
Abstract
Hepatocellular carcinoma has the second highest incidence rate among malignant cancers in China. Hepatocellular carcinoma development is complex because of the metabolism disequilibrium involving SULT1A3/4, a predominant sulfotransferase that metabolizes sulfonic xenobiotics and endogenous catecholamines. However, the correlation between SULT1A3/4 and hepatocellular carcinoma progression is unclear. By utilizing immunofluorescence and immunohistochemical analysis, we found that in nine hepatocellular carcinoma clinical specimens, SULT1A3/4 was abundantly expressed in tumor tissues compared to that in the adjacent tissues. Moreover, liver cancer cells (HepG2, MHCC97-L, and MHCC97-H) had higher basal expression of SULT1A3/4 than immortalized liver cells (L02 and Chang liver). Ultra-high-pressure liquid chromatography–tandem mass spectrometry assay results further revealed that the concentration of dopamine (a substrate of SULT1A3/4) was negatively correlated with SULT1A3/4 protein expression. As a transcriptional regulator of SULT1A3/4 in turn, dopamine was used to induce SULT1A3/4 in vitro. Interestingly, dopamine significantly induced SULT1A3/4 expression in liver cancer HepG2 cells, while decreased that in L02 cells. More importantly, the expression levels of epithelial–mesenchymal transition biomarkers (N-cadherin and vimentin) and cell stemness biomarkers (nanog, sox2, and oct3/4) considerably increased in HepG2 with dopamine-induced SULT1A3/4, whereas in L02, epithelial–mesenchymal transition and cancer stem cell–associated proteins were contrarily decreased. Furthermore, invasion and migration assays further revealed that dopamine-induced SULT1A3/4 dramatically stimulated the metastatic capacity of HepG2 cells. Our results implied that SULT1A3/4 exhibited bidirectional effect on tumor and normal hepatocytes and may thus provide a novel strategy for hepatocellular carcinoma clinical targeting. In addition, SULT1A3/4 re-expression could serve as a biomarker for hepatocellular carcinoma prognosis.
Introduction
Associated with the pathogenesis of hepatocellular carcinoma (HCC), metabolic homeostasis disrupted into chaos involving numbers of metabolism enzymes’ dysfunction, which conversely promoted carcinogenesis. 1 According to recent reports, the function and polymorphisms of metabolic enzymes such as glutathione S-transferases (GSTs) and sulfotransferases (SULTs) might increase HCC susceptibility, especially for Chinese population even in the absence of hepatitis B virus (HBV) infection. 2 , 3
SULTs are predominant phase-II enzymes that conjugate sulfonic xenobiotics and facilitate their elimination. A recent study revealed that disorder of SULT1A1, accounting for 53% of SULTs in liver, 4 was closely related to HCC progression in smoking patients. 3 In contrast with the abundant expression of SULT1A1 in hepatocytes, SULT1A3/4 was absolutely absent in adult human liver. 5 Similar to α-fetoprotein (AFP, the standard biomarker for HCC diagnosis 6 ), SULT1A3/4 was highly expressed in fetal liver5 and tumor tissues, 7 suggesting that the expression of SULT1A3/4 might reflect stemness, versatility, and malignancy in tumor development. Somehow the role of SULT1A3/4 in HCC development is unclear. Moreover, SULT1A3/4 also played key role in maintaining the homeostasis of monoamine neurotransmitters. 5 Dopamine (DA), main neurotransmitter existed in gastrointestine and liver, 8 , 9 was a substrate for SULT1A3/4, while in turn was also a regulator responsible for SULT1A3/4 transcriptional activation. 9 Although many studies demonstrated that endogenous DA secretion was negatively correlated with various cancer progressions, such as HCC, 10 , 11 breast cancer, 12 and gastric cancer, 13 whether the tumor suppression effects of DA was associated with SULT1A3/4 remains unknown.
Among all HCC patients, nearly 90% of the HCC-related deaths were ascribed to metastasis. 14 During metastasis, single tumor cells disseminated from the primary tumor underwent epithelial–mesenchymal transition (EMT) and then squeezed into the blood circulation; after extravasated and reversed the EMT to mesenchymal–epithelial transition (MET) transition, they ultimately outgrown as macroscopic metastasis. 15 Thus, EMT and percentage of cancer stem cells (CSCs), pluripotent cells attributed to tumor malignancy, were the prerequisites for tumor metastasis and recurrence. 16 The alteration of various epithelial and mesenchymal markers such as vimentin, N-cadherin, and E-cadherin were closely correlated with tumor progression. 17 The biomarkers of CSCs, such as oct3/4, nanog, and sox2, were also recommended as reference for determining the potential of tumor cell migration and invasion. 18 , 19 The progression of HCC accompanied by metabolic disequilibrium. However, whether metabolism enzymes played roles in EMT transition and CSCs acquisition, which ultimately promoted tumor metastasis, was still unknown.
In this study, nine tumor and adjacent tissues were collected from clinical HCC patients who underwent partial hepatectomy. By utilizing immunohistochemical and immunofluorescence assays, we found that SULT1A3/4 in situ was re-expressed in HCC tissues compared to that in adjacent tissues. Furthermore, by stimulating SULT1A3/4 expression via DA treatment, we found that the DA-induced SULT1A3/4 in HepG2 cells significantly promoted tumor migration and invasion as well as increased the biomarkers of EMT and CSCs. By contrast, in L02 cells, DA could suppress SULT1A3/4 and subsequently decrease EMT and CSC-associated proteins. Our results demonstrated that SULT1A3/4 exhibited a bidirectional effect on HepG2 and L02, which might provide a novel strategy for HCC clinical targeting. In addition, the re-expression of SULT1A3/4 could be a biomarker for HCC prognosis.
Materials and methods
Cell lines
Human liver cancer cell lines (MHCC97-L and MHCC97-H) were purchased from the Liver Cancer Institute of Fudan University. Human liver cancer cell line HepG2 and human immortalized liver cell lines (L02 and Chang liver 20 ) were purchased from the Institute of Biochemistry and Cell Biology (Shanghai, China). L02, HepG2, MHCC97-L, and MHCC97-H cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% (v/v) fetal bovine serum (FBS; Gibco, Invitrogen, Carlsbad, CA, USA). Chang liver cells were cultured in RPMI 1640 medium supplemented with 10% (v/v) FBS. All cells were grown in a humidified atmosphere of 5% CO2 and 95% air at 37°C.
Tissue samples
Nine of the HCC tumors and the adjacent tissues were collected from Department of Nan Fang Hospital, affiliated to Southern Medical University between March 2012 and December 2014. All the tissue specimens were collected from HBV-positive HCC patients who had HBV infection for over 5 years, and all tissues were made as paraffin sections. All procedures had been approved by the Institute Research Ethics Committee and patient consent. All samples had been histologically confirmed as liver cancer using World Health Organization (WHO) criteria as part of the routine histopathology work-up by two independent pathologists. The clinical diagnosis was also conducted by computed tomography (CT), color Doppler ultrasonography (CDUS), and magnetic resonance imaging (MRI).
Statistical analysis
All the assays were performed in three independent experiments. All results were presented as mean ± standard deviation (SD). All the accompanying bar graphs represent the data from those donors showed in the figures. Significant differences were analyzed using Student’s t-test (two groups) or one-way analysis of variance (ANOVA) followed by the least significant difference (LSD) post hoc test (for more than two groups) by SPSS Statistics (SPSS 19.0; IBM, New York, USA); *p < 0.05, **p < 0.01, and ***p < 0.001 are determined as significance.
Results
SULT1A3/4 was abundantly expressed in HCC tissues
All the tissues were analyzed by histological methods and then classified into II–IIIA stages (Table 1). Hematoxylin–eosin (HE) staining, immunohistochemical, and immunofluorescence assays were conducted in tumor and adjacent tissues to determine the basal expression of SULT1A3/4 in the HCC tissues. The HE staining results revealed that the tumor tissues were more compact and fibrocystic than the adjacent tissue. Many tumor cells combined and formed lobular structures (Figure 1(a)). The results of immunohistochemical assay showed that SULT1A3/4 was overexpressed in cancer tissues compared with that in adjacent tissues. The SULT1A3/4 expression levels were 788.73% ± 55.07% and 100% in HCC tissues and adjacent tissues, respectively (p < 0.01; Figure 1(b)). The results of the immunofluorescence assays showed that the percentages of total SULT1A3/4 fluorescence in HCC tissues and adjacent tissues were 467.2% ± 30.1% and 101.3% ± 10.3%, respectively (p < 0.01; Figure 1(c)). Also, the effect of DA on cell viabilities was determined both in immortalized liver cells (L02 and Chang liver) and liver cancer cells (HepG2, MHCC97-L, and MHCC97-H; Supplement Figure 5).
The patient information of liver cancer cases.

TNM: tumor–node–metastasis; HBV: hepatitis B virus; HBsAg: hepatitis B surface antigen; AFP: alpha-fetoprotein.
II: T2N0M0; IIIa: T3aN0M0 (T2: either a single tumor (any size) that has grown into blood vessels or more than one tumor where no tumor is larger than 5 cm (about 2 in) across; T3a: more than one tumor, with at least one tumor larger than 5 cm across; N0: the cancer has not spread to the regional lymph nodes; M0: the cancer has not spread to distant lymph nodes or other organs).
Median with range.
Clinical classification of tumors was performed using the tumor metastasis (TNM) classification system of the American Joint Committee on Cancer (AJCC) and the Union for International Cancer Control (UICC; 7th edition).
Positive number (median ± standard deviation).

Protein expression of SULT1A3/4 in tissue samples. (a) HE staining. (b) Immunohistochemistry (bottom: accompanying bar graphs represent the data of HE staining). (c) Immunofluorescence (right: accompanying bar graphs represent the data of immunofluorescence). Triplicate assays were performed for each analysis. All the accompanying bar graphs represent the data from those donors shown in the figure. Data are represented as mean ± SD (**p < 0.01 compared with the adjacent tissues).
SULT1A3/4 protein expression was negatively correlated with DA concentration in HCC tissues
Western blot, real-time polymerase chain reaction (PCR), and liquid chromatography–tandem-mass spectrometry (LC/MS/MS) analyses were utilized to determine the relationship between of SULT1A3/4 and DA in tumor and adjacent tissues. Compared with adjacent tissues, the protein expression of SULT1A3/4 in tumor tissues was significantly increased, especially in No. 4, 5, and 8 HCC patients with the respective induction of 1076.53% ± 126.32%, 328.24% ± 92.9%, and 361.14% ± 50.98% (p < 0.01; Figure 2(a)). However, the gene expression of SULT1A3/4 was decreased in seven of the nine tumor tissues compared with that in the corresponding adjacent tissues (p < 0.01; Figure 2(b)). Meanwhile, the DA concentrations were also considerably lower than those in the adjacent tissues, and most reduction was observed in the tumor tissues of the No. 5 patient (17.45% ± 1.19%; p < 0.01; Figure 2(c)). To determine the correlation of SULT1A3/4 protein and DA concentration, pairwise analysis was performed. DA levels in tissue specimens were remarkably negative related to SULT1A3/4 protein level (r = –0.885 in adjacent tissues; r = –0.814 in tumor tissues; Figure 2(d) and (e)). At the same time, we further determined the concentration of intercellular residual DA in L02 and HepG2 cells with or without DA treatment and found DA induced SULT1A3/4 upregulation in HepG2 cells, which is consistent with our previous observation in tumor tissues. While similar to the adjacent normal tissues, in immortalized liver cell (L02), DA-suppressed SULT1A3/4 attenuated DA metabolism, which consequently increased residual DA concentration in cells (Supplement Figure 1). We also found that SULT1A3/4 metabolic activity is closely related with its protein level (Supplement Figure 2, correlation: r2 = 0.978, p = 0.001).

Protein expression of SULT1A3/4 was negatively correlated with DA concentration in tissue samples. (a) Western blot assay (bottom: accompanying bar graphs represent the data of western blot band). (b) RT-PCR assay. (c) UHPLC and LC-MS/MS assays. (d and e) Presentation of pairwise correlation between SULT1A3/4 and DA. Triplicate assays were performed for each analysis. All the accompanying bar graphs represent the data from those donors shown in the figure. Data are represented as mean ± SD (n = 9; *p < 0.05, **p < 0.01, and ***p < 0.001 compared with the adjacent tissues).
SULT1A3/4 was also overexpressed in the liver cancer cells
To determine the appropriate cell lines for further mechanism analysis, immunofluorescence assay, western blot, and real-time PCR were performed in L02, Chang liver, HepG2, MHCC97-L, and MHCC97-H. Compared to L02 and Chang liver, the fluorescence of SULT1A3/4 was extremely enhanced in HepG2, MHCC97-L, and MHCC97-H. Abundant SULT1A3/4 protein was detected in HepG2 compared with that in other two cancer cell lines (MHCC97-L and MHCC97-H; p < 0.05) (Figure 3(a)). Results showed that the SULT1A3/4 protein expression levels increased by 20.39 ± 3.62% in HepG2 cells (p < 0.01; Figure 3(b)). The gene expression of SULT1A3/4 was also upregulated in the cancer cells in contrast to that in L02 cells (p < 0.01; Figure 3(c)), while in SULT1A3/4-suppressed L02 cells, the intercellular DA concentrations were contrarily increased in a DA dose–dependent manner (Revised Supplement Figure 6).
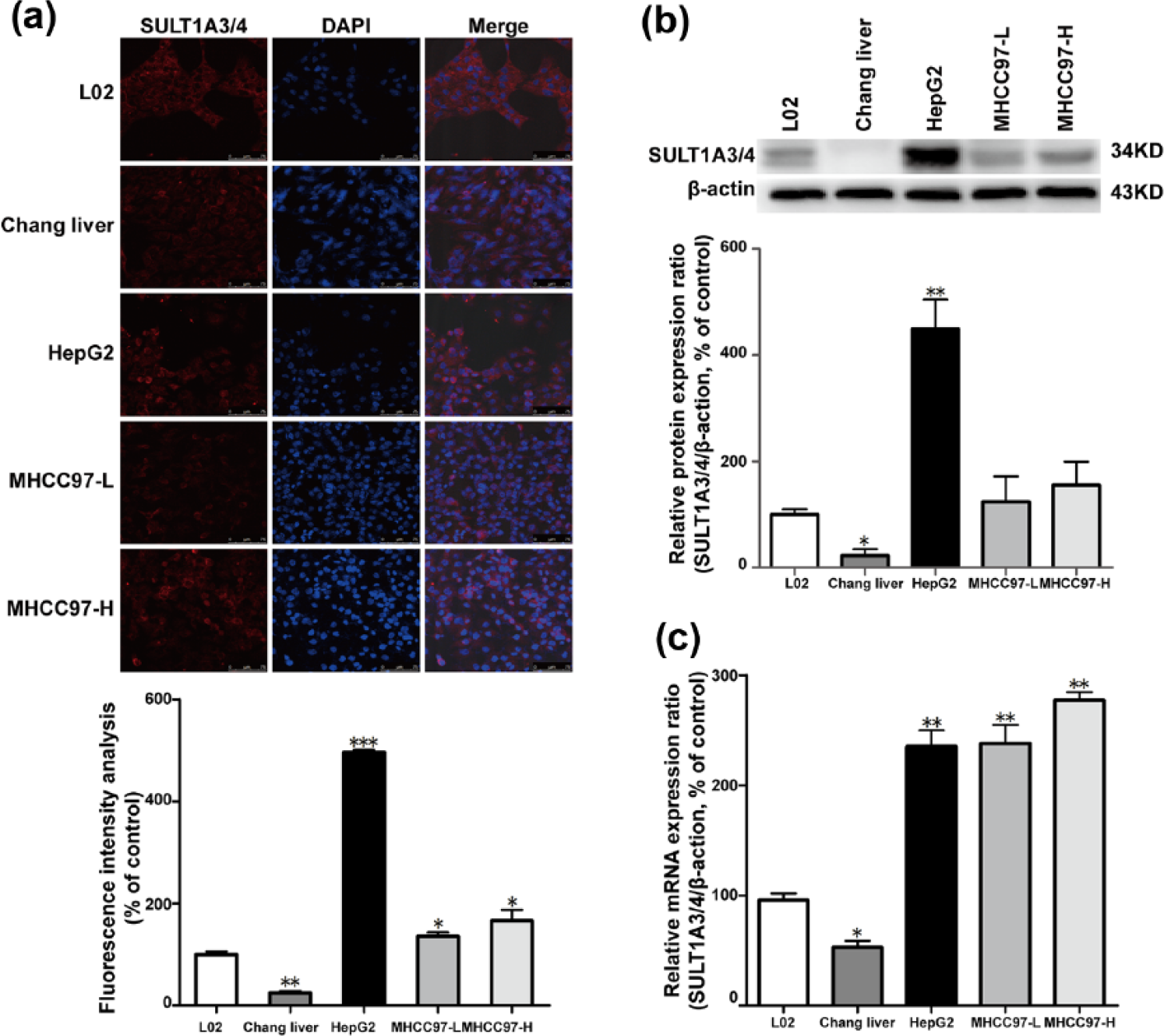
Expression of SULT1A3/4 in liver cancer cells. (a) Immunofluorescence (bottom: accompanying bar graphs represent the data of total SULT1A3/4 fluorescence). (b) Western blot assay (bottom: accompanying bar graphs represent the data of western blot band). Triplicate assays were performed for each analysis. All the accompanying bar graphs represent the data from those donors shown in the figure. Data are represented as mean ± SD (*p < 0.05 and **p < 0.01 compared with L02). (c) RT-PCR assay.
DA-induced SULT1A3/4 promoted the activation of CSCs in liver cancer cells
With DA (20 µM) treated, the fluorescence of SULT1A3/4 in HepG2 notably increased by 160.57% ± 4.12%. And the SULT1A3/4 fluorescence in L02 decreased by 59.43 ± 3.55% (p < 0.01; Figure 4(a)). The accompanying bar graphs were not shown in the article. In HepG2, the DA dose-dependently increased the SULT1A3/4 protein expression levels by 104.62% ± 3.54% and 167.08 ± 7.71% at DA concentrations of 10 and 20 µM, respectively (p < 0.01; Figure 4(b)). The SULT1A3/4 protein expression in L02 was significantly suppressed after the DA treatment (p < 0.01; Figure 4(b)). Moreover, the CSC biomarkers, such as nanog, sox2, and oct3/4, exhibited different protein expression levels in L02 and HepG2. Results showed that the protein expression levels of nanog, sox2, and oct3/4 in HepG2 were considerably enhanced in a dose-dependent manner and that in L02 was substantially decreased (p < 0.01; Figure 4(c)).

DA-induced SULT1A3/4 promotes CSC activation. (a) Immunofluorescence. (b) Western blot assay (bottom: accompanying bar graphs represent the data of western blot band). Triplicate assays were performed. All the accompanying bar graphs represent the data from those donors shown in the figure. Data are represented as mean ± SD (**p < 0.01 compared with control). (c) Western blot assay (right: accompanying bar graphs represent the data of western blot band).
DA-induced SULT1A3/4 promoted EMT of liver cancer cells
Compared with the control, the DA-induced SULT1A3/4 considerably enhanced the mesenchymal-associated markers (vimentin and N-cadherin) by 138.19% ± 7.02% and 179.18% ± 11.64% in HepG2, whereas it decreased the epithelial-associated protein E-cadherin by 45.13% ± 1.4% (p < 0.01; Figure 5(a)). By contrast, the DA-suppressed SULT1A3/4 decreased the vimentin and N-cadherin levels by 66.77% ± 1.76% and 27.21% ± 2.51%, respectively, whereas it increased the E-cadherin level by 546.14% ± 26.26% in L02 (p < 0.01; Figure 5(a)). Furthermore, DA dose-dependently stimulated the migration ratio of HepG2 with inductions of 139.17% ± 6.90% and 223.03% ± 27.54% at DA concentrations of 10 and 20 µM, respectively (p < 0.05; Figure 5(b)). Similarly, the invasion ratio of HepG2 was enhanced by 183.92% ± 2.29% and 334.07% ± 31.45% at DA concentrations of 10 and 20 µM, respectively (p < 0.01; Figure 5(c)). To further address the distinct role of DA in immortalized normal cells (L02) and liver cancer cells (HepG2), various SULT1A3/4 transcription–related factors such as DA D1 receptor, N-methyl-

DA-induced SULT1A3/4 promotes EMT. (a) Western blot assay (bottom: accompanying bar graphs represent the data of western blot band. (b and c) Wound-healing and Transwell invasion assays. Triplicate assays were performed. All the accompanying bar graphs represent the data in the figure. Data are represented as mean ± SD (*p < 0.05 and **p < 0.01 compared with control).
Discussion
There are some tumor biomarkers in serum for HCC monitoring and early diagnosis, 21 such as AFP and carcino-embryonic antigen (CEA). However, several shortcomings were inevitable, such as poor specificity and instability, at certain time points during inspection. Therefore, novel tumor-specific biomarkers remain extensively explored.
Similar to the role of AFP in fetal development, SULT1A3/4, a catecholamine-metabolizing enzyme, was also highly expressed only in fetal liver, but diminishes after birth. 22 However, alterations in SULT1A3/4 during tumor progression remain unclear. To determine the basal expression of SULT1A3/4 in HCC patients, nine clinical specimens were collected and subjected to immunohistological analysis. Results indicated that in both the HCC tumor tissues and liver cancer cells (HepG2, MHCC97-L, and MHCC97-H), the SULT1A3/4 protein expression was abundant compared to that in adjacent tissues, L02, and Chang liver (Figures 1(b), (c), 3(a) and (b)). The difference between the SULT1A3/4 basal expression levels of adjacent and tumor tissues suggested that the SULT1A3/4 acted not only as a tumor-specific target for further drug discovery but also as a tumor sensor for HCC monitoring.
The other important function of SULT1A3/4 in vivo was the modulation of neurotransmitter homeostasis by metabolizing catecholamines, such as DA and isoprenaline. 23 DA is highly distributed in the brain, 24 gastrointestinal tract,10 and liver cells. 25 By performing ultra-high-pressure liquid chromatography–tandem mass spectrometry (UHPLC/MS/MS), we found that endogenous DA levels in HCC tissues were extremely lower than those in adjacent tissues (Figure 2(c)). Results suggested that the metabolic activity of SULT1A3/4 recovered in HCC tissues (Figure 2(a)). Furthermore, the DA level was negatively correlated with SULT1A3/4 protein expression both in the tumor and adjacent tissues (Figure 2(d) and (e)). The gene expression levels of SULT1A3/4 were low in seven of the nine tumor tissues (Figure 2(b)). This result might be attributed to the weak correlation between the SULT1A3/4 protein and messenger RNA (mRNA) expression in adjacent tissues and the disrupted interaction between the SULT1A3/4 protein and mRNA expression in the tumor tissues because of DNA instability and mRNA dysfunction.
Substantial evidence demonstrated that DA was inversely correlated with various cancer progressions, such as liver cancer 25 and gastric cancer.10 Chronic stress conditions promote endogenous DA degeneration and subsequently form a conducive microenvironment for tumor growth. 8 , 26 In addition, DA could inhibit angiogenesis induced by vascular endothelial growth factors in solid tumors. 27 Although DA was a substrate of SULT1A3/4, it was a strong regulator of SULT1A3/4 expression. 28 According to our data, DA significantly induced SULT1A3/4 expression in tumor cells but reduces SULT1A3/4 expression in normal cells. The finding indicated that the response of SULT1A3/4 transcriptional activities to DA stimulation depended on cell types. We also found that DA-induced SULT1A3/4 considerably decreased the percentages of CSCs and EMT capacities in L02, whereas it promoted the EMT and CSCs in HepG2 (Figures 4(c) and 5). Results suggested that SULT1A3/4 was closely related with tumor metastasis and promoted EMT and CSCs in HCC.
Overall, our study demonstrated that DA-induced SULT1A3/4 not only promoted EMT, migration, and invasion but also stimulated the activation of CSCs in liver cancer cells. In L02, reactivated SULT1A3/4 effectively suppressed EMT and CSCs (Figure 6). Therefore, SULT1A3/4 might be a potential target for therapeutic strategy and potential biomarker of early diagnosis and monitoring of liver cancer.

Schematic illustration of dopamine-induced SULT1A3/4 in hepatocellular carcinogenesis.
Footnotes
Acknowledgements
The authors would like to express their gratitude to Gangjun Du (Pharmacy College of Henan University) for generous assistance. J.Z. and H.L. contributed equally to this manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
The authors declare that there is no study with human participants.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was mainly supported by Guangdong Provincial Natural Science Foundation (2015A030312012), the Science and Technology Development of Guangzhou City (201509010004), and Traditional Chinese Medicine Bureau of Guangdong Province (20151192).
Informed consent
Informed consent was obtained from all individual participants included in the study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.