
Abstract
Receptor tyrosine kinase–like orphan receptor 2 is an enzyme-linked receptor which specifically modulates WNT5A signaling and plays an important role in tumorigenesis, invasion, and metastasis; however, the precise role of receptor tyrosine kinase–like orphan receptor 2 in cancer is controversial. The purpose of this study was to investigate the expression and role of receptor tyrosine kinase–like orphan receptor 2 in ovarian carcinoma and clarify the biological functions and interactions of receptor tyrosine kinase–like orphan receptor 2 with non-canonical Wnt pathways in ovarian cancer. The result of the human ovary tissue microarray revealed that the receptor tyrosine kinase–like orphan receptor 2–positive rate increased in malignant epithelial ovarian cancers and was extremely higher in the metastatic tumor tissues, which was also higher than that in the malignant ovarian tumor tissues. In addition, high expression of receptor tyrosine kinase–like orphan receptor 2 was closely related with ovarian cancer grading. The expression of receptor tyrosine kinase–like orphan receptor 2 protein was higher in SKOV3 and A2780 cells than OVCAR3 and 3AO cells. Knockdown of receptor tyrosine kinase–like orphan receptor 2 inhibited ovarian cancer cell proliferation, migration, invasion, and induced morphologic as well as digestive state alterations in stably transfected SKOV3 cells. Detailed study further revealed that silencing of receptor tyrosine kinase–like orphan receptor 2 reversed the epithelial–mesenchymal transition and inhibited non-canonical Wnt signaling. Our findings suggest that receptor tyrosine kinase–like orphan receptor 2 may be an important regulator of epithelial–mesenchymal transition, primarily regulated the non-canonical Wnt signaling pathway in ovarian cancer cells, and may display a promising therapeutic target for ovarian cancer.
Keywords
Introduction
Ovarian cancer is the most lethal gynecologic malignancy worldwide. 1 The majority of women are diagnosed after the primary tumor has already metastasized, resulting in a 5-year survival rate that has changed little over the past 20 years. 2 Given the heterogeneity and invasiveness of ovarian cancer, there is a need to further understand ovarian cancer biology and develop effective treatments.
A widely recognized behavior of ovarian cancer is its ability to migrate and seed in the peritoneal cavity with exfoliated tumor cells and the formation of ascites. 3 Studies have revealed that the tumor biology is tightly linked in many ways to embryonic development. One such signaling pathway is Wnt signaling, which regulates cell growth, differentiation, and tumor formation. 4 This multifaceted signaling network is traditionally described through the following two distinct mechanisms: canonical pathway (β-catenin dependent) and non-canonical pathway (β-catenin independent).
The non-canonical Wnt pathway regulates cytoskeleton-mediated processes and establishment of polarity by activating two subpathways (Wnt-Ca2+ and planar cell polarity (PCP)). 5 Recently, experiments had revealed that WNT5A, as a non-canonical Wnt protein, is upregulated and contributes to cancer cell migration and invasion in several cancers6–8; however, emerging evidence indicates that the role of WNT5A can be altered depending on the availability of pivotal ligand–receptor pairings. 9 Hence, targeting a receptor rather than a secreted ligand is more desirable. 10
Receptor tyrosine kinase–like orphan receptor 2 (ROR2), acting as a receptor or co-receptor for WNT5A, has been shown to regulate WNT5A-induced activation of the c-Jun N-terminal kinase (JNK) pathway. 11 ROR2 is normally expressed at high levels during developmental processes, playing a key role in cell movement and polarity. 12 To date, tumor suppressive relationships of ROR2 have been established with colon cancer, 13 hepatocellular carcinoma,14,15 and multiple hematologic malignancies 16 ; however, there are several lines of evidence indicating that ROR2 is upregulated or promotes tumor growth, migration, and invasion in a multitude of tumor types, such as melanomas, osteosarcomas, and renal cell carcinoma cells.17–19 Accordingly, similar to WNT5A, ROR2 appears to have dual roles and can act to suppress or promote carcinogenesis in different tissues. 20
It has been documented that WNT5A is upregulated during ovarian carcinogenesis and mediates the migration and invasion of ovarian cancer cells,21,22 while the expression of ROR2 and its regulatory mechanisms of downstream signaling pathways in ovarian cancer are not elucidated. Given the important role of ROR2 in the regulation of the Wnt pathway and carcinogenesis, we analyzed the expression and role of ROR2 in ovarian carcinoma, which has not been highlighted until now.
Materials and methods
Cells, plasmids, and transfection
Human epithelial ovarian cancer cell lines (SKOV3, A2780, 3AO, and OVCAR3) were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in RPMI-1640 medium (Gibco, Auckland, New Zealand) supplemented with 10% fetal bovine serum (FBS; TBD, Tianjin, China) and incubated under standardized conditions (5% CO2 at 37°C).
Three different ROR2 short hairpin RNAs (shRNAs) and a negative control shRNA were purchased from GenePharma (Shanghai, China). All shRNA sequences were inserted into the vector, PGPU6/GFP/Neo. The three shRNA sequences targeting ROR2 were as follows: 5′-GCCCGATTCCAACTCTGAAAG-3′ (shROR2#1), 5′-GCCCAGCCAAGACATGGAAAT-3′ (shROR2#2), and 5′-GCAACCTTTCCAACTACAACA-3′ (shROR2#3). The negative control shRNA sequence was 5’-GCCCAGCCAAGACATGGAAAT-3’ (negative control, NC). Target plasmids were transfected into cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. For transient transfection, the transfection efficiency was examined under florescence microscopy after 48 or 72 h of transfection, and cells were harvested for further analysis. For stable transfection, selective medium with 500 µg/mL of G418 (Calbiochem) was used for 4 weeks. Stable transfected clones were collected and maintained in medium containing 350 µg/mL of G418 for further studies.
Immunohistochemical staining
Human ovary tissue microarrays (OV1005a and OV808) were purchased from the US Biomax, Inc. (Alenabio Biotechnology, Xi’an, China). The microarrays contained tissues from 3 normal ovaries, 17 cancer adjacent normal ovaries, 18 benign cystadenomas, 7 borderline cystadenomas, 45 primary malignant epithelial ovarian cancers, as well as 50 metastatic ovarian cancers. Immunohistochemistry for ROR2 was performed as described previously. 23 All the samples were observed and photographed with the Olympus IX51 Inverted Microscope (Olympus Optical, Melville, NY, USA). The product of the intensity scores was used as the final ROR2 staining score. Staining intensity was scored as 0, 1, 2, or 3 (no staining, weak, moderate, or strongly positive). The final evaluation criteria of the ROR2 expression was designated as negative or positive (negative: 0–1, positive: 2–3). All the scores of ROR2 expression were determined independently by two senior pathologists.
Western blot analysis
After the protein concentrations were quantified using the bicinchoninic acid (BCA) kit (Thermo Fisher Scientific, Waltham, MA, USA), whole-cell lysates were equally (20 µg) loaded on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) transfer membranes (Solarbio, Beijing, China) which were incubated with methanol. The membranes were blocked in 5% non-fat dried milk in Tris-buffered saline containing 0.05% Tween-20 (TBST) for 2 h, then separately incubated overnight with VEGFA (vascular endothelial growth factor A), MMP9 (matrix metallopeptidase 9), MMP13, E-cadherin, keratin, N-cadherin, vimentin, Snail, Slug, ZEB1 (zinc finger E-box binding homeobox 1), Rac1, RhoA, p-PKCδ (phospholipase protein kinase C delta), NF-κB (nuclear factor-kB) p65, β-actin, GAPDH (glyceraldehyde 3-phosphate dehydrogenase) antibodies (Cell Signaling Technology, Danvers, MA, USA), ROR2 antibody (BioVision, Milpitas, CA, USA), and JNK antibody (R&D, Minneapolis, MN, USA) at 4°C. Horseradish peroxidase–conjugated secondary antibody (Zhongshan Jinqiao Biotechnology Co., Ltd, Beijing, China) was added and incubated for 1 h at room temperature. Detection was performed using an enhanced chemiluminescence system (Thermo Fisher Scientific). Quantification was analyzed using ImageJ software (Version 1.47v; Wayne Rasband, National Institutes Health, USA).
Immunofluorescence
Cells were fixed in 4% paraformaldehyde for 30 min, permeabilized in 0.1% Triton X-100 for 10 min, and blocked with 3% bovine serum albumin (BSA) for 30 min at room temperature. The cells were then incubated with the appropriate antibodies overnight at 4°C, followed by incubation for 1 h at room temperature with secondary fluorescent antibodies. Finally, the cells were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen) for 5 min to label the nuclei. Images were acquired using an Olympus IX51 Inverted Microscope (Olympus Optical)
Cell viability assay
The control and shROR2 ovarian cancer cells were plated on 96-well plates at a density of 2.5 × 103 cells per well with 100 µL of complete culture medium. A 10 µL of Cell Counting Kit-8 (CCK8; WST-8 (2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium), Dojindo Molecular Technologies, Inc., Kumamoto, Japan) solution was added to each well at baseline and incubated for 24, 48, 72, and 96 h. Absorbance was measured using a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA) at wavelength of 450 nm. The experiments were repeated three times.
Colony formation assay
To assay colony formation, infected cells were plated into six-well plates at an initial density of 500 cells/well. The interval of medium renewal was 2 days. After 14 days incubation, cells were fixed with 4% paraformaldehyde for 30 min at room temperature and stained with freshly prepared Giemsa for 30 min. The number and size of colonies in each group were counted and recorded.
Wound healing assay
Cells infected with shROR2 and control SKOV3 were seeded on six-well plates until 90% confluency in a monolayer was reached. Then, an in vitro wound was produced using a sterile p200 pipette tip. The floating cells were aspirated with phosphate-buffered saline (PBS) for washing and replaced with 2 mL of fresh serum-free medium. An image of the wound closure was taken at 100× magnification at the indicated time points (0, 24, and 48 h). The experiments were repeated three times.
Matrigel invasion assay
Transwell chambers (Costar, Cambridge, MA, USA) with 8-µm pore polycarbonate filters were used for cell invasion assays. Each transwell was coated with 50 µL of a 1:4 dilution of Matrigel (BD Biosciences, Bedford, MA, USA). Approximately, 2.5 × 104 cells were resuspended in the upper chamber in 100 µL of serum-free medium. Corresponding medium with 10% FBS (500 µL) was placed in the lower chamber as a chemoattractant. After a 24 h of incubation, the cells on the lower surface of the chamber were fixed, stained with crystal violet, counted, and imaged. Cell invasion was quantitated microscopically by counting the number of cells in five random fields (200×). The experiments were repeated three times.
In vivo tumor xenograft model
BALB/c female nude mice, 6 weeks old, were obtained from the Animal Center of Beijing Medical University. For measurement of tumor growth in vivo, the stable single cell clones of SKOV3-shROR2 and control SKOV3 cells (1 × 107 cells/mouse (200 µL)) were injected subcutaneously into the left flank of nude mice (n = 5). After 5 weeks, tumors were isolated from nude mice, weighed, fixed in formalin, and prepared for standard histologic examination. Mice were manipulated and housed according to the protocols approved by the Animal Care Committee of Shandong University.
Statistical analysis
The values in the figures are expressed as the mean value ± standard deviation (SD). The figures in this study were representative of three different experiments. Statistical analysis of the data between the control and treated groups was performed using a Student’s t test. p values < 0.05 were considered statistically significant. All the statistical analyses were performed using GraphPad Prism (version 5.01; GraphPad Software, San Diego, CA, USA).
Results
ROR2 protein expression level was excessively elevated in malignant and metastatic epithelial ovarian cancer tissues
The ROR2 immunohistochemical staining results of different pathological types of ovarian epithelial tissues were shown in Figure 1. The positive rate of ROR2 expression was 28.57% in normal and benign tumor tissues, 57.14% in borderline ovarian tumors, 60.00% in malignant epithelial ovarian cancer, as well as 96.00% in metastatic disease tissues. The positive rate of ROR2 was extremely higher in malignant epithelial ovarian cancers and metastatic tumor tissues than that in normal and benign tissues (malignant epithelial ovarian cancers: p = 0.0174, metastatic tumor tissues: p < 0.0001), and was apparently highest in the metastatic tumor tissues (malignant epithelial ovarian cancers vs metastatic tumor tissues: p < 0.0001). In addition, the ROR2 positive rate was observably higher in grading 2 and 3 (1 vs 2, p = 0.0058; 1 vs 3, p = 0 .0190). However, we failed to detect the correlation between ROR2 expression and ovarian cancer stage or patient age (Table 1).
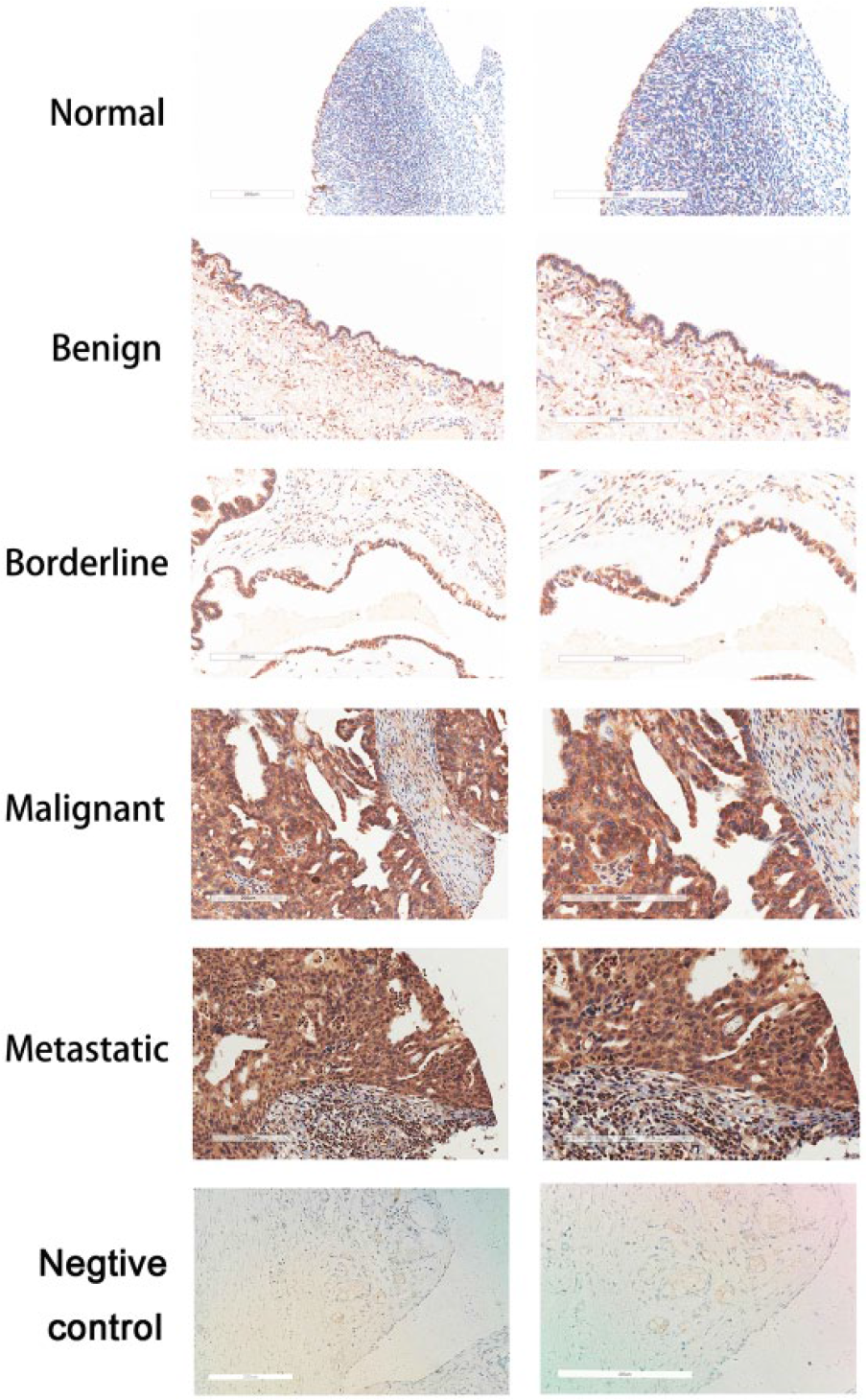
Immunohistochemical analysis of ROR2 expression in normal ovarian epithelial tissues; benign, borderline, malignant, and metastatic ovarian tumor tissues; and the negative control tissues (left panels: original magnification, 200×. Right panels: 400× magnification of area indicated on the left.).
Patients’ clinical and pathological characteristics and their association with ROR2 expression.

ROR2: Receptor tyrosine kinase–like orphan receptor 2.
Values in parentheses indicate percentage values.
Evaluated by χ2 test.
Average age of all cases was 47 years old (range: 16–75).
Knockdown of ROR2 suppressed ovarian cancer cell proliferation, migration, and invasion in transient-transfected SKOV3 and A2780 cells
To determine the level of ROR2 expression in ovarian cancer cells, we measured the expression of ROR2 protein in four different ovarian cancer cell lines. As shown in Figure 2(a), the levels of ROR2 expression were significantly higher in SKOV3 and A2780 cells than those in OVCAR3 and 3AO cells. Thus, SKOV3 and A2780 cells were selected as models for gene knockdown assays. The expression of ROR2 protein decreased significantly and similarly by each of the three shROR2 plasmids (Figure 2(b) and (c)). So, we chose shROR2#1 and shROR2#2 plasmids as models for further investigation. After that, immunofluorescence assay was used to detect the transfection efficiency of the shROR2#1 and shROR2#2. As shown in Figure 2(d) and (e), the intensity of ROR2 decreased significantly after the transfection. The CCK8 assay showed that silencing of ROR2 significantly suppressed the proliferation of SKOV3 and A2780 cells in vitro (p <0.050; Figure 3(a) and (b)). Similarly, silencing of ROR2 significantly inhibited SKOV3 (Figure 3(c)) and A2780 (Figure 3(d)) cell migration in vitro as indicated by the wound healing assay. Moreover, silencing of ROR2 also suppressed ovarian cancer cell invasion in vitro as indicated by the transwell invasion assay (Figure 3(e) and (f)).

Effects of the shROR2 on ROR2 expression in ovarian cancer cells. (a) The expression of ROR2 in cell lysates from SKOV3, A2780, 3AO, and OVCAR3 cells was detected by western blotting. (b) Silencing of ROR2 in SKOV3 cells markedly decreased ROR2 expression. (c) Silencing of ROR2 in A2780 cells markedly decreased ROR2 expression. The expression of ROR2 detected by western blotting. Data are mean ± SD (**p < 0.01). (d) Immunofluorescence staining of ROR2 in SKOV3 cells transfected plasmids of NC, shROR2#1, and shROR2#2. (e) Immunofluorescence staining of ROR2 in A2780 cells transfected plasmids of NC, shROR2#1, and shROR2#2. Nuclei were stained with DAPI.

Knockdown of ROR2 suppressed ovarian cancer cell proliferation, migration, and invasion in vitro. (a) The cell proliferation of NC and shROR2#1, shROR2#2 groups in SKOV3 cells. (b) The cell proliferation of NC and shROR2#1, shROR2#2 groups in A2780 cells. The effect on cell proliferation was determined by CCK8 assay at 0, 24, 48, 72, 96 h. Values are mean ± SD, n = 3. (c) Representative wound healing images of SKOV3 cells at 0, 24, and 48 h. (d) Representative wound healing images of A2780 cells at 0, 24, and 48 h (magnification: 100×; scale bars: 200 µm). (e) Representative invasion images of SKOV3-ROR2 silenced and NC cells. (f) Representative invasion images of A2780-ROR2 silenced and NC cells (magnification: 200×; scale bars: 100 µm). Data are mean ± SD (*p < 0.05; **p < 0.01).
Stable knockdown of ROR2 induced morphologic and digestive state alterations in SKOV3 cells
We successfully established shROR2 stably transfected SKOV3 cell line, named SKOV3-shROR2. Fluorescent imaging of SKOV3 cells transfected with shROR2 showed that >90% of green fluorescent protein (GFP)-expressing cells were observed, indicating successful infection (Figure 4(a)). Moreover, stable downregulation of ROR2 was detected by western blot analysis in SKOV3-shROR2 cells compared with control SKOV3 cells (Figure 4(b)). Interestingly, we observed that SKOV3-shROR2 cells lost the fibroblast-like morphology with spindle shapes and acquired an epithelium-like appearance (Figure 4(c)). Moreover, SKOV3-shROR2 cells showed less resistance to trypsinization than the control cells, presumably indicating less adhesion to the extracellular matrix (ECM). Furthermore, deletion of ROR2 in SKOV3 cells also increased cell–cell contact after digestion, while the control cells attached to the culture dish more tightly (Figure 4(d)).
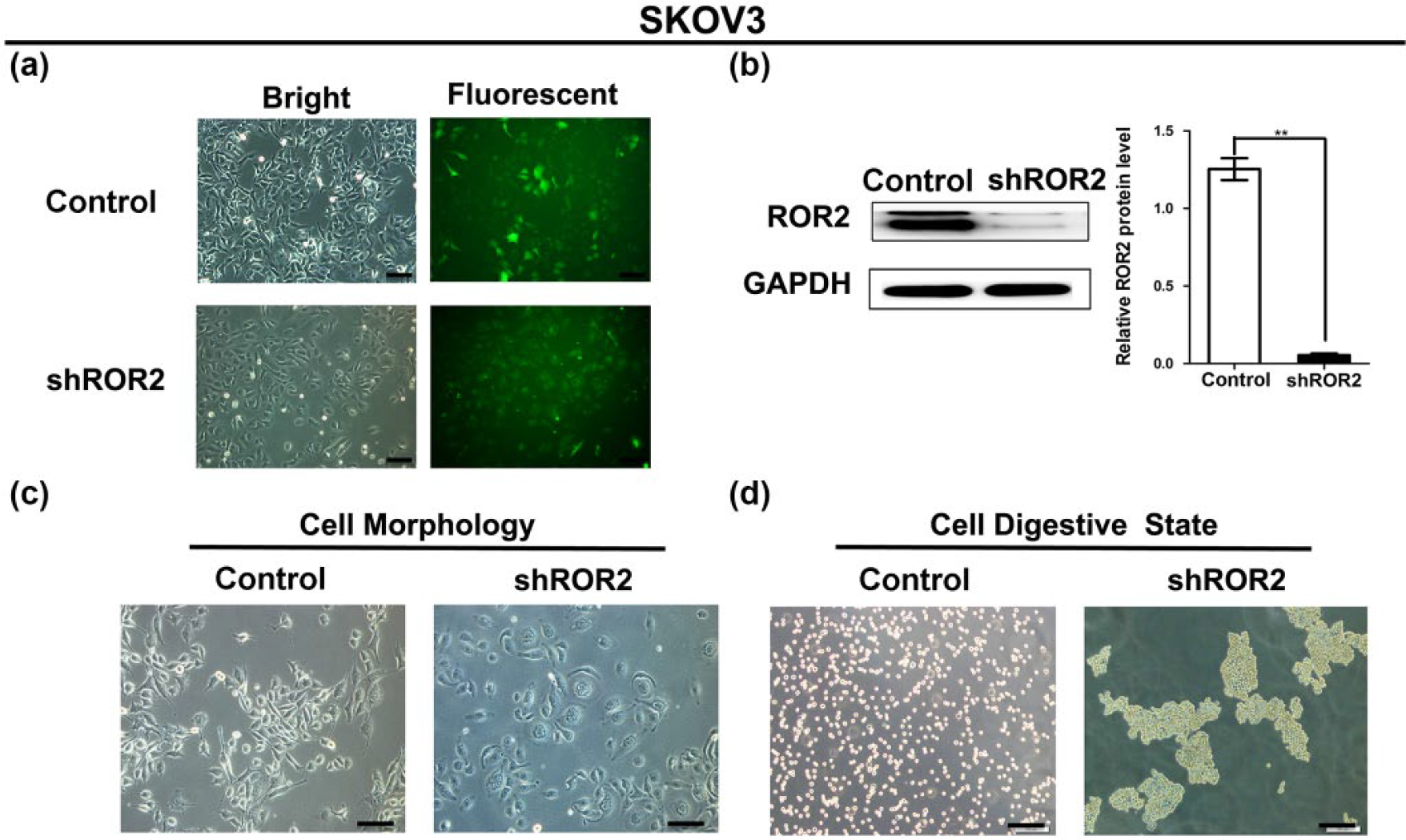
Knockdown of ROR2 induced morphological and digestive state alterations in stably transfected SKOV3 cells. (a) Fluorescent imaging of SKOV3 cells transfected by plasmids of NC and shROR2 (magnification: 200×; scale bars: 100 µm). (b) Stable downregulation of ROR2 was detected by western blotting in the SKOV3-shROR2 cells compared with NC transfected SKOV3 cells. Data are mean ± SD (**p < 0.01). (c) Cell morphology of SKOV3-shROR2 cells and NC cells (magnification: 200×; scale bars: 100 µm). (d) Cell digestive state of SKOV3-shROR2 cells and NC cells (magnification: 100×; scale bars: 200 µm).
Stable knockdown of ROR2 changed the ovarian cancer cell’s biological behavior in vitro and suppresses tumor growth in vivo
To further confirm the functional role of ROR2 in ovarian cancer cells, we examined the effects of ROR2 on cell migration, invasion, and proliferation in SKOV3-shROR2 cells. Compared to control cells, SKOV3-shROR2 cells expressed lower invasion and proliferation abilities, which were similar to the results of transient transfection (Figure 5(a)–(c)). Furthermore, the colony formation assay showed that silencing of ROR2 resulted in inhibition of colony formation of SKOV3 cells (Figure 5(d) and (e)). To confirm the functional effect of ROR2 in ovarian cancer cell proliferation in vivo, SKOV3-shROR2 cells were inoculated subcutaneously into nude mice. After 5 weeks, tumors derived from SKOV3-shROR2 cells were significantly smaller than the control (Figure 5(f) and (g)). ROR2 expression in the tumor xenografts was detected by western blot analysis, and the results showed that the level of ROR2 expression in SKOV3-shROR2 xenografts was much lower (Figure 5(h)). Moreover, significant downregulation of VEGFA and MMP9 was found in SKOV3-shROR2 xenografts. However, no change was identified in the expression of MMP13 in these tumor xenografts (Figure 5(h)).

Stable knockdown of ROR2 changed the ovarian cancer cell’s biological behavior in vitro and suppressed tumor growth in vivo. (a) Wound healing images of NC stably transfected SKOV3 cells and SKOV3-shROR2 cells at 0, 24, and 48 h (magnification: 100×; scale bars: 200 µm). (b) Invasion images of SKOV3-shROR2 and NC stably transfected SKOV3 cells. (magnification: 200×; scale bars: 100 µm). (c) The cell proliferation of SKOV3-shROR2 cells and the control cells was determined by CCK8 assay at 0, 24, 48, 72, 96 h. Values are mean ± SD, n = 3. (d) Images have shown the number and size of colonies in each group. (magnification: 100×; scale bars: 200 µm). (e) Statistical analysis of the number of colonies in each group. Data are mean ± SD (**p < 0.01). (f) Morphologic characteristics of tumors from mice inoculated with SKOV3-shROR2 cells and the control cells. (g) Tumor volumes and tumor weights of NC and shROR2 groups from (f), n = 5. Data are mean ± SD (**p < 0.01). (h) Stable downregulation of ROR2, VEGFA, and MMP9 was found in tumor xenografts, no change was identified in the expression of MMP13 in these tumor xenografts, detected by western blotting and normalized with GAPDH expression. Data are mean ± SD (**p < 0.01).
Stable knockdown of ROR2 reversed the epithelial–mesenchymal transition in SKOV3 cells
To explore the mechanism of morphologic change in SKOV3-shROR2 cells (Figure 3(c)), we examined the potential effect of ROR2 on the epithelial–mesenchymal transition (EMT) in ovarian cancer. Based on western blot analysis and immunofluorescence, we found that ROR2 knockdown in SKOV3-shROR2 cells led to the upregulation of epithelial markers (E-cadherin and keratin) and downregulation of mesenchymal markers (N-cadherin and vimentin; Figure 6(a), (b), and (e)). Besides, EMT key transcription factors, Snail and Slug were significantly decreased in SKOV3-shROR2 cells (Figure 6(c), (d), and (f)). Taken together, these findings indicated that knockdown of ROR2 reversed EMT transition in SKOV3 cells.

Stable knockdown of ROR2 reversed epithelial–mesenchymal transition in SKOV3 cells. (a) ROR2 knockdown upregulated E-cadherin and keratin expression and downregulated N-cadherin and vimentin expression in SKOV3-shROR2 cells. (b) Quantitative analyses of markers in SKOV3 cells showed in (a). Data are mean ± SD (**p < 0.01). (c) ROR2 knockdown downregulated Snail, Slug, and ZEB1 expression in SKOV3-shROR2 cells. (d) Quantitative analyses of E-cadherin and N-cadherin in SKOV3 cells showed in (c ). Data are mean ± SD (**p < 0.01). (e) Immunofluorescence staining of E-cadherin and N-cadherin were observed following ROR2 suppression. (f) Immunofluorescence staining of Snail and Slug were observed following ROR2 suppression. Nuclei were stained with DAPI.
Knockdown of ROR2 resulted in an alteration of downstream targets of non-canonical Wnt pathways
We also evaluated the effect of shROR2 transfection on β-catenin-independent Wnt signal in SKOV3 cell line in vitro. As shown in Figure 7(a) and (b), stable knockdown of ROR2 in SKOV3 cells led to a substantial decrease in β-catenin-independent targets, including Rac1, RhoA, JNK, p-PKCδ and NF-κB p65. In addition, immunofluorescence assay showed marked reduction in fluorescent signals for Rac1 and RhoA after ROR2 suppression (Figure 7).

Knockdown of ROR2 resulted in an alteration of downstream targets of non-canonical Wnt pathways. (a) ROR2 knockdown downregulated Rac1, RhoA, JNK, p-PKCδ, and NF-κB p65 expression in SKOV3-shROR2 cells, detected by western blotting. (b) Quantitative analyses of these downstream targets of non-canonical Wnt pathways in SKOV3 cells. Data are mean ± SD (**p < 0.01). (c) Immunofluorescence staining of Rac1 was observed following ROR2 suppression. (d) Immunofluorescence staining of RhoA was observed following ROR2 suppression. Nuclei were stained with DAPI.
Discussion
It has been reported previously that deregulated receptor tyrosine kinases (RTKs) play critical roles in cancer development and progression. 12 Such variations in kinase activation are now recognized as rational targets for therapeutic intervention in many cancers. 24 The recently described non-canonical Wnt RTK, ROR2, has been shown to play a pivotal role in numerous cancers. Furthermore, ROR2 appears to possess dual roles and can act to suppress or promote carcinogenesis in different tissues. 20 Our present research indicated that the ROR2 positive ratio is very low in normal ovarian surface epithelial cells and benign ovarian tumors. Nevertheless, the ROR2 positive rate was significantly higher in borderline ovarian tumors, malignant epithelial ovarian tumors, and metastatic tumor tissues. It indicates that the ROR2 expression may be correlated with tumor malignancy and, especially, the metastasis of ovarian cancer. These data support the view that ROR2, as an oncogene in ovarian cancer cells, promotes migration, invasion, and proliferation of ovarian cancer cells.
ROR2, a receptor or co-receptor for WNT5A, can promote cell proliferation, migration, and invasion in a multitude of tumor types. WNT5A–ROR2 signaling in mesenchymal stem cells promoted proliferation of gastric cancer cells. 25 Knockdown of ROR2 inhibited cell proliferation and colony formation in osteosarcoma cells by inducing arrest in cell cycle progression. 26 Besides, ROR2 expression was increased in epithelial ovarian cancer, and silencing ROR2 strongly inhibited the proliferation, migration, and invasion abilities of ovarian cancer cells. 27 Furthermore, silencing ROR2 in chemoresistant ovarian cancer significantly inhibited cell migration and invasion. 28 Similarly, this study confirmed that ROR2 expression was increased in SKOV3 and A2780 ovarian cancer cells. Knockdown of ROR2 significantly suppressed cell migration and invasion in vivo, and dramatically decreased the proliferative capability of ovarian cancer cells in vitro and in vivo. Unfortunately, we did not perform this procedure in non-cancer ovarian surface epithelial cells, which would show us more clearly whether ROR2 was involved in the initiation of tumor growth. Since previous report had demonstrated that ovarian surface epithelial cells can develop into papillary carcinoma in mice by introducing carcinogenic gene, such as HER2/neu. 29 It should be a very interesting hypothesis, and more research should be carried out to explore it.
The EMT is a critical process during the early steps of tumor progression, during which tumor cells disseminate from the primary tumor. Recent novel insight into the EMT has focused on the formation of invasive structures, reorganization of the actin cytoskeleton, and tumor metastasis. 30 In this study, knockdown of ROR2 not only inhibited cell migration and invasiveness but also caused more epithelial morphologic characteristics in SKOV3-shROR2 cells. Such changes are associated with increased expression of the epithelial markers, E-cadherin and keratin, and reduced expression of the mesenchymal markers, N-cadherin and vimentin, resulting in weakened motility and enhanced adhesion ability. The transcription factors, Snail, Slug, and ZEB1, which have been identified as critical stimulators of the EMT,31,32 were also shown to be downregulated in ROR2 knockdown cells. From these, the loss of ROR2 in ovarian cancer cells played a reverse role in the EMT transition. In human renal proximal tubule cells, silencing ROR2 could decrease the expression of transforming growth factor beta 1 (TGFβ1), which is a potent inducer of EMT. 33 Therefore, detection of the TGFβ1 expression in shROR2-SKOV3 cells as well as the rescue experiment for mesenchymal-epithelial transition (MET) upon ROR2 knockdown using TGFβ1 might add to the strength of our research, which would be performed in the further study. Accumulating evidence has shown that the EMT is associated with enhanced synthesis of MMPs, which are clearly implicated in ECM remodeling, cellular recruitment of malignant tumors, and tumor cell motility.34,35 Based on in vivo experiments, we found decreased expression of MMP9 accompanied with ROR2 knockdown; however, no change was identified in the expression of MMP13 in these cells. A previous study reported that MMP13 had no expression in two ovarian cancer cell lines (BG-1 and OAW-42); 36 these findings indicated that cell type–dependent expression of members of the MMP family is implicated in ovarian cancer, in which MMP9, but not MMP13, exhibits the characteristic EMT gene profile. Moreover, VEGFA, as an important regulator of angiogenesis, was decreased following ROR2 knockdown, suggesting that angiogenesis might be regulated by a network involving non-canonical Wnt pathways.
Previous reports have shown that non-canonical Wnt pathways are mainly subdivided into the Wnt/PCP and Wnt/Ca2+ pathways.37–39 As molecules of Wnt/PCP downstream signaling pathways, Rac1 and RhoA are well-studied cytoskeletal regulators that function to participate in regulating cell migration, morphogenesis, and the EMT of a broad range of cell types. 40 In this study, ROR2 knockdown in SKOV3 cells resulted in a significant decrease of β-catenin-independent targets (Rac1 and RhoA) at the protein level. It appears that ROR2 may be a part of larger program of gene expression and play a direct role in regulating Rac1 and RhoA pathways, as a critical downstream mechanism by which ROR2 may regulate changes in cell migration and the actin cytoskeleton. JNK, one of the downstream targets of ROR2, which might regulate a distinct branch of non-canonical Wnt signaling,11,38 was also decreased at the protein level following ROR2 silencing. Being calcium-sensitive enzymes, protein kinase C (PKC) can mediate calcium signaling, which plays an important role in regulating the cell migration. 41 PKCδ, a member of PKC families, also plays a role of PCP modulator, can activate Dvl for membrane recruitment and promote breast cancer progression. 37 In agreement with these findings, our results showed that inhibition of ROR2 signaling pathway resulted in inhibition of phospholipase PKCδ (p-PKCδ) expression, and cell impairing invasion ability supervened. The nuclear factor NF-κB is recognized as a crucial transcription factor in many steps of cancer initiation and progression, and the p65 transactivator subunit of NF-κB, RelA, was recognized as a potential oncogene and prognostic factor in metastatic serous ovarian carcinoma. 42 Consistent with this, we found NF-κB p65 expression was decreased following ROR2 silencing. Nevertheless, we are unable to conclude that ROR2 is involved in the non-canonical Wnt pathways. Indeed, we tested the protein level of β-catenin-independent target expression, but the activities of Rac1, RhoA, JNK, and NF-κB were not analyzed in this study, so further investigation is warranted and the related research is underway.
Taken together, our findings highlight the carcinogenic role of ROR2 in ovarian cancer cells. ROR2 silencing inhibited cell proliferation, migration, and invasiveness as well as the EMT, which may primarily be involved in the inactivation of non-canonical Wnt signaling. All our results suggest the ability of ROR2 to serve as a potential drug target for ovarian cancer therapy.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All procedures performed in the studies involving animals were in accordance with the ethical standards of the institution at which the studies were conducted.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the National Natural Science Foundation of China (Grant number: 81101981).