
Abstract
Bladder cancer has a considerable morbidity and mortality impact with particularly poor prognosis. Curcumin has been recently noticed as a polyphenolic compound separated from turmeric to regulate tumor progression. However, the precise molecular mechanism by which curcumin inhibits the invasion and metastasis of bladder cancer cells is not fully elucidated. In this study, we investigate the effect of curcumin on the bladder cancer as well as possible mechanisms of curcumin. The expression of β-catenin was detected by quantitative real-time polymerase chain reaction and immunohistochemical analysis in a series of bladder cancer tissues. In addition, bladder cancer cell lines T24 and 5637 cells were treated with different concentrations of curcumin. The cytotoxic effect of curcumin on cell proliferation of T24 and 5637 cells was measured by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. The migration and invasion capacity of T24 and 5637 cells were measured by transwell assay. The effects of curcumin on expression levels of β-catenin and epithelial–mesenchymal transition marker were determined by western blotting. The β-catenin expression was significantly upregulated in bladder cancer tissues when compared with corresponding peri-tumor tissues. Furthermore, curcumin inhibited the cell proliferation of T24 and 5637 cells, and curcumin reduced the migration and invasive ability of T24 and 5637 cells via regulating β-catenin expression and reversing epithelial–mesenchymal transition. Curcumin may be a new drug for bladder cancer.
Bladder cancer (BC) is the fourth most common cause of cancer and the eighth most common cause of cancer death in men. 1 While the majority of first diagnosed BC present as non-invasive early-stage tumors, up to one third of non-muscle-invasive bladder cancer (NMIBC) will progress to muscle-invasive bladder cancer (MIBC) and metastasize over time. 2 Because most locally advanced BC patients experience relapse after radical cystectomy, 3 adjuvant chemotherapy is usually performed in an effort to delay recurrence and prolong survival. However, only 50% of MIBC patients have responded to adjuvant chemotherapy. 4 It is an urgent need to identify the new molecular therapeutic drugs for BC.
Curcumin is a phytochemical, naturally occurring from the Curcuma longa, which has anti-inflammatory, anti-oxidative, anti-proliferative, apoptosis-inducing, anti-tumor, and anti-anxiety properties in a variety of cancer types in vitro and in vivo.5–6 At present, some studies have reported that curcumin inhibits epithelial–mesenchymal transition (EMT) in tumors. 7 EMT was identified as a special regulator of metastasis which epithelially polarized cells to become motile mesenchymal-appearing cells.8,9 The Wnt/β-catenin signaling pathway has been implicated in transforming growth factor (TGF)-β1-induced EMT in cancer cells. 10 β-Catenin could mediate the membrane and facilitate the transfer of molecules from the cytoplasm into the nucleus through the Wnt pathway. 11 Curcumin also has anti-tumor effects through the inhibition of the Wnt signaling pathway via downregulating β-catenin expression.12,13 However, the precise molecular mechanism by which curcumin inhibits the invasion and metastasis of BC cells is not fully elucidated. In this study, we investigated the anti-cancer effects of curcumin on BC in vitro. We investigated whether curcumin induced BC cell apoptosis and inhibited BC cell survival and invasion. Furthermore, the mechanisms underlying the role of curcumin on BC were investigated.
Materials and methods
Patients and tissue samples
BC tissues were obtained from a series of 144 consented patients who had received surgical resection at Department of Urology, Chinese PLA General Hospital. Furthermore, 144 normal urothelial specimens, which were more than 3 cm away from the BC tissues, were obtained during surgery and used as normal controls. All these samples were formalin-fixed and paraffin-embedded. Besides, fresh resected tumors and adjacent non-cancerous mucosa were harvested and then immediately frozen in liquid nitrogen and stored at −80°C. This study was approved by the legislation and ethical boards of Chinese PLA General Hospital. All subjects or their caregivers have written informed consent.
Cell lines and culture
The human BC cell lines (T24, 5637, and J82) and HEK 293T cell line were purchased from American Type Culture Collection (Manassas, VA, USA) and maintained in RPMI-1640 medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO, USA) and cultured at 37°C with a humidified environment of 5% CO2 in air. Curcumin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was dissolved in dimethyl sulfoxide (DMSO) and stored at −20°C.
Immunohistochemical analysis
All specimens were fixed in 10% buffered formalin, embedded in paraffin, and sectioned (thickness: 4 µm). Sections were then deparaffinized and rehydrated with xylene and graded alcohol. Subsequently, the sections were washed in phosphate-buffered saline (PBS, pH 7.2) for 10 min. The endogenous peroxidase activity was blocked by incubating in 3% H2O2 at room temperature for 10 min and then heated to 95°C for 30 min for antigen retrieval. After washing with PBS three times, the sections were blocked in goat serum and incubated with primary anti-β-catenin (1:100; Abcam, Cambridge, MA, USA) at 4°C for overnight. Subsequently, the slides were incubated with polymer enhancer (reagent A) and goat anti-mouse antibody (reagent B) and developed in freshly prepared 3,3′-diaminobenzidine (DAB) substrate. Finally, the sections were counterstained with hematoxylin, dehydrated, air-dried, and mounted.
Evaluation of immunostaining
All slides were evaluated by two experienced pathologists who were blinded to the clinical data or the disease outcome. The immunostaining was determined in 10 fields (magnification: 100×) for each slide. Intracellular brown particles were deemed as positive for β-catenin, and positive β-catenin located at the cell membrane, cytoplasm, and nucleus. The results were determined according to the method of Maruyama et al. 14 Normal expression was defined as positive membrane staining which is seen in >70% cells; otherwise, it was deemed as a deletion of membrane expression. Positive cytoplasmic and nuclear expression were defined when staining was observed in >10% cells. Deletion of membrane expression and positive cytoplasmic and nuclear expression were proposed as defined abnormal expression.
Total RNA extraction and quantitative real-time polymerase chain reaction
Total RNA was extracted using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). Complementary DNA was synthesized using the PrimeScript RT Reagent Kit (TaKaRa Biotechnology Co., Ltd., Dalian, China), according to the manufacturer’s instructions. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using SYBR Green PCR Master Mix (Applied Biosystems) in a total volume of 10 µL using the Light-Cycler 480 instrument (Roche Diagnostics, Penzberg, Germany) with the following conditions: 95°C for 5 min; followed by 45 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 15 s; and a final extension step of 72°C for 5 min. The primer sequences used are as follows—β-catenin: 5′-GATTTGATGGAGTTGGACATGG-3′ (forward) and 5′-TGTTCTTGAGTGAAGGACTGAG-3′ (reverse); glyceraldehyde 3-phosphate dehydrogenase (GAPDH): 5′-TGCACCACCAACTGCTTAGC-3′ (forward) and 5′-GGCATGGACTGTGGTCATGAG-3′ (reverse). The qRT-PCR results were calculated according to 2−ΔΔCt, and all experiments were repeated in triplicate.
Western blot analysis
Total protein was extracted using 500 µL of radioimmunoprecipitation assay (RIPA) buffer containing 1% protease inhibitor cocktail (Calbiochem; Roche Diagnostics, Basel, Switzerland). Tissue lysates were centrifuged at 15,000g for 15 min at 4°C, and protein concentrations were measured using the bicinchoninic acid (BCA) protein assay kit (Pierce, Rockford, IL, USA). A volume of 30 µg of protein samples were loaded and separated on 10% sodium dodecyl sulfate (SDS)-polyacrylamide gels and transferred to polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). The membranes were first blocked with 5% non-fat skim milk for 2 h and then incubated with anti-β-catenin (1:1000; Abcam), anti E-cadherin, anti-N-cadherin, anti-vimentin (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and anti-β-actin antibody (Cell Signaling Technology, Inc., Danvers, MA, USA). Specific proteins were detected using an enhanced chemiluminescence kit (Pierce) according to the manufacturer’s instructions.
Cell viability assay
Cell viability was measured by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay which was performed as reported in Xu et al.’s study. 15
Cell colony formation assay
Cells in the experimental or control group (200 cells/well) were plated in triplicate onto six-well plates and cultured for 2 weeks. They were then stained with 0.1% crystal violet, and the colony formation rate was calculated using the following equation: colony formation rate (%) = (number of colonies/number of seeded cells) × 100.
Determination of cell apoptosis by flow cytometry
After treatment, apoptosis was detected using the Annexin V-FITC Apoptosis Detection Kit. Cells were detached by trypsinization and washed three times in PBS, centrifuged at 1000g for 5 min, and resuspended in 195 µL of Annexin V–fluorescein isothiocyanate (FITC) binding buffer. A volume of 5 µL of Annexin V–FITC was added and mixed. Then, the cells were stained in the dark for 10 min at room temperature. After that the cells were centrifuged at 1000g for 5 min and resuspended in 190 µL of Annexin V–FITC binding buffer. Finally, 10 µL propidium iodide (PI) staining solution was added and mixed. The cells were then kept on ice in the dark and immediately subjected to flow cytometry analysis. The data were analyzed using the CellQuest software (BD Biosciences). The experiment was repeated three times.
Wound-healing migration assay
The cells were seeded in culture medium onto 24-well plates at a density of 1.2 × 105 cells per well. The confluent monolayer of cells was scratched with a fine pipette tip, and cell migration into the wound was visualized and scored by measuring the size of the initial wound and comparing it to the size of the wound after 24 h by microscopy.
Transwell cell migration and invasion assay
Cell migration or invasion assays were performed using a 24-well transwell chamber (Corning Incorporated, Corning, NY, USA) with or without Matrigel coating. After 48 h, treated and control cells (1 × 104) were detached and seeded into the upper chamber of an 8-µm pore size insert in the 24-well plate and cultured for another 12 h. The cells were allowed to migrate or invade the bottom chamber containing 15% FBS. The non-migratory cells on the surface of the upper membrane were removed with a cotton tip, and the migratory or invasive cells attached to the lower membrane surface were fixed with 4% paraformaldehyde and stained with crystal violet. The number of migratory and invasive cells was counted in five randomly selected high-power fields under a microscope. The presented data represent three individual wells.
Statistical analysis
The data are presented as mean ± standard error of the mean (SEM). Statistical significance was determined using the Student’s t test. All data were analyzed using SPSS 16.0 (SPSS Inc., Chicago, IL, USA). All statistical tests were two tailed and statistical significance was assumed for p < 0.05.
Results
β-Catenin is overexpressed in BC tissues
qRT-PCR was performed to detect the expression of β-catenin messenger RNA (mRNA) in 144 paired BC tissues and corresponding non-cancerous tissues. β-Catenin expression was significantly high in BC tissues compared with the related normal peri-carcinomatous tissues (Figure 1(a)), indicating that β-catenin expression is commonly elevated in human BC. We then examined the β-catenin expression in clinical BC specimens. Immunohistochemical staining results showed that β-catenin expression in BC specimens was significantly upregulated compared with that in the adjacent non-tumoral tissues. β-Catenin overexpression was observed in 102 of 144 (70.83%) BC specimens when compared with adjacent non-neoplastic tissues (56 of 144, 38.89%), the difference of β-catenin expression was statistically significant (Figure 1(b); p < 0.001).

β-Catenin is upregulated in bladder cancer tissues compared with the corresponding adjacent non-neoplastic tissues. (a) The relative expression levels of β-catenin were determined by qRT-PCR in 144 pairs of bladder cancer tissues and the adjacent non-neoplastic tissues. (b) High expression of β-catenin in the bladder cancer.
β-Catenin is overexpressed in BC cell lines
qRT-PCR and western blot analysis were used to determine the levels of β-catenin in BC cell lines. Results show that the levels of β-catenin mRNA and protein were significantly higher in human BC cell lines T24, 5637, and J82 cells in comparison with HEK 293T cell lines (Figure 2(a) and (b)). Our results showed that T24 and 5637 cells showed the higher expression of β-catenin, so we used T24 and 5637 cells as a model to investigate the effect of curcumin on BC progression.

(a) Quantitative real-time PCR showing the relative expression levels of β-catenin mRNA in human bladder cancer cells in comparison with HEK 293T cell line. (b) Western blots showing the expression levels of β-catenin protein in human bladder cancer cells in comparison with HEK 293T cell line.
Curcumin inhibits the proliferation of T24 and 5637 cells
To determine the inhibitory effect of curcumin on BC cells, MTT assay was performed. When the curcumin was used at varying concentrations of 5, 10, 20, and 30 µmol/L, there is a strong inhibition effect in survival rates of T24 and 5637 cells in a time- and dose-dependent manner, and the difference was statistically significant (p < 0.01; Figure 3(a) and (b)). To further confirm the reduction in cellular viability, colony-forming ability of the T24 and 5637 cells was assessed. As shown in Figure 4(c) and (d), the clonogenic ability of T24 and 5637 cells was reduced after administration of 10 µmol/L curcumin.
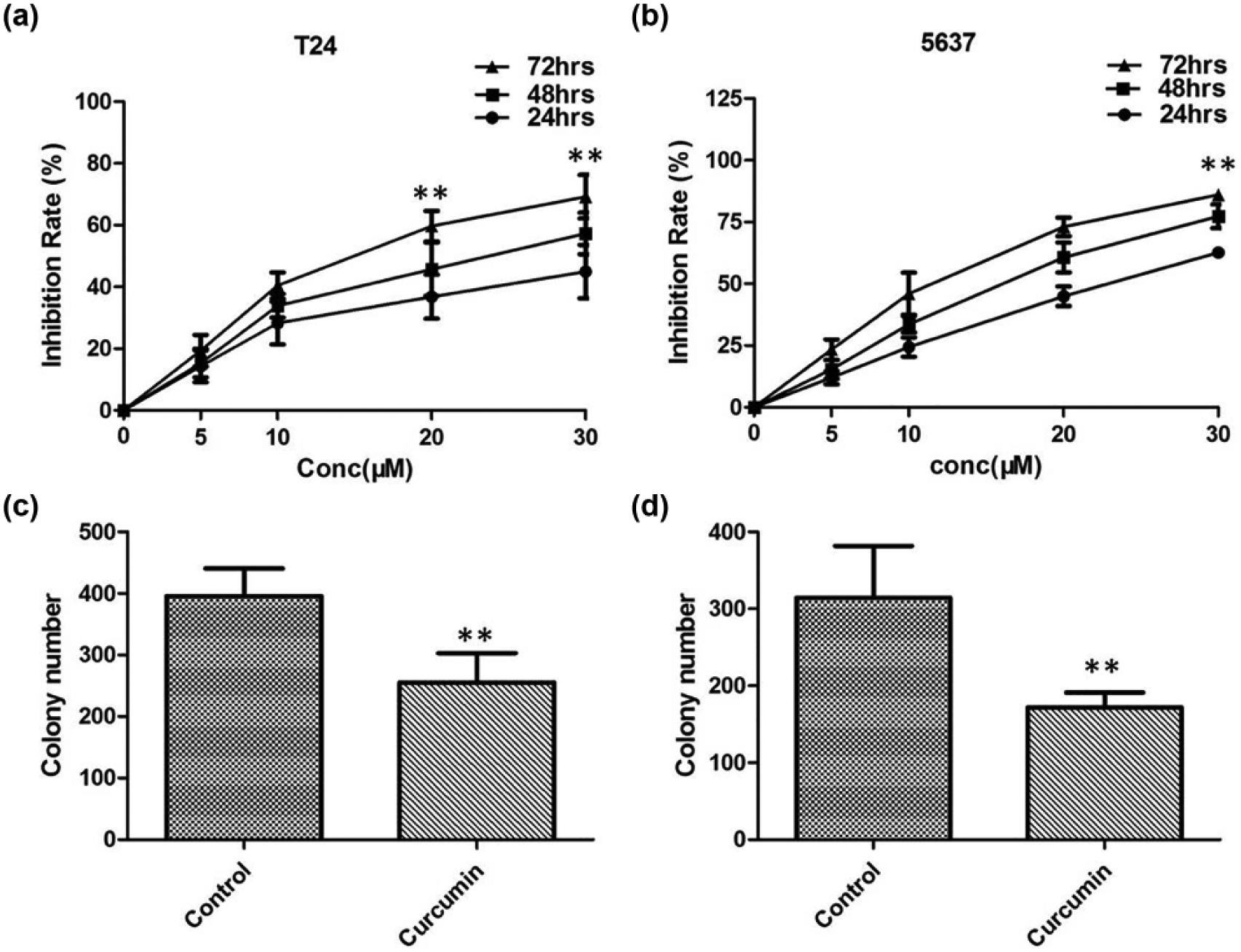
The inhibition effect on T24 and 5637 cells measured by MTT and colony-forming assay. (a) The survival rate of T24 cells with different concentrations of curcumin. (b) The survival rate of 5637 cells with different concentrations of curcumin. (c) The clonogenic ability of T24 cell lines was reduced after administration of curcumin. (d) The clonogenic ability of 5637 cell lines was reduced after administration of curcumin.

(a and b) Apoptosis in T24 and 5637 cells treated with different drugs after 24 h. After administration of curcumin, cells were stained with FITC-conjugated Annexin V and PI, followed by flow cytometric analysis.
Effect of curcumin on the apoptosis of T24 and 5637 cells
To determine whether curcumin induces apoptosis of T24 and 5637 cells, Annexin V–FITC/PI staining was performed. Annexin V and PI staining results showed that with the increase of curcumin concentration after 24 h, the number of apoptotic cells also increased obviously (p < 0.01; Figure 4(a) and (b)), indicating that curcumin could induce apoptosis in both T24 and 5637 cells.
Curcumin suppresses the migration and invasion of T24 and 5637 cells via regulation of EMT and β-catenin
Next, we evaluated whether curcumin plays a role in the inhibition of migration and invasion of BC cells. We carried out a scratch wound-healing and Matrigel invasion assay in T24 and 5637 cells. We found that curcumin (5 and 10 µM) markedly diminished wound-healing capacity of T24 and 5637 cells (Figure 5(a) and (b)). Curcumin (5 and 10 µM) was used for a standard invasion assay by being added to the upper compartment of the invasion chamber. T24 cells penetrating into the lower chamber were significantly decreased after the treatment of curcumin in a dose-dependent manner (Figure 5(c) and (d)).
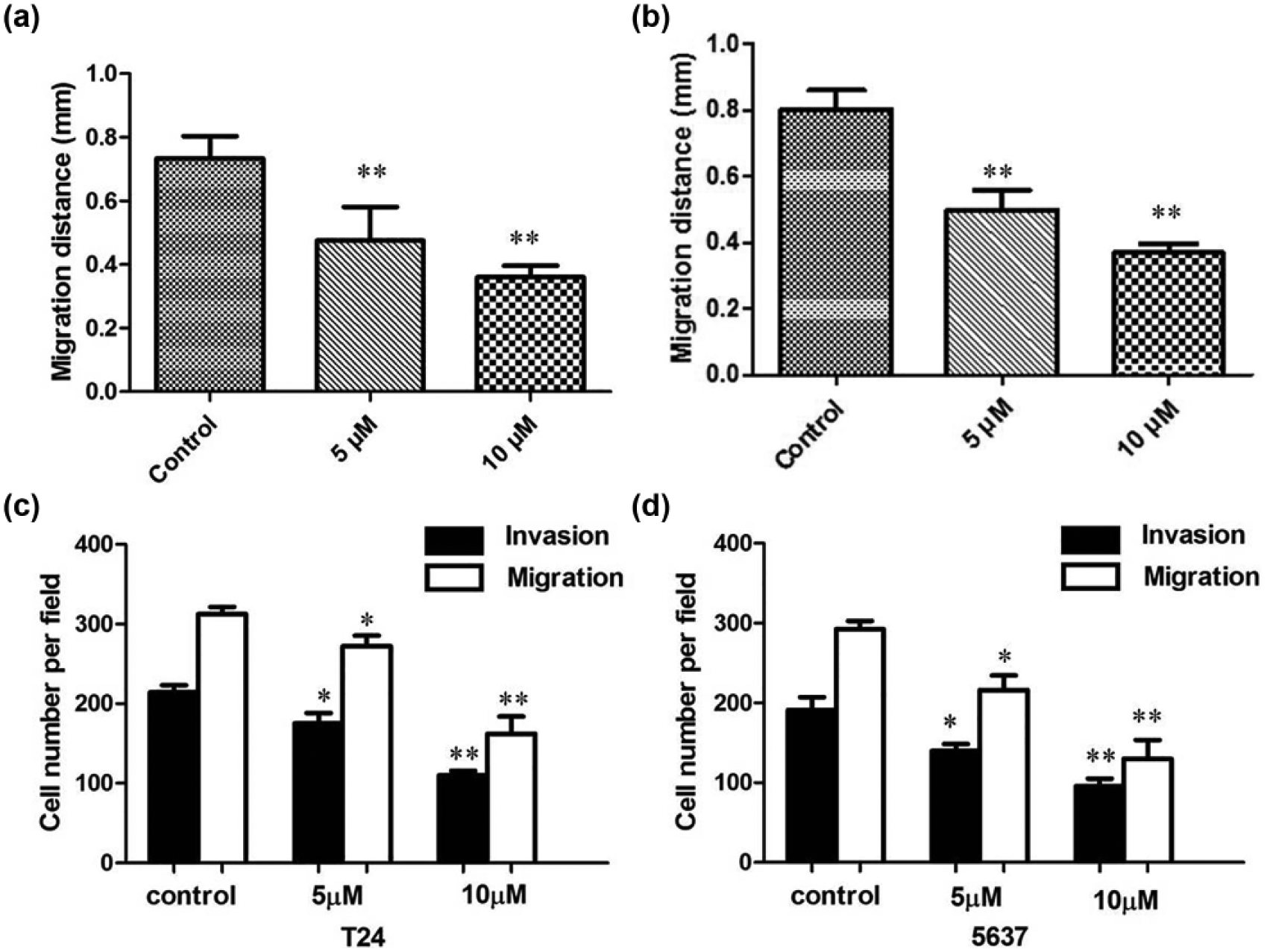
(a) Curcumin markedly diminished wound-healing capacity of T24 cells. (b) Curcumin markedly diminished wound-healing capacity of 5637 cells. (c) Migration and invasion of T24 cells treated with different concentrations of curcumin after 24 h. (d) Curcumin markedly diminished wound-healing capacity of 5637 cells.
In addition, we assessed the expression levels of β-catenin and EMT markers (vimentin, N-cadherin, and E-cadherin) in T24 and 5637 cells under curcumin treatment. Our results showed that the levels of β-catenin, vimentin, and N-cadherin decreased in a dose-dependent manner when the cells were treated with curcumin, and E-cadherin expression was significantly increased (Figure 6(a) and (b)), indicating that curcumin inhibited the invasiveness of BC cells through EMT and β-catenin pathway.

The levels of β-catenin and EMT markers were analyzed by Western blot analysis. (a) Western blot analysis of the expression of proteins after T24 cells treated with different concentrations of curcumin. (b) Western blot analysis of the expression of proteins after 5637 cells treated with different concentrations of curcumin.
Discussion
BC is the second most common urological malignancy around the world and is by far the most frequent urological malignancy in China. 16 At initial diagnosis, nearly 75%–80% of cases are NMIBC, with the remaining MIBC. Despite years of intensive efforts on surgical techniques and adjuvant chemotherapy, BC remains a highly prevalent and lethal malignancy.17,18 Therefore, novel treatment strategies based on new molecular networks are urgently needed to improve the poor prognosis of BC.
The Wnt/β-catenin signaling pathway has been shown to be an essential signal transduction pathway in tumorigenesis and progression of various types of cancer.19–21 Moreover, it has been reported that β-catenin has been considered as a potential therapeutic target for cancer therapy.11,22 Downregulation of β-catenin expression by small interfering RNA (siRNA) inhibited the progression of various cancers. In this study, we accurately detected the expression level of β-catenin in BC tissues, and the results showed that both the protein and mRNA levels of β-catenin were upregulated in the clinical tissues of BC.
In recent years, curcumin has been paid more attention for its anti-cancer activity.23,24 Curcumin blocks cancer cell migration, invasion, angiogenesis, cell cycle, and neoplasia. However, the therapeutic effect of curcumin and its target molecules in BC remains to be clarified. In this study, we found that curcumin could inhibit cell proliferation of T24 and 5637 cells in a time- and dose-dependent manner. Curcumin has shown the ability to effectively modulate apoptotic effect of cancer cells. 25 Our results showed that the number of apoptotic cells increased obviously with the increase of curcumin concentration, indicating that curcumin could induce apoptosis in T24 and 5637 cells. The phytochemical curcumin is a known repressor of several tumor properties, including tumor cell migration. A previous in vitro study has shown that curcumin could impede cell metastasis via downregulation of Src and FAK kinase activity. Our results in agreement with previous reports states that curcumin could inhibit the migration and invasion ability of T24 and 5637 cells.
EMT is characterized by the loss of cell polarity and intracellular junctions and acquirement of mesenchymal features, which contributes to tumor development and metastasis. At present, some studies have reported that curcumin inhibits EMT in tumors. 26 Here, we determined the expression of epithelial markers in BC cells treated with different concentrations of curcumin. Our results showed that E-cadherin expression was significantly increased, while expression of N-cadherin and vimentin was greatly reduced in T24 and 5637 cells treated with curcumin, suggesting that curcumin could reverse EMT.
Loss of E-cadherin-mediated cell adhesion is one of the key mechanisms involved in metastatic conversion of epithelial cells and EMT. 27 One of the probable mechanisms involved in E-cadherin dysfunction, especially loss of its expression and consequent promotion of tumor progression, is through β-catenin signaling. 28 As the main binding partner of β-catenin, E-cadherin plays a pivotal role in β-catenin stabilization and function. In addition, our results showed that the levels of β-catenin decreased in a dose-dependent manner when the cells were treated with different doses of curcumin.
To the best of our knowledge, this is the first study to demonstrate that the anti-metastatic effects of curcumin are associated with the β-catenin. Curcumin inhibited cell proliferation and migration of BC cells through suppression of β-catenin signaling pathways. Results of this study therefore offer a new perspective on the role of curcumin in preventing the progression of BC.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.