
Abstract
β-glucans are one of the most abundant forms of polysaccharides known as biological response modifiers which influence host’s biological response and stimulate immune system. Accordingly, this study was initiated to evaluate irradiated β-glucan as a modulator for cellular signaling growth factors involved in the pathogenesis of hepatocellular carcinoma in rats. Hepatocellular carcinoma was induced with 20 mg diethylnitrosamine/kg BW. Rats received daily by gastric gavage 65 mg irradiated β-glucan/kg BW. It was found that treatment of rats with diethylnitrosamine induced hepatic injury and caused significant increase in liver injury markers with a concomitant significant increase in both hepatic oxidative and inflammatory indices: alpha-fetoprotein, interferon gamma, and interleukin 6 in comparison with normal and irradiated β-glucan–treated groups. Western immunoblotting showed a significant increase in the signaling growth factors: extracellular signal–regulated kinase 1 and phosphoinositide 3-kinase proteins in a diethylnitrosamine-treated group while both preventive and therapeutic irradiated β-glucan treatments recorded significant improvement versus diethylnitrosamine group via the modulation of growth factors that encounters hepatic toxicity. The transcript levels of vascular endothelial growth factor A and inducible nitric oxide synthase genes were significantly higher in the diethylnitrosamine-treated group in comparison with controls. Preventive and therapeutic treatments with irradiated β-glucan demonstrated that the transcript level of these genes was significantly decreased which demonstrates the protective effect of β-glucan. Histological investigations revealed that diethylnitrosamine treatment affects the hepatic architecture throughout the significant severe appearance of inflammatory cell infiltration in the portal area and congestion in the portal vein in association with severe degeneration and dysplasia in hepatocytes all over hepatic parenchyma. The severity of hepatic architecture changes was significantly decreased with both β-glucan therapeutic and preventive treatments. In conclusion, irradiated β-glucan modulated signal growth factors, vascular endothelial growth factor A, extracellular signal–regulated kinase 1, and phosphatidylinositol-3-kinase, which contributed to experimental hepatocarcinogenesis.
Keywords
Introduction
β-glucans are polysaccharides found as constituents in several types of plants and microorganisms, including oat, barley, mushroom, seaweed, some bacteria, and yeast. β-glucans from several sources are differential in their structure, conformation, and thus biological activity. Oat and barley β-glucans are principally linear with β-1,4 linkages; mushroom and fungus β-glucans have the β-1,3 backbone branched with short β(1,6)-linked side chains. 1 Accordingly, these structural differences could affect both the β-glucan extraction and the biological activity. 2 Depolymerization of β-glucan using ionizing radiation by the cleavage of the glycosidic bonds used simply as an effective method to produce low molecular weight β-glucans with high solubility and low viscosity, easily absorbed by intestinal mucosa without causing any notable changes in the functional-group status as compared with acidic or enzymatic hydrolysis.3,4 Most β-glucans pass to the proximal small intestine and some are caught by macrophages.5,6 The targeted immune cells of β-glucans include macrophages, neutrophils, monocytes, natural killer (NK) cells, and dendritic cells. 7 Consequently, β-glucans can modulate both innate and adaptive responses, and they can enhance phagocytosis.8,9
Hepatocellular carcinoma (HCC) represents 80%–90% of all liver cancers and is the third most common cause of cancer morbidity, resulting in almost 1 million deaths every year. 10 Most cases of HCC are secondary to either a viral hepatitis infection (hepatitis B or C) or cirrhosis. 11 Modulated apoptosis and cell proliferation lead to the multistep development and progression of a hyperplastic to the dysplastic nodule, early HCC, and advanced HCC. 12 The receptors for these growth factors are broadly divided into G protein–coupled receptors and protein kinases.13,14 On ligand binding, these receptors induce their intracellular molecules in a cascade fashion. Many of the growth factor receptors have tyrosine kinase activity such as the epidermal growth factor receptor (EGFR) and vascular endothelial growth factor receptor (VEGFR) (Figure 1). However, the mitogen-activated protein kinase (MAPK), and the extracellular signal–regulated kinase (ERK) are serine/threonine kinases. Activation of receptor kinases by their ligands activates signaling pathways with effects upon angiogenesis, proliferation, apoptosis, or survival of cells. These growth factor signaling pathways can be molecular targets for treatment of HCC as second-line therapy along with standard therapies.13,15–18 Therefore, the objective of this investigation was to evaluate irradiated β-glucan (IGLU) as a modulator for cellular signaling growth factors involved in the pathogenesis of HCC in rats.

Cellular signaling pathways implicated in the pathogenesis of hepatocellular carcinoma.
Materials and methods
Glucan preparation
Glucan was extracted from dry mushroom, Agaricus bisporus, obtained from Ploshia (Giza, Egypt). The dry mushroom was added to 0.1 mol/L of NaOH and stirred for 30 min at 60°C. The material was heated to 115°C at 8.5 psi for 45 min and then allowed to settle for 72 h. The sediment was suspended and washed with distilled water by centrifugation at 350 g for 20 min. The alkali-insoluble solids were combined with 0.1 mol/L acetic acid and heated to 85°C for 1 h and then allowed to settle at 38°C. The acid-insoluble solids were drawn off and centrifuged. The compacted solid material was mixed with 3% H2O2 and refrigerated for 3 h with periodic mixing. The material was centrifuged and the pellet was washed twice with distilled water followed by two washes with 100% acetone. The harvested solid material (glucan powder) was dispersed on drying trays and dried under vacuum at 38°C for 2 h in the presence of Ca2SO4, and dried overnight under vacuum at room temperature. 19
Gamma irradiation
Glucan powder was irradiated using Co-60 Gamma chamber 4000A at the National Center for Radiation Research and Technology, Atomic Energy Authority (Cairo, Egypt). β-glucan was exposed to a dose of 50 kGy at room temperature (25 ± 2°C). The source strength was approximately 11.1 PBq with a dose rate of 10 kGy/h. IGLU samples were stored at 4°C.
Animals
The present study was carried out using 30 adult male Swiss albino rats (200±20 g) obtained from Nile Company for Pharmaceuticals and Chemical Industries (Cairo, Egypt). The rats were naïve to diethylnitrosamine (DEN) and glucan. Rodents were maintained with free access to standard laboratory pellet chow and water ad libitum. Rats were kept in the laboratory under controlled conditions of temperature (27 ± 2°C) and humidity (60 ± 5%) with 12 h light/12 h dark cycles in well-ventilated cages. All the experimental procedures were carried out according to “Guide for the Care and Use of Laboratory Animals” (NIH publication No. 85–23, 1985).
Experimental design
After an acclimatization period of 7 days, rats were randomly allocated and divided into equally-sized five groups of six animals each.
Group I rats kept as controls.
Group II rats received daily intraperitoneal injection of DEN (20 mg/Kg BW) for 6 weeks. 20
Group III rats were orally administrated daily with IGLU (65 mg/Kg BW) for 6 weeks. 21
Group IV rats were administrated IGLU daily for 2 weeks then experimentally induced with DEN concomitant with IGLU for a month (preventive treatment).
Group V rats were experimentally induced daily with DEN for 6 weeks, then treated with IGLU for 6 weeks (therapeutic treatment).
At the end of experiment, all animals were sacrificed 24 h after the last treatment under urethane anesthesia. Blood samples were collected and serum was separated and stored at −80°C for subsequent analyses. At autopsy, livers were harvested, perfused with cold isotonic saline, dried carefully and stored at −80 °C.
Biological assays
Serum alanine aminotransferase (ALT) and gamma glutamyl transferase (GGT) activities were determined using kits purchased from Biodiagnostic (Giza, Egypt). Levels of alpha-fetoprotein (AFP), interferon gamma (IFN-γ), and interleukin 6 (IL-6) were determined in serum using the corresponding rat enzyme-linked immunosorbent assay (ELISA) kit following protocols provided by the manufacturers (R&D Systems; DRG, USA).
Western immunoblotting
Liver tissue proteins were extracted using TRIzol reagent (Invitrogen) according to Chomczynski 22 following protocol provided by the manufacturer (ThermoFisher). A volume of 20 µg of protein per lane was separated with 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene difluoride (PVDF) membranes. The membranes were incubated at room temperature for 2 h with blocking solution comprising 5% nonfat dried milk in 10 mM Tris-Cl pH 7.5, 100 mM NaCl, and 0.1% Tween 20, and then incubated overnight at 4°C with the primary antibodies toward ERK-1 and PI3K with β-actin as loading control. After washing three times in 10 mM Tris-Cl pH 7.5, 100 mM NaCl, and 0.1% Tween 20, the membranes were incubated with the secondary monoclonal antibodies conjugated to horseradish peroxidase at room temperature for 2 h, and then membranes were washed four times with the same washing buffer. Membranes were developed and visualized by chemiluminescence using Amersham detection kit according to the manufacturer’s protocols and then exposed to X-ray film. Primary and secondary antibodies were purchased from Cell Signaling Technology (USA). Quantification of ERK-1 and PI3K proteins was performed by densitometric of the autoradiograms using a scanning laser densitometer analysis (Biomed Instrument Inc., USA). Results were expressed as arbitrary units after normalization for β-actin.
Detection of inducible nitric oxide synthase and vascular endothelial growth factor A relative gene expression in liver tissues
RNA isolation and reverse transcription
To investigate the changes in messenger RNA (mRNA) expression for growth factors, vascular endothelial growth factor A (VEGF-A) and inducible nitric oxide synthase (iNOS), total RNA were isolated from 100 mg of liver using TRIzol reagent (Invitrogen) according to Chomczynski22 in accordance with the manufacturer’s instructions (ThermoFisher). RNA integrity was confirmed by 1% agarose gel electrophoresis and stained with ethidium bromide. The first strand complementary DNA (cDNA) synthesis was performed with reverse transcriptase (Invitrogen) according to the manufacturer’s protocol using 1 µg of total RNA as the template.
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reactions (RT-PCRs) were performed in a Thermal Cycler Step One Plus (Applied Biosystems, USA) using Sequence Detection Software (PE Biosystems, USA). The oligonucleotides utilized in these experiments are listed in Table 1 according to gene bank accession numbers: NM_001287114.1 for VEGF-A, NM_012611.3 for iNOS, and NM_031144.3 for β-actin. A reaction mixture of total volume of 25 µL consisting of 2× SYBR Green PCR Master Mix (Applied Biosystems), 900 nM of each primer, and 2 µL of cDNA was prepared. The PCR thermal-cycling conditions included an initial step at 95°C for 5 min, 40 cycles at 95°C for 20 s, 60°C for 30 s, and 72°C for 20 s. Curve analysis was performed at the end of the reaction. The data were normalized using the β-actin gene that was amplified in each set of PCR experiments. Relative expression of target mRNA was calculated using the comparative Ct method using the Pfaffl method. 23
List of primers used in RT-PCR.

Histological examination
Liver samples were taken from rats in different groups and fixed in 10% formalin for 48 h. Tissues were washed by water then serial dilutions of alcohol (methyl-, ethyl-, and absolute ethyl) were used for dehydration. Specimens were cleared in xylene and embedded in paraffin beeswax at 56°C in hot air oven for 24 h. Paraffin tissue blocks of 4 µm thickness were cut by sledge microtome. The obtained tissue sections were collected on glass slides, deparaffinized, and stained by hematoxylin and eosin (H&E) stain for routine examination with microscope. 24
Statistical analyses
All data were expressed as the mean ± standard deviation (SD). One-way analysis of variance (ANOVA) with least significant difference (LSD) was used to test for differences in means of variables between groups. A probability of p < 0.05 was considered to be statistically significant. All data were analyzed by Statistical Package for Social Science (SPSS) version 13 for Windows (SPSS, USA) software program.
Results
Effect of IGLU on DEN-induced liver injury and inflammatory disturbance
It was found that treatment of animals with DEN induced hepatic injury throughout the significant increase in plasma ALT and GGT activities along with increments in AFP levels accompanied by significant increase in the inflammatory cytokines: IFN-γ and IL-6 in comparison with both controls and IGLU-treated groups as shown in Figure 2. Preventive and therapeutic administration of IGLU showed statistically significant improvements versus the DEN group throughout the exhibited significant modulation in studied parameters that augment the hepatic toxicity (Figure 2).
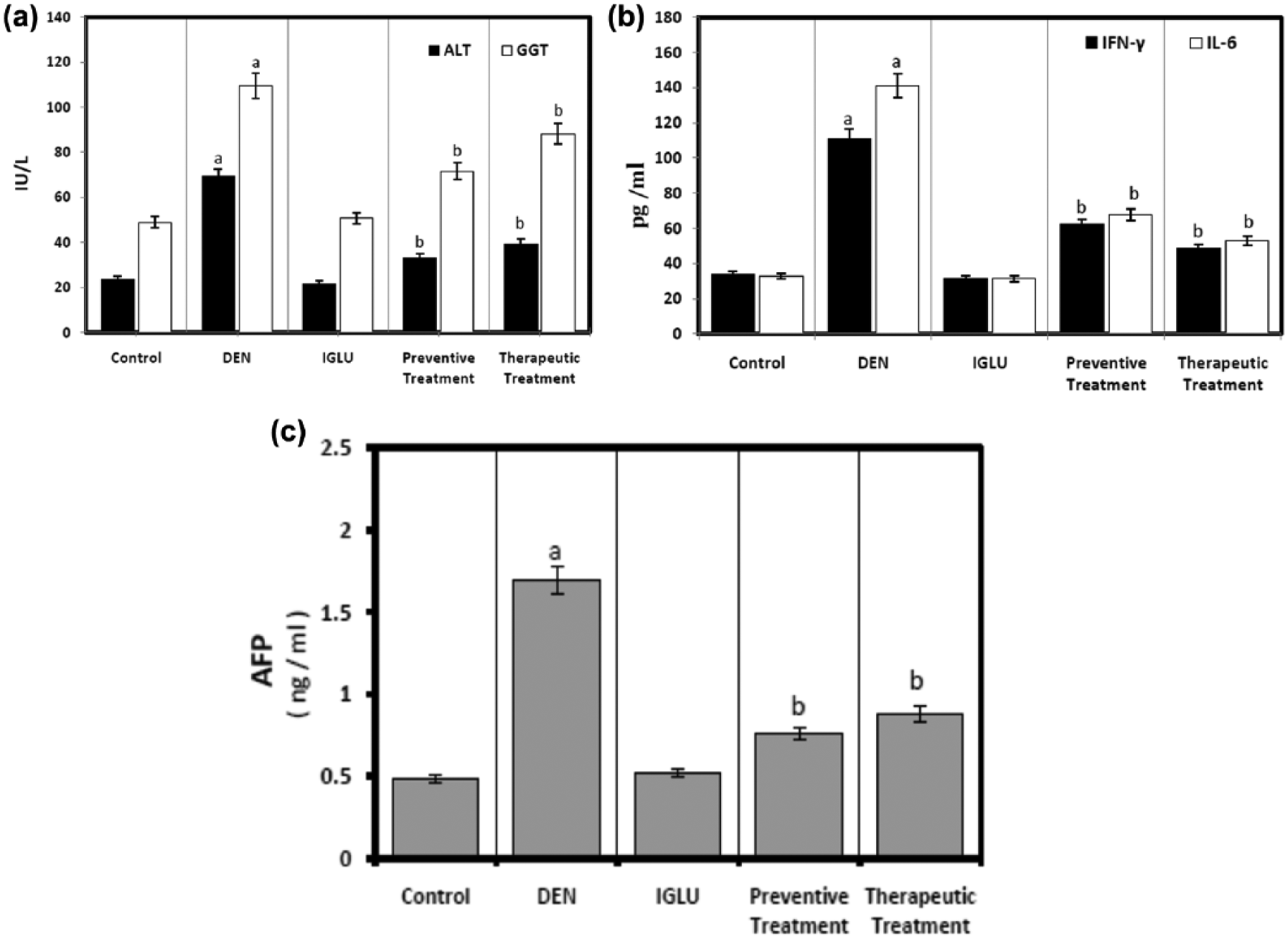
Effect of DEN upon serum liver function enzymes, inflammatory markers, and AFP in different treatments compared with controls. Each column represent mean ± SD (n = 6; ap < 0.05 compared to control; bp < 0.05 compared to DEN group).
Effect of IGLU on DEN-induced ERK-1 and PI3K signaling
DEN treatment induced markedly significant higher levels of the signaling growth factors: ERK-1 and PI3K proteins. IGLU preventive and therapeutic administrations showed statistically significant improvement versus the DEN group (Figure 3). These data suggest that the ERK-1 and PI3K signaling pathways are essential for the DEN-induced hepatocarcinogenesis.

Western blotting analysis of signaling growth factors: ERK-1 and PI3K protein expression in treated rats. Each bar represent mean ± SD for 6 animals in each group. (ap < 0.05 compared to control; bp < 0.05 compared to DEN group).
Effect of IGLU on DEN-induced alteration in mRNA gene expression
Expression of VEGF-A and iNOS genes was analyzed by RT-PCR. The transcript levels of VEGF-A and iNOS genes were significantly higher in DEN-treated group (Figure 4) in comparison with controls. Interestingly, after treatment with preventive and therapeutic IGLU, the transcript level of these genes was significantly lower than that in the DEN-treated animals.

Effect of IGLU upon DEN-induced alteration in mRNA expression of VEGF-A and iNOS expressed as relative quantification compared to the control group using quantitative RT-PCR analysis. Each bar represents mean ± SD for 6 animals in each group. (ap < 0.05 compared to control; bp < 0.05 compared to DEN group).
Histopathological findings
Liver tissues for control and IGLU treated rats showed normal hepatic architecture of normal parenchyma and normal vascular, as well as normal stroma with no inflammatory infiltrate or fibrosis in the portal tracts and no degenerative changes (Figure 5(a) and (c)). Liver specimens with DEN treatment showed significant marked severe fibrosis in-between the degenerated dysplastic hepatocytes as well as inflammatory cell infiltration in portal area (Figure 5(b1)–(b3), Table 2). Administration of IGLU, pre or post DEN exposure, showed a significant moderate fibrosis in the portal area of liver tissues, as well as in-between the degenerated dysplastic hepatocytes without any inflammatory cell infiltration in portal area (Figure 5(d), Table 2), while therapeutic treatment showed significant moderate fibrosis in the portal area and in-between the degenerated dysplastic hepatocytes with inflammatory cell infiltration in the portal area (Figure 5(e1)–(e3), Table 2). These observations showed the protective effect of IGLU supplementation.

DEN: diethylnitrosamine; IGLU: irradiated β-glucan.
Severity: mild (+), moderate (++), and severe (+++).
Groups sharing different superscripts are significantly different (p < 0.05; n = 6).
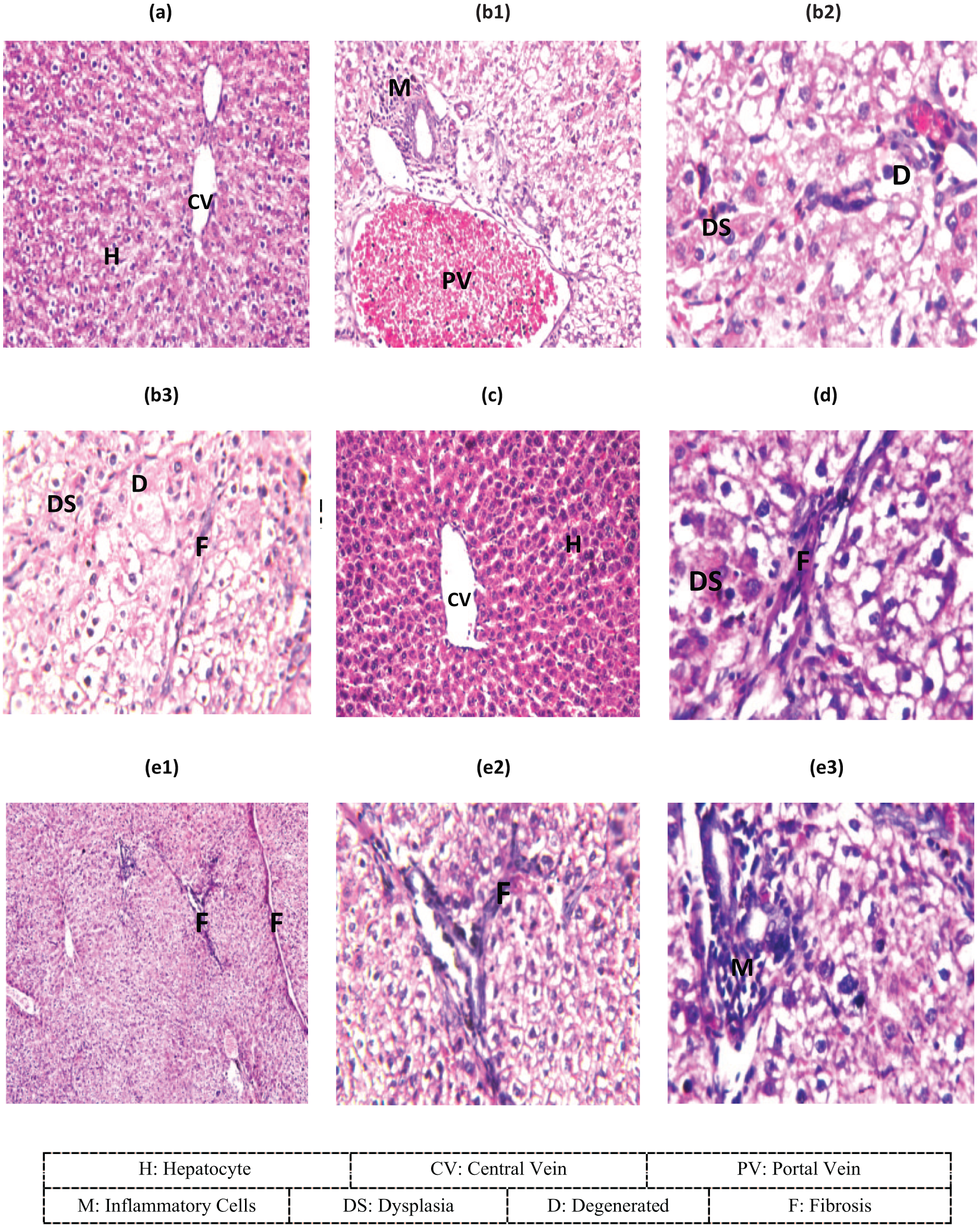
Photomicrographs of liver specimens stained with hematoxylin and eosin. (a) Normal hepatic architecture in control animals. (b1) The portal area with inflammatory cell infiltration and congestion in portal vein in DEN-treated rats. (b2) The degeneration and dysplasia of the hepatocytes all over the parenchyma in DEN-treated rats. (b3) Fibrosis in-between the degenerated dysplastic hepatocytes in DEN-treated rats. (c) Normal hepatic architecture in IGLU treatment. (d) Fibrosis in portal area as well as in-between the degenerated dysplastic hepatocytes in IGLU preventive treatment. (e1 and e2) Fibrosis in portal area and in-between the degenerated dysplastic hepatocytes in IGLU therapeutic treatment. (e3) Inflammatory cells infiltration in the portal area in IGLU therapeutic treatment.
Discussion
HCC is a highly pervasive, treatment-resistant malignancy with a multifaceted molecular pathogenesis. 11 Current evidence reported that during hepatocarcinogenesis, the major pathogenic mechanisms have been linked with alterations in several important cellular signaling pathways. These pathways are of interest from a therapeutic perspective because targeting them may help to overthrow, delay, or prevent tumorigenesis. 13 Improved knowledge of oncogenic processes and the signaling pathways that regulate tumor cell proliferation, differentiation, angiogenesis, invasion, and metastasis has led to the identification of several possible therapeutic targets that have driven the development of molecular targeted therapies. 12
Numerous anti-cancer drugs cause severe adverse effects, including damage to the immune system, which constrains their use in treatment. 16 Therefore, it is important to examine novel anti-tumor drugs that offer improved immune stimulatory and toxicity profiles. In such manner, many polysaccharides and polysaccharide–protein complexes isolated from mushrooms, fungi, yeasts, algae, lichens, and plants are attracting attention owing to their immunomodulatory and anti-cancer effects.8 β-Glucans, which is found in some foods, are accepted to be one of the most powerful immune response modifiers, especially the low molecular weight β-glucans for the high solubility and low viscosity.3,25 In this trend, a strong rationale for targeting growth factors that drive the angiogenic process as a potential therapeutic strategy for the treatment of HCC was explained.
In this investigation, our data showed that stimulation of liver with DEN leads to tissue damage as was established by the elevated levels of ALT, GGT, and AFP. IGLU had no effect on leakage of ALT and GGT. DEN stimulation causes a release of cytokines by liver. The pro-inflammatory cytokines IFN-γ and IL-6 are abundantly produced by hepatocytes in response to DEN administration which exhibited the extent of liver toxicity, while preventive and therapeutic administrations of IGLU revealed a significant hepatic improvement that encounters hepatic toxicity.
AFP, the traditional gold standard and most commonly used biomarker for HCC, has been perceived in the presence of acute and chronic viral hepatitis as well as in patients with cirrhosis caused by hepatitis C. 26 Likewise, IL-6 is a promising tumor marker for HCC. IL-6 may help to identify a subset of HCC patients with low AFP level and may serve as a complementary tumor marker. 27 They concluded that the diagnostic value of IL-6 increased when it is associated with AFP measurement. Combining the two markers provides a new perspective in the diagnosis of HCC. In the same way, IFN-γ is a cytokine known for its immunomodulatory and anti-proliferative action. In the liver, IFN-γ directs the fate of hepatocytes toward either apoptosis or a cell cycle arrest. IFN-γ production is attributed to activated lymphocytes, such as NK cells, T lymphocytes, and natural killer T (NKT cells), which either reside in the liver or are recruited to the liver in response to inflammation and injury. During liver injury and inflammation, hepatocytes increase expression of the transmembrane IFN-γ receptor, which presumably increases their sensitivity to IFN-γ stimulation. 28
The ERK/MAPK pathway is a ubiquitous signal transduction pathway that regulates crucial cellular processes, including proliferation, differentiation, angiogenesis, and survival. 29 Vitally, the overexpression or activation of components of this pathway is believed to contribute to tumorigenesis, tumor progression, and disease metastasis in a variety of solid tumors, 30 which triggers a cascade of specific phosphorylation events. 31 ERK1/2 regulates cellular activity by acting on more than 100 substrates in the cytoplasm and nucleus, including indirect inducers of gene expression, transcription factors, and cell cycle–related kinases. 32
In this investigation, our data showed that DEN treatment induced markedly significant higher levels of the signaling growth factors—ERK-1 and PI3K proteins—while the preventive and therapeutic administrations of IGLU showed a significant improvement. These data suggest that the ERK-1 and PI3K signaling pathways are essential for the DEN-induced hepatocarcinogenesis. Constitutive activation of the PI3K signaling pathway has been firmly established as a major determinant of tumor cell growth and survival in a multitude of solid tumors. 33 In the PI3K signaling pathway, binding of growth factors to their receptors activates PI3K. 34 PI3K subsequently produces the lipid second messenger phospho inositol triphosphate, which in turn activates the serine/threonine kinase AKT. Activated AKT phosphorylates several cytoplasmic proteins, which increases cellular proliferation, decreases apoptosis, and increases cell survival. In normal tissues, this pathway is negatively regulated by tumor suppressor phosphatase on chromosome 10, which targets the lipid products of PI3K. 16
Nitric oxide (NO) plays an important role in HCC development and its progression.35–37 Hepatocytes produce NO in response to several inflammatory stimuli. Tumor cells themselves are able to produce large amounts of NO due to induced expression of iNOS, which may prevail in rapidly growing tumors. Thus, NO production by hepatic tissues is accelerated in patients with HCC. Increased NO generation is well recognized as an essential step in initiating neoplastic transformation along with HCC development and its progression (metastasis). 38 In this investigation, our data showed that the transcript levels of VEGF-A and iNOS genes were significantly higher with DEN treatment. In the treatments using only IGLU, this transcript did not show any significant difference in the levels from controls. However, after treatment with preventive and therapeutic IGLU, the transcript level of these genes was significantly lower than that in the DEN-treated animals. This result demonstrates that the β-glucan treatment had a protective effect.
NO production was significantly increased in liver in the presence of DEN. Since iNOS expression paralleled this increase in NO production, it is likely that this is due to iNOS induction. This increased expression of iNOS protein is preceded by increased mRNA expression of iNOS in the liver stimulated with DEN. iNOS mRNA expression was elevated indicating that induction of iNOS mRNA was continuously triggered by the inflammatory response of the DEN stimulus.
Investigations of human tumor xenografts in immunodeficient mice demonstrated that neutralization of VEGF inhibited tumor growth and decreased blood vessel density in a variety of tumor types. 39 Increased expression of VEGF and VEGF receptors (VEGFRs; which include VEGFR-1, -2, and -3) has been observed in HCC cell lines and tissues, as well as in the serum of patients with HCC.40–42 Furthermore, increased VEGF expression has been reported in cirrhotic and dysplastic liver tissues, suggesting a possible role for VEGF-mediated angiogenesis in hepatocarcinogenesis. 43 VEGF clearly has an important regulatory role in HCC; high levels of VEGF expression have been linked with HCC tumor grade, 44 poor outcome after resection, 45 disease recurrence, poor disease-free and overall survival, 46 vascular invasion, 47 and portal vein emboli. 48
VEGF is a key angiogenic factor, and several agents that target VEGF or VEGFR are currently in development for the treatment of HCC.49,50 Also, ligands that bind to the EGFR, such as EGF, have a vital role in both tumor angiogenesis and proliferation, thought to be primarily through activation of the RAF/MAPK kinase (MEK)/ERK and PI3K/AKT/mechanistic target of rapamycin (mTOR) pathways. It was hypothesized that agents such as tyrosine kinase inhibitors targeting epidermal growth factor (EGF)/EGR receptor (EGFR) signaling may be beneficial in HCC treatment. Tumor growth beyond the size of 1–2 mm 3 requires the formation of new blood vessels in order to supply the malignant tissues with nutrients and oxygen. 51 Central hypoxia is the main driving force of tumor angiogenesis and upregulates pro-angiogenic growth factors like VEGF. Therefore it is not surprising that VEGF is upregulated in most human tumors and that direct correlation with intratumoral microvessel density exists. VEGF signals through VEGFR (tyrosine kinase receptor) on the cell surface. VEGF ligand binding induces dimerization and autophosphorylation of VEGFRs. Phosphorylated tyrosine residues in the receptor serve as a docking site for various signal transduction proteins that can eventually activate cellular processes involved in angiogenesis.37 Anti-VEGF strategies might have a potential in the treatment of HCC, liver fibrosis, portal hypertension, and in the inhibition of malignant ascites.31 VEGF expression levels may be useful as prognostic markers in HCC patients and as predictors of tumor response to treatment. 12
In this investigation, the histopathological study showed that liver tissues with DEN treatment showed significant severe fibrosis in-between the degenerated dysplastic hepatocytes as well as inflammatory cells infiltration in portal area. With IGLU preventive and therapeutic treatments, liver tissues showed significant improvement with β-glucan supplementation upon hepatocytes. These results are in line with those of Aydogan et al. (2013) 51 who observed areas of intracellular vacuolization, sinusoidal dilatation, congestion, and focal necrosis of the liver parenchyma within the ischemia/reperfusion liver injury rat group while these changes were not observed with β-glucan supplementation group suggesting that β-glucan reduced the extent of histological changes. Pretreatment with β-glucan ameliorated liver injury that was attributed to its antioxidant efficacy. Suchecka et al. (2016) 52 showed that high molecular weight β-glucan supplementation inhibits lipid oxidation in lipopolysaccharide (LPS)-treated animals. The results obtained from experiment encourage for dietary intervention with oat β-glucans for liver protection.
Conclusion
Our findings showed that IGLU ameliorated hepatic damage induced by DEN and modulated targeted signal growth factors which contributed to experimental hepatocarcinogenesis in rats. IGLU is considered as a promising treatment toward hepatocarcinogenesis.
Footnotes
Acknowledgements
The authors would like to acknowledge the technical assistance in histopathological examination of Prof. Dr Adel M. Bakeer, Faculty of Veterinary Medicine, Cairo University, Egypt.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.