
Abstract
Oropharyngeal head and neck squamous cell carcinoma is a common malignant tumor in the oral cavity. High-risk human papillomavirus 16 infection is a major cause of oropharyngeal head and neck squamous cell carcinoma development. Strong antitumor immune responses, especially CD8+ T cell responses, are thought to be essential to effective cancer treatment and are associated with better prognosis in oropharyngeal head and neck squamous cell carcinoma. In this study, we examined the role of the Tim-3/Gal-9 pathway in oropharyngeal head and neck squamous cell carcinoma patients. We found that Gal-9 expression by CD4+ T cells was increased in human papillomavirus-positive oropharyngeal head and neck squamous cell carcinoma patients, but not in human papillomavirus-negative oropharyngeal head and neck squamous cell carcinoma patients. Increased Gal-9 secretion by CD4+ T cells presented multiple immunosuppressive effects. Coculturing monocytes with high Gal-9-expressing CD4+ T cells resulted in the expansion of Tim-3+ monocytes, which suppressed interferon gamma production by activated CD8+ T cells. Subsequently, total monocytes incubated with exogenous Gal-9, or high Gal-9-expressing CD4+ T cells, suppressed the expression of interferon gamma by CD8+ T cells. Exogenous Gal-9 and high Gal-9-expressing CD4+ T cells also suppressed the secretion of both interleukin 10 and interleukin 12 by monocytes. These effects are Tim-3/Gal-9-dependent because blocking Tim-3 and/or Gal-9 could enhance the support of CD8+ T cell interferon gamma production and the interleukin 10 and interleukin 12 secretion by monocytes. Together, these data suggest that the high Tim-3 expression in monocytes could be utilized by tumor-promoting Gal-9 expression on CD4+ T cells. Immunotherapy in human papillomavirus-positive oropharyngeal head and neck squamous cell carcinoma patients therefore faces an additional challenge posed by Tim-3 and Gal-9 and likely requires the blockade of these molecules.
Introduction
Oropharyngeal head and neck squamous cell carcinoma (oral HNSCC) is the most common head and neck cancer, characterized by malignant growth of tumors in oral cavity, oropharynx, hypopharynx, and larynx. In recent years, high-risk human papillomavirus (HPV) 16 infection is identified as a major etiologic cause to the development of oral HNSCC.1,2 Interestingly, it was shown that HPV-positive HNSCC presented better response to conventional therapy than non-HPV tumors, 3 possibly due to antitumor immune responses that target the HPV antigens. Supporting this hypothesis, HPV-positive tumors were sensitive to radiotherapy and cisplatin in immunocompetent mice but not in immunocompromized mice. 4 These results indicated an important role of immune responses in HPV-positive oral HNSCC patients.
Despite recent improvements in conventional therapy, the prognosis of HPV-positive oral HNSCC remains poor. 5 The main challenge is that HPV infection is largely asymptomatic or causes only minor lesions. Discovery of HPV-positive oral HNSCC usually occurs at advanced stages, when extensive local invasions and metastasis could prevent complete surgical removal, leading to high recurrence rate and low 5-year survival rate. 6 Moreover, conventional chemotherapy and radiation therapy are highly toxic. Therefore, there is a need to develop novel therapeutic strategies utilizing the antitumor properties of the immune system.
To achieve this goal, it is essential to understand the cellular and molecular participants in the immune responses that combat HPV-positive oral HNSCC. Many recent studies have demonstrated that the frequency of CD8+ T cells in peripheral blood and in resected tumor and the CD8+/Foxp3+ tumor-infiltrating lymphocyte (TIL) ratio were positively correlated with HPV-positive HNSCC prognosis.7–9 demonstrating an essential role for CD8+ T cells. At the same time, many immune checkpoints exist in solid tumors that prevent and suppress CD8+ T cell antitumor immunity, one of which is the Tim-3/Gal-9 signaling pathway. Tim-3 was first identified as a marker of interferon (IFN)-γ-expressing CD4+ and CD8+ T cells. Upon engaging Gal-9, Tim-3+ T cells demonstrate loss of cytokine production and elevated cell death. 10 Tim-3 is also expressed by myeloid cells, monocytes/macrophages, and tumor-associated dendritic cells. 11 It is known that HPV-positive HNSCC express lower Tim-3/CD45 messenger RNA (mRNA) ratio in the tumor and has better prognosis than HPV-negative HNSCC. 12 The role of the Tim-3/Gal-9 signaling in myeloid cells and how it affects the immune responses in oral HNSCC is still unclear.
In this study, we found that in peripheral blood mononuclear cells (PBMCs) from oral HNSCC patients, Gal-9 was predominantly expressed by CD4+ T cells from HPV-positive oral HNSCC patients and was significantly elevated compared to control individuals. Interestingly, Gal-9-expressing CD4+ T cells promoted the expansion of Tim-3+ monocytes/macrophages through Tim-3/Gal-9-dependent pathway. Exogenous Gal-9 and Gal-9-expressing CD4+ T cell-treated monocytes/macrophages suppressed autologous CD8+ T cell responses, and presented a low cytokine secretion profile, but this immunosuppression could be reverted by blocking Tim-3/Gal-9. Together, these results suggested that immunotherapies in HPV-positive oral HNSCC patients potentially face an additional Tim-3/Gal-9-posed challenge, involving the Gal-9+CD4+ T cells and Tim-3+ monocytes/macrophages.
Methods
Study participants
Age- and sex-matched HPV-positive oral HNSCC patients, HPV-negative oral HNSCC patients, and healthy controls were recruited in The Third Hospital of Hebei Medical University. Clinical-pathological characteristics of the subjects were analyzed by DICAT (Vancouver, Canada), and are summarized in Table 1. No HNSCC patients received neoadjuvant chemotherapy or radiation therapy before blood sample collection. Between 1.5 × 108 and 2 × 108 PBMCs were obtained from heparinized blood by centrifuging across a Ficoll-Paque (GE Healthcare, Little Chalfont, UK) gradient. All patients and controls provided written informed consent. All study procedures were approved by the Institutional Ethics Board at The Third Hospital of Hebei Medical University.
Clinical-pathological information of participants.
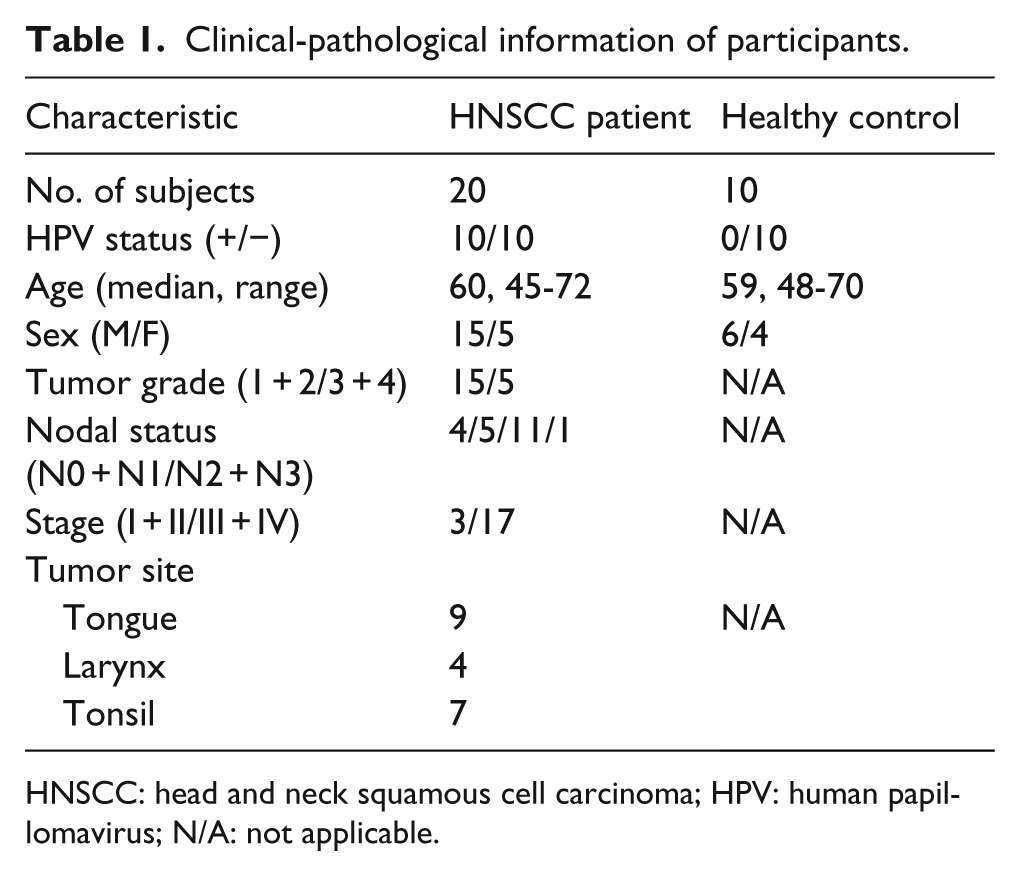
HNSCC: head and neck squamous cell carcinoma; HPV: human papillomavirus; N/A: not applicable.
Cell sorting
Sorting of CD4+ T cells, CD8+ T cells, and monocytes from PBMCs was performed using the EasySep Human CD4+ T Cell, CD8+ T cell, and Monocyte Enrichment Kits (STEMCELL Technologies, Vancouver, Canada), respectively, following the instructions provided by the manufacturer. Sorting of Tim-3− and Tim-3+ monocytes was performed by staining purified monocytes with PE antihuman Tim-3 antibody (BioLegend, San Diego, CA, USA) and then applying the EasySep Human PE Positive Selection Kit (STEMCELL Technologies).
Quantitative real-time polymerase chain reaction
Total mRNA from purified CD4+ T cells was harvested using the RNeasy Kit (Qiagen, Hilden, Germany) following the protocol provided by the manufacturer. Complementary DNA (cDNA) synthesis was performed using the Applied Biosystems High-capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA, USA). Amplification of cDNA was performed using the Power SYBR Green Master Mix (Thermo Fisher Scientific). The following primer sets were used (in 5′-3′ order): Gal-9 forward CAGTGCTCAGAGGTTCCACA, Gal-9 reverse TGAGGCAGTGAGCTTCACAC, β-actin (control) forward GGACTTCGAGCAAGAGATGG, and β-actin reverse AGCACTGTGTTGGCGTACAG. Triplicate experiments were performed for each sample/condition.
T cell/monocyte coculture
CD4+ T cells or CD8+ T cells were cocultured with autologous monocytes at 1/1 ratio. Additional stimulating/blocking reagents were used as follows: anti-CD3 mAb OKT3, anti-CD28 mAb CD28.2 (BioLegend), and lipopolysaccharide (LPS) (Sigma-Aldrich, St. Louis, MO, USA) at 2 µg/mL; recombinant human Gal-9 (R&D Systems, Minneapolis, MN, USA) at 1 µg/mL; neutralizing antihuman Gal-9 mAb 9M1-3 at 5 µg/mL (plate-bound); 13 and anti-Tim-3 blocking mAb F38-2E2 coating at 10 µg/mL. For monocyte-specific coating, 105 per mL purified monocytes were incubated with anti-Tim-3 antibody for 30 min at 4°C.
Flow cytometry
In some experiments, cells were first labeled with CFSE—Cell Labeling Kit (Abcam, Cambridge, UK) before incubation. If intracellular IFN-γ was examined, 1 × brefeldin A and 1 × monensin (eBioscience, Waltham, MA, USA) were added 5 h before harvesting cells. After incubation, cells were washed twice and labeled with LIVE/DEAD Fixable Aqua Dead Cell Stain Kit (Thermo Fisher Scientific) for 30 min at 4°C, such that dead cells could be removed later on in flow cytometry analysis. The cells were then washed once, stained with surface antibodies for 30 min at 4°C, washed twice, and if necessary, stained with intracellular antibodies using CytoPerm/CytoFix and Perm Wash Buffer Kit (BD, Franklin Lakes, NJ, USA) following the manufacturer’s instructions.
Enzyme-linked immunosorbent assays
The Human Galectin-9 and interleukin 10 (IL-10) Quantikine ELISA Kits (R&D Systems) were used for the detection of Gal-9 and IL-10, respectively. The Human IL-12 p70 ELISA Ready-SET-Go Kit (eBioscience) was used for the detection of IL-12.
Statistical analyses
Data normality was assessed using D’Agostino and Pearson test. For comparisons between multiple groups, one-way analysis of variance (ANOVA) followed by Tukey’s test was used, or two-way ANOVA followed by Sidak’s test was used. For paired comparison of the sample at two conditions, paired t-test was used. All analyses were performed in Prism (GraphPad, La Jolla, CA, USA). The p-values less than 0.05, 0.01, or 0.001 were considered significant.
Results
Gal-9 expression is elevated in circulating CD4+ T cells from HPV-positive oral HNSCC patients
Gal-9 is a secreted glycoprotein that binds to the carbohydrate motif on Tim-3. 14 It was shown that CD4+ T helper (Th) cells could suppress and regulate inflammation through the expression of Gal-9.10,15 Therefore, we first examined Gal-9 expression by CD4+ Th cells in HPV-positive and HPV-negative oral HNSCC patients, as well as in healthy controls. Gal-9 mRNA transcription and protein expression by CD4+ T cells were significantly higher in HPV-positive HNSCC patients than in healthy controls and in HPV-negative HNSCC patients (Figure 1(a) and (b)). Gal-9 was originally discovered as a suppressor of Th1 inflammation. Subsequently, we examined Gal-9 expression under anti-CD3/CD28-stimulated conditions, with or without IFN-γ enrichment in cell culture. Again, we found that Gal-9 mRNA transcription and protein expression were significantly higher in HPV-positive HNSCC patients than in healthy controls and in HPV-negative HNSCC patients (Figure 1(a) and (b)). Interestingly, addition of IFN-γ significantly increased Gal-9 transcription and protein secretion by CD4+ T cells from HPV-positive HNSCC patients. Together, these data demonstrated that CD4+ T cells by HPV-positive HNSCC patients presented significantly elevated Gal-9 expression, a feature not shared by HPV-negative HNSCC patients.

CD4+ T cells from HPV-positive oral HNSCC patients presented high Gal-9 expression than healthy controls and HPV-negative oral HNSCC patients. (a) The Gal-9 mRNA transcript level in CD4+ T cells from healthy controls (Control), HPV-positive (HPV+) HNSCC patients, and HPV-negative (HPV−) HNSCC patients. PBMCs were unstimulated, stimulated with anti-CD3/CD28 antibodies, or stimulated with anti-CD3/CD28 antibodies and additional IFN-γ for 24 h. mRNA level was examined in purified CD4+ T cells. (b) The Gal-9 protein secretion by CD4+ T cells from Control, HPV+ HNSCC, and HPV− HNSCC subjects. Purified CD4+ T cells were unstimulated, stimulated with anti-CD3/CD28 antibodies, or stimulated with anti-CD3/CD28 antibodies and additional IFN-γ for another 6 h. Supernatant was harvested for ELISA. *p < 0.05; **p < 0.01; ***p < 0.001.
Gal-9-expressing CD4+ T cells promoted Tim-3+ monocyte enrichment through the Tim-3/Gal-9 pathway
Monocytes make up a significant proportion of circulating PBMCs and express high levels of Tim-3. 16 We investigated the role of CD4+ T cell-mediated Gal-9 secretion on Tim-3+ monocytes. First, we stimulated PBMCs with anti-CD3/CD28, in the absence or presence of additional IFN-γ as described in Figure 1. The CD4+ T cells cultured in the absence of IFN-γ were low Gal-9-expressing (lo-Gal-9) while the CD4+ T cells cultured in the presence of IFN-γ were high Gal-9-expressing (hi-Gal-9) as shown in Figure 1. These CD4+ T cells were then isolated by negative selection, and then cocultured with autologous monocytes from PBMCs for 6 days. We found that CD14+ monocytes incubated with lo-Gal-9 CD4+ T cells and with hi-Gal-9 CD4+ T cells presented similar levels of proliferation (Figure 2(a)), but those incubated with hi-Gal-9 CD4+ T cells were significantly more enriched in Tim-3+ cells at Days 4 and 6 of coculture (Figure 2(b)). This enrichment of Tim-3+ monocytes could be inhibited by neutralization of Gal-9 and/or Tim-3, demonstrating that it required the Tim-3/Gal-9 pathway (Figure 3).
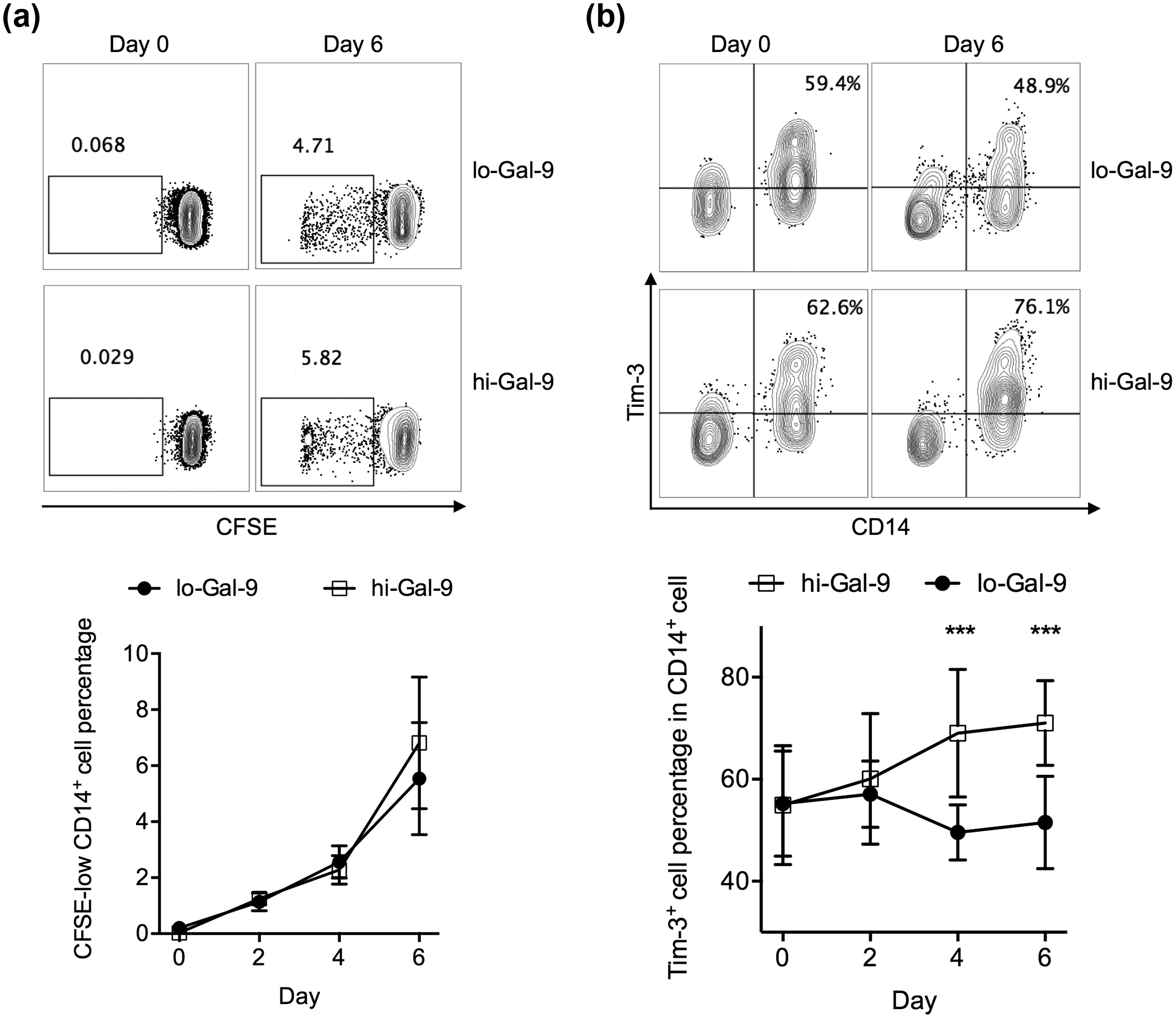
Monocytes cocultured with autologous high Gal-9-expressing CD4+ T cells were enriched in Tim-3+ cells. Low Gal-9-expressing (lo-Gal-9) and high Gal-9-expressing (hi-Gal-9) CD4+ T cells were obtained by stimulating PBMCs for 24 h with anti-CD3/CD28 antibodies and with anti-CD3/CD28 antibodies and additional IFN-γ, respectively, and subsequently purifying CD4+ T cells. (a) The gating strategy and total frequency of dividing (CFSE-low) cells in CD14+ monocytes, after 6-day coculture with lo-Gal-9 or hi-Gal-9 CD4+ T cells. (b) The gating strategy and total frequency of Tim-3+ cells in CD14+ monocytes, after 6-day coculture with lo-Gal-9 or hi-Gal-9 CD4+ T cells. ***p < 0.001.

Blocking Tim-3 and/or Gal-9 significantly reduced the frequency of Tim-3+ cells in monocytes. Monocytes were incubated with hi-Gal-9 CD4+ T cells for 6 days, in blank media, with anti-Gal-9 blocking (plate-bound anti-Gal-9 neutralizing antibody), with anti-Tim-3 blocking (monocytes coated with anti-Tim-3 blocking antibody), or with both anti-Gal-9 and anti-Tim-3 blocking. The percentage of Tim-3+ cells in CD14+ monocytes was then examined by flow cytometry. The difference between each blocking condition with blank media was examined. *p < 0.05; **p < 0.01; ***p < 0.001.
Gal-9-treated and Gal-9-expressing CD4+ T cell-treated monocytes suppressed CD8+ T cell activation
Tim-3+ monocytes/macrophages are thought to be functionally quiescent.16,17 We therefore investigated the outcome of Gal-9-mediated Tim-3+ enrichment in monocytes. First, we investigated the support of CD8+ T cell response by Tim-3− and Tim-3+ monocytes with or without Gal-9 treatment. The anti-CD3/CD28-stimulated CD8+ T cells incubated with Tim-3+ sorted monocytes expressed significantly lower levels of IFN-γ, a CD8+ T cell activation marker, than those incubated with Tim-3− sorted monocytes (Figure 4(a)), demonstrating that Tim-3+ monocytes suppressed IFN-γ secretion in CD8+ T cells. Monocytes pretreated with Gal-9 also suppressed IFN-γ secretion in CD8+ T cells (Figure 4(b)). Similarly, monocytes cocultured with hi-Gal-9 CD4+ T cells resulted in less IFN-γ secretion in CD8+ T cells, than monocytes cocultured with lo-Gal-9 CD4+ T cells (Figure 4(c)). Furthermore, to confirm the involvement of Tim-3/Gal-9 pathway in these assays, we blocked Tim-3 on monocytes and Gal-9 in supernatant during coculture with hi-Gal-9 CD4+ T cells. This procedure significantly elevated the IFN-γ production by CD8+ T cells cocultured with monocytes (Figure 4(d)). Together, these results showed that Gal-9- and Gal-9-expressing CD4+ T cell-treated monocytes suppressed CD8+ T cell activation, due to Tim-3/Gal-9-mediated signaling.
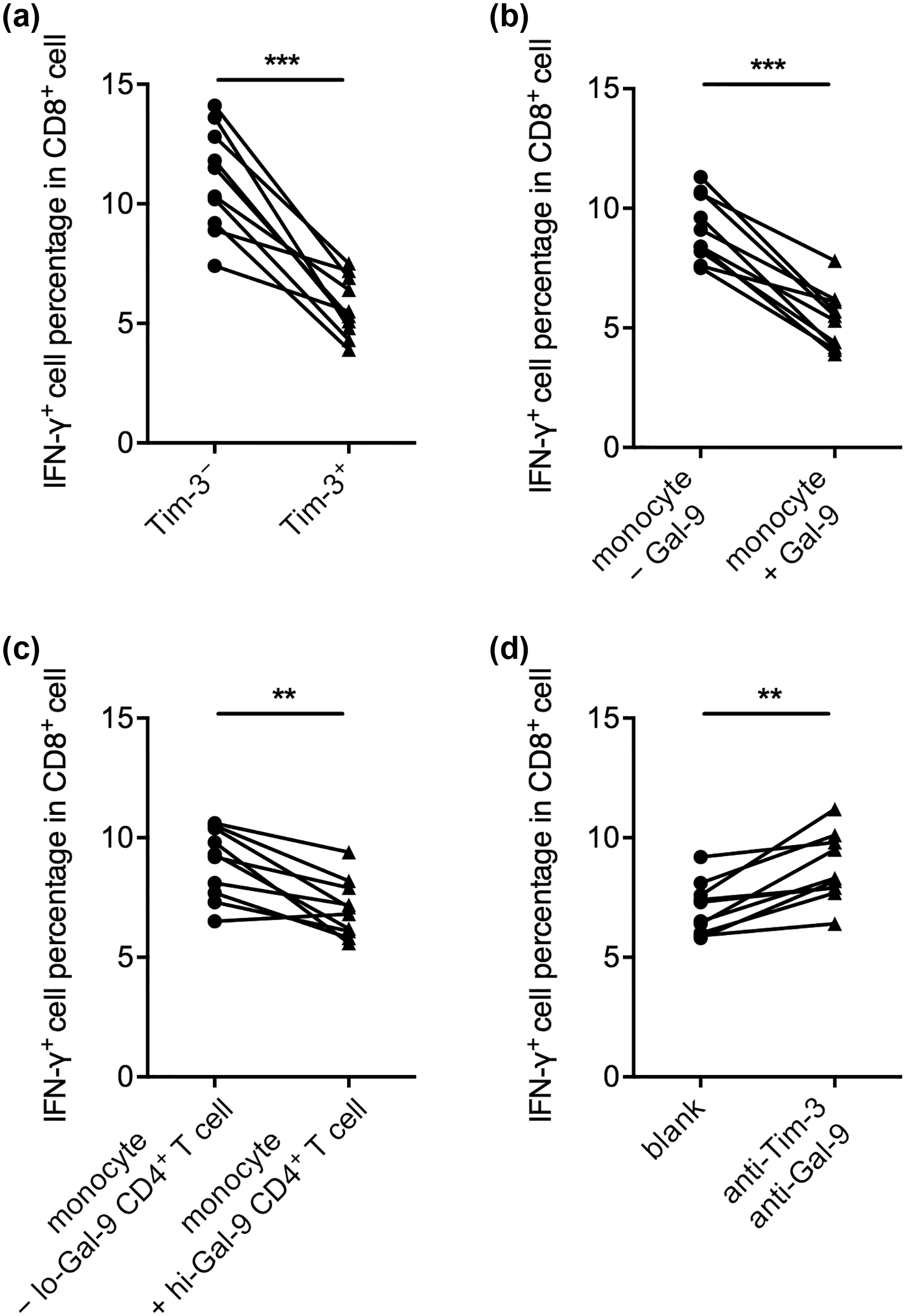
Gal-9- and Gal-9-expressing CD4+ T cell-treated monocytes suppressed CD8+ T cell IFN-γ response due to Tim-3/Gal-9 signaling. Purified CD8+ T cells were cocultured with various types of autologous monocytes for 24 h in the presence of anti-CD3/CD28 antibodies, and then the percentage of IFN-γ+ cells in CD8+ T cells was examined by flow cytometry. The monocytes used were (a) Tim-3−- or Tim-3+-sorted monocytes, (b) whole monocytes preincubated for 6 days without Gal-9 (− Gal-9) or with Gal-9 (+ Gal-9) protein, (c) whole monocytes preincubated for 6 days with lo-Gal-9 CD4+ T cells or with hi-Gal-9 CD4+ T cells, and (d) whole monocytes preincubated for 6 days with hi-Gal-9 CD4+ T cells, in the absence (blank) or presence of Tim-3 and Gal-9 blocking. **p < 0.01; ***p < 0.001.
Gal-9-treated and Gal-9-expressing CD4+ T cell-treated monocytes were impaired at cytokine secretion
Next, we investigated the cytokine secretion by Gal-9- and Gal-9-expressing CD4+ T cell-treated monocytes. IL-10 and IL-12 are crucial to macrophage function and have been used to distinguish macrophage subtypes into classically activated M1 (low IL-10 and high IL-12) and alternatively activated M2 (high IL-10 and low IL-12) types.18,19 Here, we observed that under unstimulated condition, exogenous Gal-9 significantly suppressed the expression of both IL-10 and IL-12 (Figure 5(a)). In LPS-stimulated monocytes, both exogenous Gal-9 and hi-Gal-9 CD4+ T cells suppressed the expression of IL-10 as well as IL-12 (Figure 5(b)). By inhibiting the Tim-3/Gal-9 pathway, the hi-Gal-9 CD4+ T cell-mediated suppression of IL-10 and IL-12 secretion from monocytes could be rescued (Figure 5(c) and (d)). Together, these results demonstrated that Gal-9 and Gal-9-expressing CD4+ T cells could inhibit the IL-10 and IL-12 expression from monocytes in a Tim-3-dependent manner.

Gal-9- and Gal-9-expressing CD4+ T cell-treated monocytes were impaired at IL-10 and IL-12 secretion. Monocytes were incubated alone, with Gal-9 protein (+ Gal-9), or with hi-Gal-9 CD4+ T cells for 6 days (a) without stimulation or (b) with LPS stimulation. The IL-10 and IL-12 secretion were measured in supernatant by ELISA. (c) and (d) Monocytes were incubated with hi-Gal-9 CD4+ T cells for 6 days without blocking (blank) or with anti-Tim-3 and anti-Gal-9 blocking. (c) IL-10 and (d) IL-12 secretion was measured in supernatant by ELISA. *p < 0.05; **p < 0.01; ***p < 0.001.
Discussion
In summary, we demonstrated that Gal-9 expression by CD4+ T cells was increased in HPV-positive oral HNSCC patients, but not in HPV-negative HNSCC patients. Gal-9-expressing CD4+ T cells promoted the expansion of Tim-3+ monocytes, which suppressed IFN-γ production from activated CD8+ T cells. Also, monocytes incubated with high Gal-9-expressing CD4+ T cells were less effective at expressing IL-10 and IL-12, which is in line with another study demonstrating that IL-12 in hepatitis C virus (HCV) infection was primarily expressed by Tim-3− monocytes/macrophages. 17 These data suggested that the inflammatory response of monocytes in HPV-positive HNSCC patients could be obstructed by the Tim-3/Gal-9 pathway. Previously, Tim-3 expression was considered a marker for senescent/exhausted CD8+ T cells.20,21 Since the Tim-3/Gal-9 pathway was detrimental to antitumor responses in both CD8+ T cells and monocytes, broad-spectrum blockade of Tim-3 and Gal-9 might serve as a novel strategy in immunotherapies.
In oral squamous cell carcinoma, HPV-positive patients demonstrated better prognosis than HPV-negative patients, which, as suggested by Licitra et al., 3 might be attributed to genetic instability induced by HPV oncogenes E6 and E7 and increased immune responses against HPV antigens. However, we found that HPV-positive HNSCC monocytes had high Tim-3 expression that could be utilized by Gal-9-expressing CD4+ T cells to suppress IFN-γ response by CD8+ T cells. This Tim-3/Gal-9 pathway may take away some, but not all, advantages “enjoyed” by HPV-positive patients. So overall, HPV-positive patients still fare better than HPV-negative patients, but further improvements can be made in HPV-positive patients’ prognosis by blocking the Tim-3/Gal-9 pathway.
We also demonstrated that the Tim-3/Gal-9 signaling did not favor either M1 or M2 macrophages over the other, but rather suppressed the characteristic cytokines of both types. It is known that IL-12 drives the secretion of IFN-γ in both CD4+ and CD8+ T cells, promotes CD8+ T cell division, and augments antigen-specific CD8+ memory T cell generation.22–24 Therefore, the suppression of IL-12 in monocytes likely contributed to the suppression of CD8+ T cells. The effect of reduction in IL-10 is less clear. In the past, IL-10 was mostly recognized as an anti-inflammatory cytokine that inhibited CD4+ and CD8+ T cell effector functions, but recent data suggested that IL-10 contributed to the activation and expansion of IL-10R-expressing tumor-resident CD8+ T cells, and was shown to enhance tumor rejection in mice.25,26 More studies are needed to elucidate the specific role of IL-10 in oral HNSCC.
The proliferation of CD14+ monocytes was accelerated starting at Day 4 of coculture with CD4+ T cells, possibly resulted from division of newly divided monocytes. The Tim-3 upregulation, however, plateaued at Day 4 and onward. Two nonmutually exclusive reasons could explain this. One, it is possible that there might be two subsets of monocytes, one with the potential to upregulate Tim-3 under Gal-9 stimulation and one without. Two, by Day 4, there were too many monocytes and the amount of Gal-9 per cell was not sufficient for additional Tim-3 upregulation. Shown in Figure 3, Gal-9 expression was required for CD14+ monocytes to express Tim-3. The declining percentage of Tim-3+ monocytes in the lo-Gal-9 group (Figure 2(b)) might also have resulted from the lack of Gal-9 in cell culture by Day 4 and Day 6, such that the concentration of Gal-9 was insufficient to maintain high Tim-3 expression. Further experiments are required to examine this dynamic.
This study also raised more questions regarding the role of the Tim-3/Gal-9 pathway in oral HNSCC. First, it is yet unclear why Gal-9 overexpression was only observed in HPV-positive, but not HPV-negative, oral HNSCC individuals. Since Gal-9 could act to suppress Tim-3+ T cells to prevent excessive inflammation, 10 it is possible that Gal-9 was upregulated in CD4+ T cells response to chronic HPV stimulation. Supporting this, in squamous cervical cancer, Gal-9 expression was significantly associated with the presence of HPV 16 or HPV 18, 27 but direct evidence or specific mechanism of HPV inducing Gal-9 is lacking. Second, cervical cancer patients expressing Gal-9 in tumor tended to present better survival, but the result was borderline insignificant (p = 0.087). 27 Whether Gal-9 by tumor cells and CD4+ T cells has different roles in tumor progression requires further studies. Third, we showed that blocking Tim-3 and/or Gal-9 could inhibit the Tim-3/Gal-9-mediated suppression of monocytes in vitro. Their effect in vivo should therefore be examined in clinical trials.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The author(s) received no financial support for the research, authorship, and/or publication of this article.