and total AMPK (Figure 3(g)).
Synergistic activity of gemigliptin with AUY922 in induction of cell death is involved in Bcl2 family proteins in thyroid carcinoma cells
To document the mode of cell death in the synergism between gemigliptin and AUY922-induced cytotoxicity in thyroid carcinoma cells, cells were simultaneously treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, and the percentage of apoptotic cells was measured using FACS analysis (Figure 4(a)–(c)). Under cotreatment of gemigliptin and AUY922, compared with treatment of AUY922 alone, the percentage of apoptotic cells was elevated.

The mode of cell death and the involvement of Bcl2 family proteins in combination of gemigliptin with AUY922 in thyroid carcinoma cells. (a) SW1736 and (b) TPC-1 cells were simultaneously treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, and apoptotic cells were measured using FACS analysis, and (c) the percentage of apoptotic cells was calculated. (d) SW1736 and TPC-1 cells were treated with gemigliptin at 0.25, 0.5, and 1 mM for 48 h and at 1 mM for 24 and 48 h, and the protein levels of Bcl2 and Bax were measured. (e) SW1736 and TPC-1 cells were treated with AUY922 at 0.5, 1, and 3 μM for 48 h and at 3 μM for 24 and 48 h, and the protein levels of Bcl2 and Bax were measured. (f) SW1736 and TPC-1 cells were treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, and the protein levels of Bcl2 and Bax were measured, quantified by densitometry, and normalized to β-actin levels. (g) The relative levels of protein to β-actin were calculated. All experiments were performed in triplicate. The blots are representative of independent experiments. Data are expressed as mean ± SE (*p < 0.05 vs each matched control and †p < 0.05 vs cells treated with AUY922 alone).
To determine the involvement of Bcl2 family proteins in synergistic activity of gemigliptin with AUY922 in induction of death of thyroid carcinoma cells, cells were treated with gemigliptin at 0.25, 0.5, and 1 mM for 48 h and at 1 mM for 24 and 48 h, and then the protein levels of Bcl2 and Bax were measured (Figure 4(d)). As a result of treatment, Bcl2 protein levels were elevated, whereas Bax protein levels were not changed. When cells were treated with AUY922 at 0.5, 1, and 3 μM for 48 h and at 3 μM for 24 and 48 h, Bax protein levels were elevated, and Bcl2 protein levels were reduced (Figure 4(e)). In cells treated with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, compared with AUY922 alone, the protein levels of Bax and cleaved PARP were elevated, while Bcl2 protein levels were not altered (Figure 4(f)). As depicted in Figure 4(g), Bax/Bcl2 ratio was elevated after cotreatment of gemigliptin and AUY922, compared with treatment of AUY922 alone.
To recognize whether Bax takes part in synergistic cytotoxicity of gemigliptin with AUY922 in thyroid carcinoma cells, cells were transfected with Bax siRNA prior to simultaneous treatment with both gemigliptin at 1 mM and AUY922 at 3 μM for 48 h, after which cell viability, the percentage of viable cells, and cytotoxic activity were measured (Supplementary Figure 3). In Bax siRNA-transfected cells, compared with control siRNA-transfected cells, cell viability, the percentage of viable cells, and cytotoxic activity were not changed.
Discussion
To our best knowledge, this is the first study demonstrating that the DPP-IV inhibitor gemigliptin results in cell death in conjunction with alterations in Akt, ERK1/2, and AMPK in thyroid carcinoma cells. In addition, our results indicate that gemigliptin has a synergistic activity with the hsp90 inhibitor AUY922 in induction of cell death via regulation of Akt, ERK1/2, and AMPK and in involvement of Bcl2 family proteins in thyroid carcinoma cells.
DPP-IV inhibitor exerts divergent properties independent of its glucose-lowering actions. 27 With respect to the effect of DPP-IV inhibitor on malignant cells, sitagliptin and saxagliptin promote tumor metastasis with provocation of nuclear factor E2–related factor 2–mediated anti-oxidant response in various cancer cell lines and produce metastatic nodules in animal cancer models. 11 In contrast, it was shown that DPP-IV inhibitor was not relevant to increased risk of metastasis in type 2 diabetic patients with breast, prostate, and gastrointestinal cancers. 12 Although it was reported that gemigliptin repressed expression of vascular adhesion molecules and inflammatory cytokines through activation of Akt and AMPK in human vascular endothelial cells, 15 the influence of gemigliptin on survival of malignant cells including thyroid carcinoma cells has not been investigated. In this study, gemigliptin caused cell death in a time- and dose-dependent manner and increased phospho-protein levels of Akt, ERK1/2, and AMPK. When the inhibitors of Akt, ERK1/2, and AMPK were administered before treatment of gemigliptin, wortmannin led to cell death, whereas AZD6244 and compound C did not affect cell survival. Taken together, these results imply that gemigliptin induces cell death by modulating Akt but not ERK1/2 and AMPK in thyroid carcinoma cells. Furthermore, our findings suggest that suppression of Akt intensifies gemigliptin-induced death of thyroid carcinoma cells. Considering our previous data that inhibition of Akt augments cytotoxic effect of other chemotherapeutic agents on ATC cells,28–30 Akt inhibitor may be an adjuvant in thyroid carcinoma cells treated with gemigliptin. In this regard, further studies for the possibility of clinical use of gemigliptin in thyroid cancer patients, especially type 2 diabetic patients with thyroid cancer, will be needed.
Akt and ERK have fundamental roles in survival, migration, invasion, and epithelial-to-mesenchymal transition of thyroid carcinoma cells.31,32 Meanwhile, AMPK acts as a cellular energy sensor to maintain energy homeostasis and is overexpressed in PTC tissues compared with normal thyroid tissues.33,34 With respect to connection among Akt, ERK1/2, and AMPK, it was shown that Akt regulated AMPK activity in gemigliptin-treated human vascular endothelial cells, and ERK1/2 repressed AMPK activity in human leukemia cells.15,26 In this study, AZD6244 enhanced phospho-Akt protein levels, and wortmannin diminished phospho-AMPK protein levels in cells treated with gemigliptin. By contrast, the impact of AZD6244 on AMPK and wortmannin on ERK1/2 was cell specific. All taken together, these results imply that suppression of ERK1/2 is related to phosphorylation of Akt, and stimulation of Akt is relevant to phosphorylation of AMPK in thyroid carcinoma cells exposed to gemigliptin.
Hsp90 acts as a defender against proteotoxic stress and is overexpressed in cancer cells compared with normal cells. 35 Moreover, hsp90 is present in an activated superchaperone complex hypersensitive to hsp90 inhibitors in cancer cells, whereas it is uncomplexed and less sensitive to hsp90 inhibitors in normal cells. 19 Because stressful environment makes cancer cells dependant on hsp90, cytotoxicity of hsp90 inhibitors is highly potent in cancer cells compared with normal cells.36,37 Although the net effect of hsp90 inhibition in normal and cancer cells may be not always consistent, it is evident that therapeutic selectivity can be accomplished against cancer cells. However, combination of hsp90 inhibitors with other chemotherapeutic agents should be cautiously proposed since there is potential for antagonism as well as synergism and additivity.
AUY922 has a powerful anti-tumor activity and potentiates cytotoxicity of other chemotherapeutic agents in cancer cells.22–24 In our previous study, AUY922, compared with other hsp90 inhibitors, had a potent cytotoxic activity in ATC cells and exhibited synergistic cytotoxicity with the histone deacetylase inhibitor PXD101 in conjunction with the inactivation of PI3K/Akt and survivin and the activation of DNA damage response in ATC cells. 25 Thus, we hypothesized that gemigliptin would augment AUY922-induced cytotoxicity in thyroid carcinoma cells. In this study, when gemigliptin was combined with AUY922, all of the CI values in drug combination analysis were lower than 1.0, suggesting that gemigliptin synergizes with AUY922 in induction of death in thyroid carcinoma cells. In addition, gemigliptin in combination with AUY922, compared with AUY922 alone, accelerated cytotoxicity, based on the data of cell viability, the percentage of viable cells, cytotoxic activity, and mitochondrial membrane potential, providing additional evidences for the synergism. Intriguingly, cotreatment of gemigliptin and AUY922, compared with treatment of AUY922 alone, elevated the percentage of apoptotic cells, implying that apoptotic cell death contributes to synergistic cytotoxicity in combination of gemigliptin with AUY922 in thyroid carcinoma cells. Furthermore, gemigliptin in combination with AUY922, compared with AUY922 alone, elevated the protein levels of total and phospho-Akt, phospho-ERK1/2, and phospho-AMPK without change in those of total ERK1/2 and total AMPK. Taken collectively, these results suggest that synergistic activity of gemigliptin with AUY922 is accompanied by alterations in Akt, ERK1/2, and AMPK in induction of cytotoxicity in thyroid carcinoma cells, and gemigliptin combined with AUY922 may be an attractive therapeutic option in human thyroid cancer.
Bcl2 family proteins engage in various cellular processes, especially survival. 38 In this regard, relative expression of the anti-survival protein Bax and the pro-survival protein Bcl2 is closely associated with survival of cancer cells.39–41 In this study, gemigliptin increased Bcl2 protein levels, while it did not change Bax protein levels. In contrast, AUY922 decreased Bcl2 protein levels and increased Bax protein levels. When cotreatment of gemigliptin and AUY922, compared with treatment of AUY922 alone, was done, the protein levels of Bax and cleaved PARP increased, whereas Bcl2 protein levels were not altered resulting in increment of Bax/Bcl2 ratio. Under condition of Bax inhibition by siRNA, cell viability, the percentage of viable cells, and cytotoxic activity were not varied. In summary, these results imply that gemigliptin has synergistic cytotoxicity with AUY922 via involvement of Bcl2 family proteins in thyroid carcinoma cells.
In conclusion, our results suggest that gemigliptin induces cytotoxicity with concomitant changes in expression of Akt, ERK1/2, and AMPK in thyroid carcinoma cells. Moreover, gemigliptin synergizes with AUY922 in induction of cytotoxicity through modulation of Akt, ERK1/2, and AMPK as well as participation of Bcl2 family proteins in thyroid carcinoma cells. This study will provide the possibility of clinical implications of gemigliptin alone or in combination with hsp90 inhibitors as a therapeutic regimen in type 2 diabetic patients with thyroid cancer refractory to conventional chemotherapeutic agents.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. 2015R1A2A2A01003589) to S.J.L., Republic of Korea.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.